急性熱應激對雞呼吸系統損傷及肺臟熱休克蛋白表達的影響
2021-03-10 08:27:52段滇寧陳超楊潤澤洪麒翔陳洪博唐姝張曉輝
畜牧與獸醫 2021年3期
段滇寧,陳超,楊潤澤,洪麒翔,陳洪博,2*,唐姝,張曉輝
(1. 龍巖學院生命科學學院,福建 龍巖 364012;2. 福建省預防獸醫學與獸醫生物技術重點實驗室,福建 龍巖 364012;3. 南京農業大學動物醫學院,江蘇 南京 210095)
熱應激影響動物精神狀態、生產性能、健康狀況,造成多種器官的損傷,甚至導致動物猝死,熱應激造成的經濟損失已在世界范圍內廣泛報道[1]。與哺乳動物相比,家禽由于代謝旺盛、生長速度快,羽毛豐厚、皮膚沒有汗腺等特點而更容易受到熱應激的損傷[2]。作為一種非特異性應激源,熱應激導致多器官受損。前期研究證實,熱應激對雞心臟組織造成嚴重的傷害[3-4]。肺臟是機體進行氣體交換的場所,熱應激后雞的呼吸次數明顯增多。然而,目前對熱應激后雞肺臟組織損傷研究缺乏。
熱休克蛋白(heat shock proteins,HSPs)是細胞內一類高度保守的蛋白質,保護細胞免受應激原刺激,在許多生理過程中起著至關重要的作用,如蛋白質折疊、運輸和組裝。根據分子量不同,將HSPs分為幾個家族,如小熱休克蛋白家族、HSP47、HSP60、HSP70、HSP90和HSP110[5]。前期研究表明,HSPs能保護心肌細胞免受熱應激的影響,HSPs過量表達后可降低熱應激對心肌細胞的損傷[6-7]。肺臟作為機體重要生命器官,在氣體交換、參與代謝等方面具有重要作用,然而熱應激對呼吸系統的損傷及肺臟組織內HSPs的表達變化規律仍不清楚。本試驗旨在通過急性熱應激處理,觀察熱應激后肉雞肺臟組織形態學變化,在組織和細胞水平探討肺臟組織HSPs的表達與組織損傷的關系,闡明熱應激誘發呼吸系統機能障礙的發生機理。
1 材料與方法
1.1 試驗材料
人工氣候培養箱(隔離器,RXZ-500D)為浙江寧波新江南機械有限公司產品;鼠抗HSP27、HSP60、HSC70、HSP72和HSP90單克隆抗體購自Enzo Biochem Inc.;山羊抗鼠IgG-HRP、BCA 蛋白檢測試劑盒和化學發光底物檢測試劑盒均購自上海碧云天生物公司;乳酸脫氫酶(LDH)試劑盒、血鉀檢測試劑盒和血鈣檢測試劑盒購自南京建成生物技術公司;1日齡SPF雛雞購自南京乾元浩生物技術有限公司,飼養于江蘇省農業科學院實驗動物中心。
1.2 試驗設計
將60只1日齡SPF雛雞飼養于溫度(25±1)℃,相對濕度(45±5)%人工氣候室環境中,自由采食和飲水,常規免疫。飼養至35日齡時,將SPF雞隨機分為對照組和熱應激1、2、3、5和10 h組,每組10只,共6組。將熱應激組雞轉移至(35±2)℃,相對濕度(70±10)%的人工氣候室,試驗開始后觀察雞的臨床癥狀,分別按照試驗設定的時間進行處理。試驗結束后翅靜脈采血取樣,分離血清;頸靜脈快速放血致死,迅速剖殺取材,采集試驗雞肺臟組織,一部分固定于10% 中性福爾馬林溶液(24 h后換新的中性福爾馬林溶液),另一部分經液氮速凍后-80 ℃保存備用。
1.3 呼吸頻率的統計
觀察雞腹部羽毛起伏次數,統計1 min腹部羽毛起伏的次數,即為呼吸頻率。
1.4 血液生化、血氣指標檢測
取對照組和熱應激組血清,用pH計測定血清pH值,血清中LDH、鉀和鈣濃度測定按照試劑盒說明書進行測定。
1.5 病理組織學檢測
常規制作石蠟切片,然后切成5 μm的連續切片,HE染色,鏡檢。具體步驟參考文獻[8]。
