光強對歐洲舌齒鱸稚魚眼組織基因表達的影響
2021-03-11 05:44:00吳禹濛姜潔明任星月姚安琪宋昌斌閆紅偉
海洋科學 2021年2期
吳禹濛 , 袁 震 , 姜潔明 , 張 磊 , 任星月 姚安琪 宋昌斌,閆紅偉 , 劉 鷹 ,
(1. 大連海洋大學, 遼寧 大連 116023; 2. 設施漁業教育部重點實驗室, 遼寧 大連 116023; 3. 中國科學院半導體研究所, 北京 100083; 4. 青島海洋科學與技術試點國家實驗室, 山東 青島 266237)
光照是影響魚類生長發育中的重要因素, 它對魚類攝食、生長、發育和繁殖等都有一定影響[1-2]。光照的三要素為光照強度、光譜和光周期[3]。不同魚類和魚類發育的不同階段對光照強度的需求均存在差異, 如 Vander 等[4]研究了光照強度對大西洋鱈(Gadus morhua)生長的影響, 結果顯示高光照強度下大西洋鱈生長更快、存活率更高。謝從新等[5]也發現隨著光照強度的增大, 烏鱧(Channa argus)幼魚的攝食強度逐漸減小。一般認為, 大多數海水魚靠視覺進行攝食, 無論其與食物的距離長短, 視覺在捕食過程中均非常關鍵, 在水產養殖中與魚類視覺系統匹配的照明條件可以增加其對餌料的辨識度, 縮短搜索食物時間并最終促進生長和提高存活率[6-7]。研究證實, 光照強度對不同魚類的影響具有差異[8-9], 光照強度過低或過高, 會影響魚類的攝食行為, 從而對生長產生脅迫[10-13]。有研究表明光照強度會影響魚類的視覺系統, 將白化斑馬魚(Danio rerio)和白化虹鱒(Oncorhynchus mykiss)飼養在連續強光照射條件下, 發現其視網膜受到光損傷, 主要表現為感光層細胞的凋亡[14]。即使是對外界光脅迫抵抗能力較高的體色正常的魚類, 如歐洲舌齒鱸(Dicentrarchus labrax)、大西洋鱈和大西洋鮭(Salmo salar), 24 h 持續照明條件下, 它們視網膜的感光層會變薄[15]。因此,魚類在過高的光照強度下, 其視覺會受到損害, 進而有可能影響其攝食行為和生長。
在前人的研究中發現, 在藍光照射下, 金魚(Carassius auratus)的TUNEL 測定結果顯示, 視網膜有大量的細胞凋亡, 另外黑色素濃縮激素(melanin concentration hormone, MCH) 和半胱天冬酶-3(caspase-3)基因表達上調, MCH-R mRNA 的表達隨著藍光暴露和強度的增加而顯著增加, 增加視網膜黑色素聚集, caspase-3 是廣泛用于細胞凋亡的指標,是細胞凋亡的核心作用酶, 以上兩個基因上調表明藍光照射下會對視網膜造成損傷, 這與TUNEL 測定結果相符[16]。而在另一項關于金魚的研究表明, 在綠光處理下更有利于維持視網膜的穩定, 增強視網膜細胞的再生[17]。但光照強度對魚類視網膜內基因表達的影響還未見報道。
歐洲舌齒鱸, 或稱舌齒鱸、狼鱸, 隸屬鱸形目(Perciformes) 、 狼 鱸 科(Moronidae) 、 舌 齒 鱸 屬(Dicentrarchus)[18], 歐洲舌齒鱸的含肉率高、營養價值高、抗病力強、適宜池塘和工廠化循環水養殖[19],是歐洲商業化養殖的第一個非鮭科海水魚類, 是歐洲和地中海區域水產養殖業中的重要經濟魚類[20]。2010 年, 中國科學院海洋研究所劉鷹研究員率先于2010 年將其引種到我國進行人工養殖, 并于2014 年突破其苗種繁育技術[21]。為探究光照強度是否會影響歐洲舌齒鱸稚魚的視覺系統, 本研究以孵化后30 d 的歐洲舌齒鱸作為研究對象, 在白光2.0 W/m2(W 2.0)、1.0 W/m2(W 1.0)和0.3 W/m2(W 0.3)條件下對其進行為期66 d 的養殖實驗。實驗結束后比較了3 組稚魚的體長、濕重和存活率, 根據結果構建了W 2.0 和W 0.3 兩組稚魚眼組織的轉錄組文庫并進行了高通量測序, 以期為查明光照強度對魚類生長和視覺的影響機制提供基礎數據, 并為養殖生產者提供參考。
1 材料和方法
1.1 實驗材料
本實驗所用歐洲舌齒鱸(孵化后30 d, 體長12.52±1.34 mm, 濕重92.35±4.77 mg)由大連富谷水產有限公司提供。
1.2 方法
1.2.1 養殖與光處理
首先將上述歐洲舌齒鱸稚魚隨機放置到 9 個100 L 圓柱形桶(桶高62 cm)中, 每桶養殖密度為700 尾, 養殖周期為66 d。根據魚類生長情況投喂鹵蟲無節幼體(孵化后第30 至40 d)和鹵蟲成體(孵化后第41 至96 d), 每天投喂6~8 次, 每次飽食投喂,投喂前均使用強化劑(50 DE 微囊, 山東省升索漁用飼料研究中心)強化鹵蟲無節幼體。每天各養殖水桶統一換水數次, 并清潔養殖容器, 去除殘餌糞便與死亡幼體。保持各養殖水桶中鹽度為 33, 溫度為18.5~19.5℃溶氧為8 mg/L 以上, pH 7.9~8.1, NH4-N <0.2 mg/L, NO2-N < 0.05 mg/L。
本實驗采用人工LED 光源(深圳市超頻三科技股份有限公司)照明, 共設置3 種不同的光照處理組(3種光照強度)來開展養殖實驗, 包括0.3 W/m2、1.0 W/m2和2.0 W/m2的白光(表示為: W 0.3, W 1.0, W 2.0)。由可調節光強的3 盞LED 燈提供光照, 每個光照處理組設置3 個養殖平行(共在9 個養殖水桶中開展實驗)。采用不透光的灰色幕布將2 個處理組進行隔離并防止外界光線的影響, 且每天使用光譜輻射分析儀(PLA-20, 杭州遠方光電信息股份有限公司)在水面上進行測試一次, 確保實驗期間光環境的穩定。