1.6 肺臟組織蛋白提取
取雞肺臟組織100 mg,置于2 mL離心管中,加200 μL預冷的RIPA裂解液(含1% PMSF),4 ℃條件下將組織用手持式研磨器快速研磨均勻,-20 ℃冰箱反復凍融3次后12 000 r/min離心5 min,用移液槍吸取上清至新的離心管中,即得到組織總蛋白,BCA法測定樣品蛋白濃度,-80 ℃保存備用。
1.7 Western blot檢測HSPs表達
將樣品與6×蛋白變性緩沖液混合后煮沸變性5 min,冷卻備用取25 μg樣品進行檢測,一抗1∶1 000,稀釋,4 ℃孵育過夜,二抗1∶10 000稀釋,室溫孵育2 h,顯影后進行灰度掃描,Quantity One軟件進行分析。具體實驗步驟參考文獻[9]。
1.8 統計分析
用SPSS 20.0統計軟件提供的多重比較的單因素分析對熱應激組與試驗中的差異進行統計分析。每個試驗重復3次,結果以平均值±標準差表示。*表示差異顯著,P<0.05;**表示差異極顯著,P<0.01。
2 結果
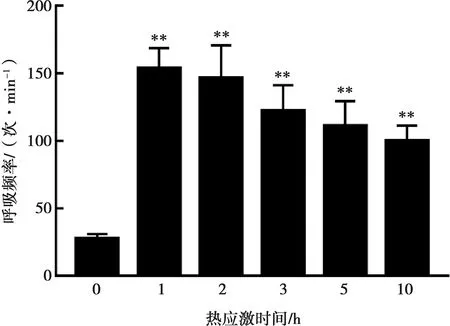
2.1 急性熱應激對雞行為指標影響
急性熱應激對雞呼吸頻率影響如圖1所示,試驗開始前雞呼吸頻率為25次/min,熱應激1~10 h呼吸頻率極顯著升高(P<0.01),臨床表現為熱應激后雞采食次數減少,飲水次數增加,站立雞數量減少,俯伏休息雞數量增多,雞翅膀垂下、張口呼吸。熱應激3 h后試驗組雞呼吸頻率開始降低,但仍極顯著高于正常對照組。
2.2 急性熱應激對雞血液生化、血氣指標的影響
與對照組相比,熱應激3 h后血清pH顯著升高(P<0.05),熱應激1 h后血清pH呈升高趨勢,但與對照組無顯著差異,整個熱應激過程中血清pH值都呈升高狀態(圖2A)。血清LDH水平均在熱應激后呈極顯著升高的變化(P<0.01),LDH變化總趨勢呈升高趨勢,在熱應激5 h組達到最高點,隨后LDH濃度開始降低(圖2B)。與對照組相比,熱應激1 h后血鉀濃度有升高趨勢,但差異不顯著(P>0.05);隨著熱應激時間增加,血鉀濃度開始降低,熱應激5、10 h后血鉀濃度顯著減低(P<0.05);熱應激10 h 組血鉀濃度高于熱應激5 h組,但二者之間差異不顯著(P>0.05),見圖2C。熱應激后血鈣濃度呈降低趨勢,熱應激5、10 h后血鈣濃度顯著減低(P<0.05),熱應激10 h組血鈣濃度高于熱應激5 h組,但二者之間差異不顯著(P>0.05),見圖2D。
注:各組不同時間點與對照組相比,*:P<0.05差異顯著,**:P<0.01差異極顯著。下同
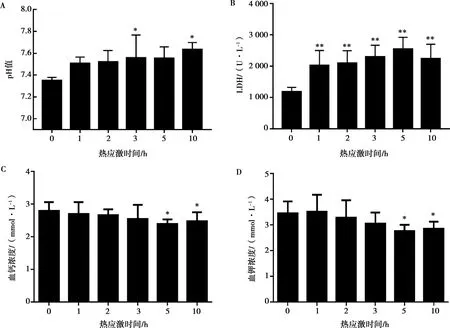
圖2 熱應激對雞血清pH(A)、LDH(B)、鈣濃度(C)及鉀濃度(D)的影響
2.3 急性熱應激雞肺臟組織學變化
對照組雞肺臟組織可見肺房內無異物出現(圖3A)。熱應激前1~3 h后,肺組織內血管尤其是肺泡壁毛細血管顯著充血,肺房結構基本完整(圖3B、圖3D)。熱應激5 h后肺房內充血性變化明顯,肺房內有大量的紅細胞存在(圖3E)。熱應激10 h肺房內的異物減少,但肺上皮細胞大量脫落,組織結構損傷嚴重(圖3F),氣管黏膜高度水腫,黏膜的完整性受到嚴重的損傷,氣管黏膜上皮細胞大面積脫落。

A,B,C,D,E和F分別為熱應激0,1,2,3,5和10 h時的肺臟組織
2.4 肺臟組織中HSPs的表達