1.2.2 生長數據計算
在實驗結束時隨機每組選取20 尾稚魚, 冰上低溫麻醉后, 測定其體長和濕重。每天計數表面和底部死亡魚的個數, 存活率的測定參考Yan 等的研究[22]。
1.2.3 RNA 提取
根據生長數據結果, 為保證試驗數據可靠性,另取W 0.3 和W 2.0 組的稚魚麻醉(20 尾/桶), 于低溫條件下分離眼組織, 放入裝有RNA later 的離心管中,之后在–80℃超低溫冰箱中進行保存。眼組織的總RNA 的提取采用RNeasy Mini Kit 試劑盒(Qiagen, 德國), 根據說明書操作。采用Agilent 2100 生物分析儀和Nanodrop ND-1000 分光光度計檢測RNA 的質量和濃度。
1.2.4 基因文庫構建及轉錄組分析
樣品檢測合格后, 將之前提取的總RNA 等量混合, 送樣至北京諾禾致源生物有限公司進行文庫構建和轉錄組測序。
測序產生的數據(reads)作為原始數據(raw data),將帶接頭的、低質量的reads 過濾除去進而得到高質量可用數據(clean reads)。利用組裝軟件Trinity 對獲得的高質量測序數據進行序列組裝。
通過BLASTX 將轉錄本與NCBI、SwissProt、KEGG、GO、COG、KOG、EggNOG、Pfam9 個數據庫比對。利用Blast2 GO 軟件進行基因本體(GO)注釋, 并使用TopGo 進行富集分析。序列也進一步與COG 和EggNOG 數據庫進行比對, 并對基因序列進行功能預測和功能分類。利用 Perl script 進行KEGG 通路的富集對基因產物在細胞中的功能及其代謝途徑進行系統分析。通過在線KEGG 自動注釋服務(KAAS), 可以得到的每條序列的京都基因與基因組同源體系分析(KO)注釋, 并映射到相應的KEGG 通路中。
1.2.5 差異表達基因的鑒定
利用FPKM 值表示對應非重復序列基因(unigene)的表達豐度。FPKM 計算公式如下:
每一百萬個map 上的reads 中map 到外顯子的每1K 堿基上的Fragments 個數(FPKM)=cDNA 片段(cDNA fragments)/圖譜片段(百萬)[mapped fragments(millions)]/轉錄長度[transcript lengths (kb)]。
篩選差異表達基因的標準為錯誤發現率(False Discovery Rate, FDR)≤0.01 和差異倍數(log2fold change, FC)>1。
1.2.6 實時定量PCR
為了驗證轉錄組數據的正確性, 采用實時定量PCR 檢測晶狀體纖維主要固有蛋白、視黃醇結合蛋白3、維生素A 脫氫異構酶、熱休克同源70、伸長因子1-α 和葡萄糖-6-磷酸異構酶6 個基因的表達。引物使用Primer Premier 5.0 軟件進行設計(表1)。以提取的眼組織的 RNA 為模板, 按照 SYBR FAST qPCR Kit Master Mix(2×)試劑盒說明書合成cDNA,然后以其為模板, 以β-actin 為內參基因進行實時定量PCR 擴增(Applied Biosystems 7900 HT Real-TimePCR儀)。PCR 條件為: 95℃, 5 min; 95℃, 3 s 和60℃, 20 s共40 個循環[23]。

表1 引物序列Tab. 1 Sequences of the primers used for PCR amplification of differentially expressed genes in Dicentrarchus labrax
1.2.7 統計分析
實時熒光定量數據以2–ΔΔCT法處理。采用統計軟件IBM SPSS 22.0(IBM, Armonk, NY, 美國)中的t檢驗分析眼內的基因表達的顯著性差異, 使用Duncan 檢驗兩個不同光照組之間稚魚體長、濕重和存活率, 顯著性設定為P<0.05 和P<0.01。
2 結果
2.1 不同光強的白光對歐洲舌齒鱸稚魚生長的影響
在實驗結束時, W 2.0 組稚魚的體長和濕重顯著小于W 0.3 和W 1.0 處理組下飼養的稚魚(P< 0.05),但三組歐洲舌齒鱸稚魚的存活率無顯著性差異(P>0.05)(圖1)。
2.2 轉錄組測序與組裝
經Illumina Hiseq 2500 測序, W 0.3 處理組獲得raw reads(統計原始序列數據) 50 907 730 個, W 2.0處理組獲得raw reads 50 265 736 個; W 0.3 處理組獲得clean reads 50 273 392 個, W 2.0 處理組獲得clean reads 49 277 200 個; W 0.3 處理組獲得clean bases(測序序列的個數乘以測序序列的長度) 7.54 GB, W 2.0處理組獲得clean bases 7.39 GB。
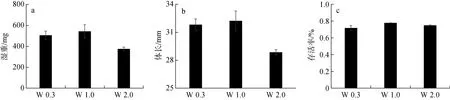
圖1 W 0.3、W 1.0 與W 2.0 組歐洲舌齒鱸稚魚的濕重、體長和存活率Fig. 1 Wet weight, body length, and survival rate of Dicentrarchus labrax juveniles in the W 0.3, W 1.0, and W 2.0 groups