由圖4可知,HSP27和HSP72在正常肺臟組織中含量較低,熱應激后表達量極顯著升高(P<0.01)。HSP27在熱應激1 h后表達量極顯著升高(P<0.01),在熱應激過程中一直維持較高水平。HSP72在熱應激1 h后表達量顯著升高(P<0.05),熱應激5 h后表達量極顯著升高(P<0.01),隨著熱應激時間延長,HSP72含量開始降低,但仍顯著高于對照組(P<0.05)。與對照組相比,HSP60的表達量在熱應激1 h后顯著升高(P<0.05),而隨著熱應激時間增加HSP60表達量降低,與對照組無顯著差異。HSC70作為結構型HSPs在熱應激雞肺臟組織表達變化無明顯差異,始終維持在較高水平。熱應激后雞肺臟組織內HSP90表達量呈下降趨勢,熱應激50 h后表達量顯著升高(P<0.05),隨著熱應激時間延長,HSP90含量開始降低,但與對照組相比無顯著差異(P>0.05)。
3 討論
熱應激是一種非特異性的應激原,它會影響家畜的健康甚至導致死亡。課題組前期研究揭示了熱應激可對雞心臟、肝臟和腎臟造成嚴重損傷[8,10-11],對肺臟損傷研究報道較少。熱應激發生時,動物會出現喘息、頻繁呼吸,導致機體CO2排出過多、血液pH值升高等現象。為了保持機體電解質和酸堿平衡,腎上腺皮質分泌醛固酮增加,血液中Na+、K+排出量升高,引起血中鈉、鉀離子水平下降,導致血漿滲透壓降低[12]。本試驗中隨著熱應激時間增加,雞飲水增加,俯臥休息增多,翅膀伸展頻次增加,呼吸頻率的明顯升高,肺臟通氣過度,熱應激1~2 h維持在較高頻率,隨著熱應激時間延長,呼吸頻率降低,但仍極顯著高于對照組。呼吸加快導致大量CO2排出,使血液pH值升高,在熱應激試驗過程中一直維持較高的狀態,表明機體發生呼吸性堿中毒,機體CO2濃度降低可引起腦血管收縮和血流量減少,導致腦組織缺氧,使雞由開始煩躁不安狀態轉化為精神沉郁、萎靡不振,甚至昏迷。與對照組相比,熱應激后3 h后pH與對照組差異顯著,但熱應激后5 h pH與對照組無顯著差異,可能原因是熱應激3 h以后雞群慢慢適應所處的環境,呼吸頻率開始降低,通過大量飲水、體內某些蛋白大量表達來發揮保護作用(如熱休克蛋白等),緩解機體呼吸性堿中毒。本研究中發現血鉀濃度在熱應激后降低,原因可能是為了維持機體的酸堿平衡,腎小管上皮細胞代償性排出H+減少(保酸),相應的排出K+增多,細胞外液K+進入細胞內交換H+,因此使血鉀濃度降低。另外,由于血液pH值升高,機體分泌有機酸增多,有機酸與鈣離子大量結合,導致血鈣濃度下降,血鈣濃度降低,引起神經肌肉細胞閾電位下降,肌肉組織興奮性升高,使雞出現肌肉抽搐,甚至發生痙攣。LDH被廣泛用作細胞損傷的標志物,本研究中熱應激后雞血清LDH水平明顯高于對照組,表明熱應激對雞組織細胞可造成一定的損傷。另外,課題組前期研究發現在熱應激2 ~ 3 h時雞有死亡現象,但隨后死亡率反而降低,熱應激10 h以后雞群又開始出現死亡[4,13],而本試驗中則沒有出現雞因急性應激所致的應激性猝死現象,這可能與飼養環境中空氣流通、濕度和溫度控制較好有關[1]。

同一指標在熱應激不同時間進行比較,*表示差異顯著(P<0.05),**表示差異極顯著(P<0.01)
病理組織學檢測發現熱應激雞肺臟呈現明顯的動態病理變化,主要表現為支氣管擴張充血,肺小葉毛細血管擴張,肺房內有水腫液和紅細胞,氣管黏膜的完整性受到嚴重的破壞,肺房上皮細胞脫落、破裂會嚴重影響肺臟的呼吸功能,可能是造成熱應激缺氧及呼吸性堿中毒的主要原因。目前熱應激引起肺臟細胞損傷的機制尚不明確,但是通過本試驗的觀察與綜合分析可以認為,熱應激對組織的損傷機理與組織的氧化損傷以及細胞內有毒物質的積累有著密切的聯系。在正常生理情況下,體內自由基產生和清除處于動態平衡狀態,當動物受到熱應激時,會使得自由基的產生急劇增加,而自由基清除的能力不足,造成體內自由基蓄積,導致動物的組織氧化受損。
熱應激反應是自然界存在的應激類型中最普遍的一種,從原核生物(如細菌)到人類在高溫環境下均可選擇性合成一類蛋白質,即熱休克蛋白,這類蛋白質參與機體的應激反應,具有多種功能,包括賦予細胞耐熱性以抵抗高溫,作為分子伴侶以防止蛋白聚集,對抗正常的細胞死亡,從而調節細胞的生存和死亡的平衡,現已成為研究應激領域中的熱點方向[4,13]。當動物機體受到外界多種應激刺激時,細胞內的HSPs會被大量合成和消耗,以抵抗外界刺激對機體的損傷,從而維護細胞的正常功能和代謝。大量研究表明,HSPs在抗應激損傷過程中起重要作用,對心臟、肝臟、腎臟等組織器官起保護作用[5,14-15],而HSPs在肺臟內的表達變化規律尚不清楚。HSP27的主要功能是在體內提供耐熱性、細胞保護以及在應激條件下維持細胞存活,它通過不依賴于ATP的伴侶作用抑制蛋白質聚集和穩定部分變性蛋白質,從而確保HSP70復合物的再折疊[15]。HSP60在線粒體蛋白質的運輸和維護以及線粒體DNA的傳遞和復制中起著重要作用[16]。在熱應激條件下,HSC70被動員來保護細胞,并通過修復因應激改變其構象的蛋白質來幫助HSP70發揮其保護功能[17]。HSP72可與其他蛋白結合,從而穩定現有蛋白的聚集,并介導在細胞質和細胞器中新翻譯蛋白的正確折疊[14,18]。細胞受到應激刺激后HSP90在細胞中起到防止蛋白非特異性聚集的作用,保證蛋白質進行正確折疊,維持細胞內穩態[5]。本研究發現熱應激導致雞肺臟組織內HSPs表達量出現明顯變化,其中HSP27和HSP72兩種熱休克蛋白升高最顯著,以HSP72表達變化最大。HSP72在正常肺臟組織中表達量較低,在熱應激后表達量開始顯著升高,在熱應激5 h表達量達到最高,隨后開始降低。HSP27在熱應激1 h后呈現極顯著升高狀態,在整個熱應激過程中表達量一直維持在較高水平。HSP60、HSC70和HSP90在雞肺臟組織中均有較高的表達量,熱應激1 h后HSP60表達量顯著升高,隨著熱應激時間延長表達量開始降低,但仍維持在較高水平,HSC70作為結構型HSPs,熱應激后表達量無明顯變化,HSP90在熱應激后表達量有先降低后升高的趨勢,與對照組相比無顯著差異。研究結果表明熱應激后肺臟組織中HSPs表達量發生明顯變化,HSPs表達量的提高可能與提高組織細胞抗熱應激能力相關。
綜上所述,熱應激能引起雞血清pH值和LDH含量升高,血鈣和血鉀濃度降低,造成肺臟組織嚴重損傷,多種HSPs在熱應激的雞肺臟組織中表達量升高。HSPs在熱應激后高表達可能與機體損傷修復中的保護作用有關。然而,HSPs的確切作用需要進一步研究闡明。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
中老年保健(2021年3期)2021-08-22 06:50:04
昆明醫科大學學報(2021年1期)2021-02-07 01:06:36
現代臨床醫學(2021年1期)2021-01-26 00:56:02
中華養生保健(2020年4期)2020-11-16 01:31:40
中西醫結合肝病雜志(2020年2期)2020-10-27 02:18:50
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