2.3 差異表達基因篩選及其功能富集分析
根據兩個RNA 樣本基因的差異表達豐度來做差異表達分析。如圖2 所示, 共得到的差異表達基因有368 個, 與W 0.3 組相比, W 2.0 組中234 個基因上調表達, 134 個基因下調表達(圖2)。
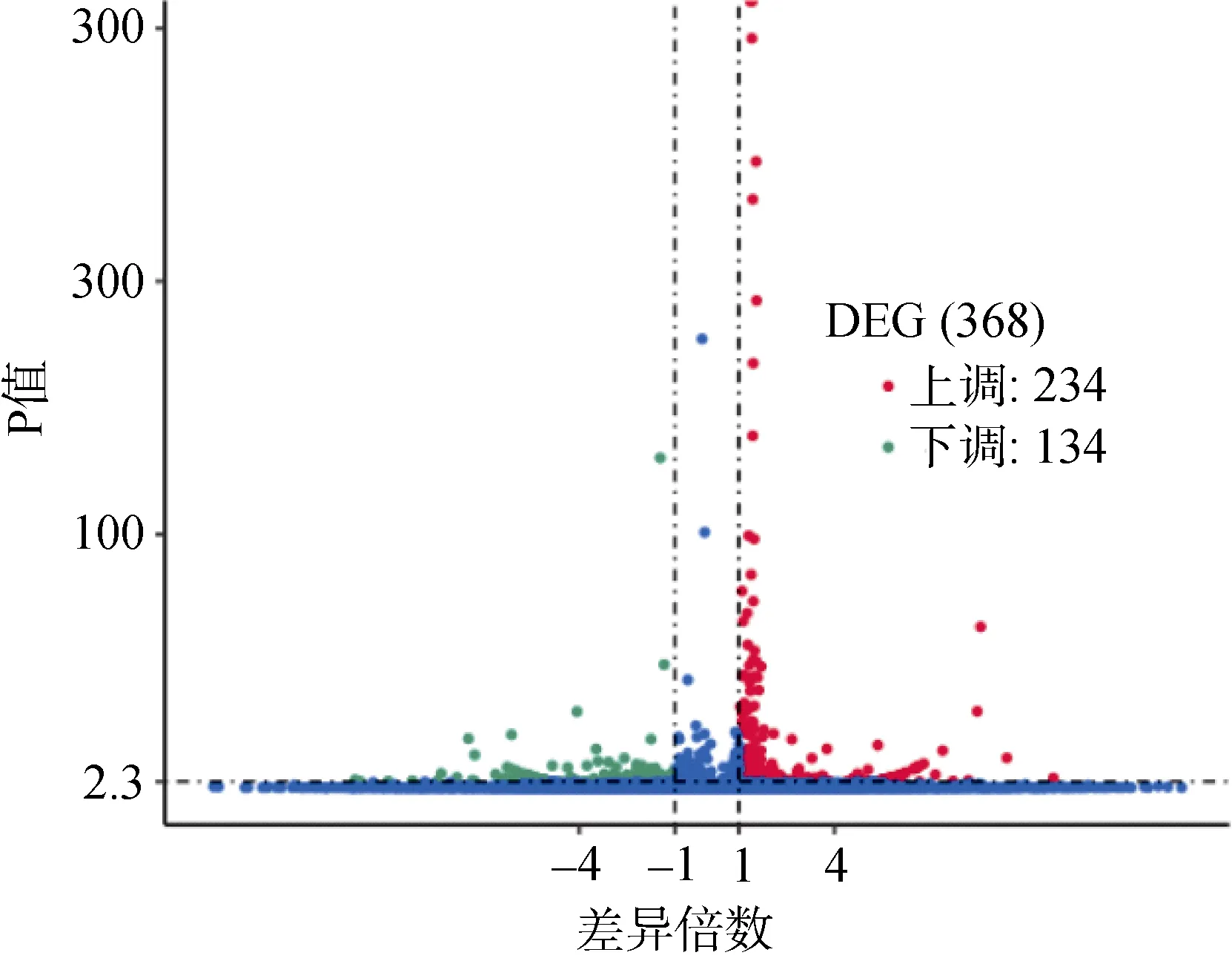
圖2 W 0.3 與W 2.0 組歐洲舌齒鱸稚魚眼內差異表達基因的火山圖Fig. 2 Volcano plot of differentially expressed genes in the eye of D. labrax between the W 0.3 and W 2.0 groups
在篩選得到的差異表達基因中, 晶狀體纖維主要固有樣蛋白(MIP),晶狀體纖維膜固有蛋白(lens fiber membrane intrinsic protein), 晶狀體纖維膜固有蛋白(lens fiber membrane intrinsic protein), 晶狀體纖維膜固有蛋白樣亞型X1(lens fiber membrane intrinsic protein-like isoform X1)等, rho 相關的btb 結構域, 包含蛋白 2 樣亞型 X1(rho-related BTB domain-containing protein 2-like isoform X1), 胰島素樣生長因子結合蛋白1(insulin-like growth factor-binding protein 1), 維生素A 脫氫異構酶(RPE65)等均在W 2.0 組出現了上調(表2)。
對得到的368 個差異表達基因進行GO 分析發現, 差異表達基因主要富集在分子功能(MF)和生物學功能(BP)上。其中在MF 上差異表達基因主要分布在細胞黏附(cell adhesion)、生物黏附(biological adhesion)、磷酸化(phosphorylation)等通路上。而在BP上差異表達基因主要分布在葡萄糖-6-磷酸異構酶激(glucose-6-phosphate isomerase activation)、分子內氧化還原酶(intramolecular oxidoreductase), 且分布較少(圖3)。
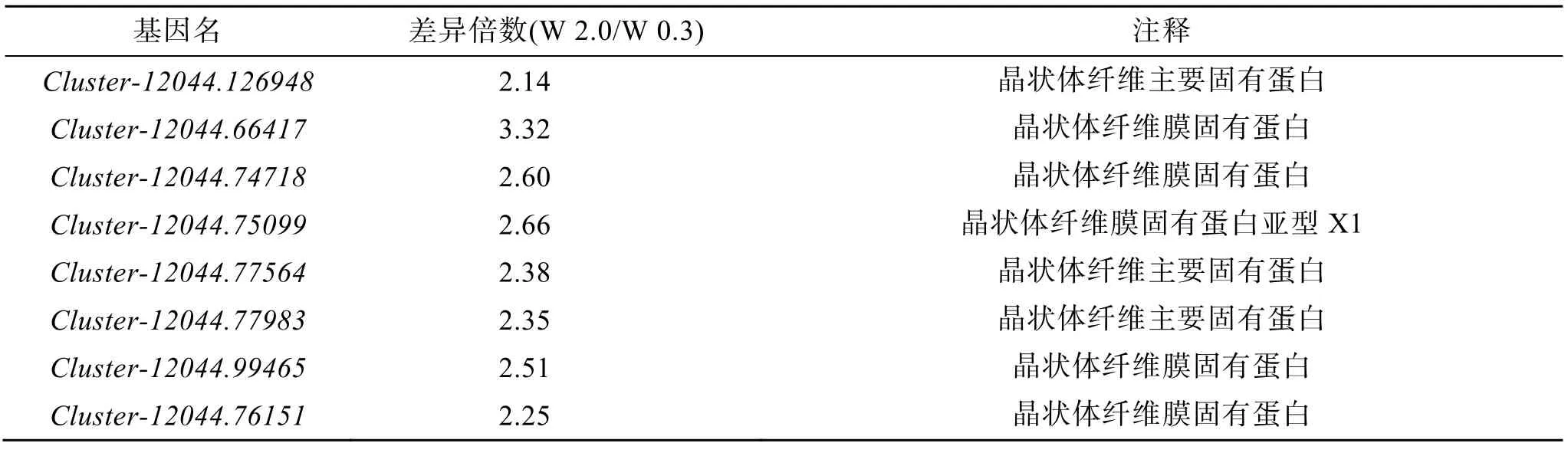
表2 W 0.3 與W 2.0 組歐洲舌齒鱸眼內代表性的差異表達基因Tab. 2 Differentially expressed genes in the eye of D. labrax between the W 0.3 and W 2.0 groups
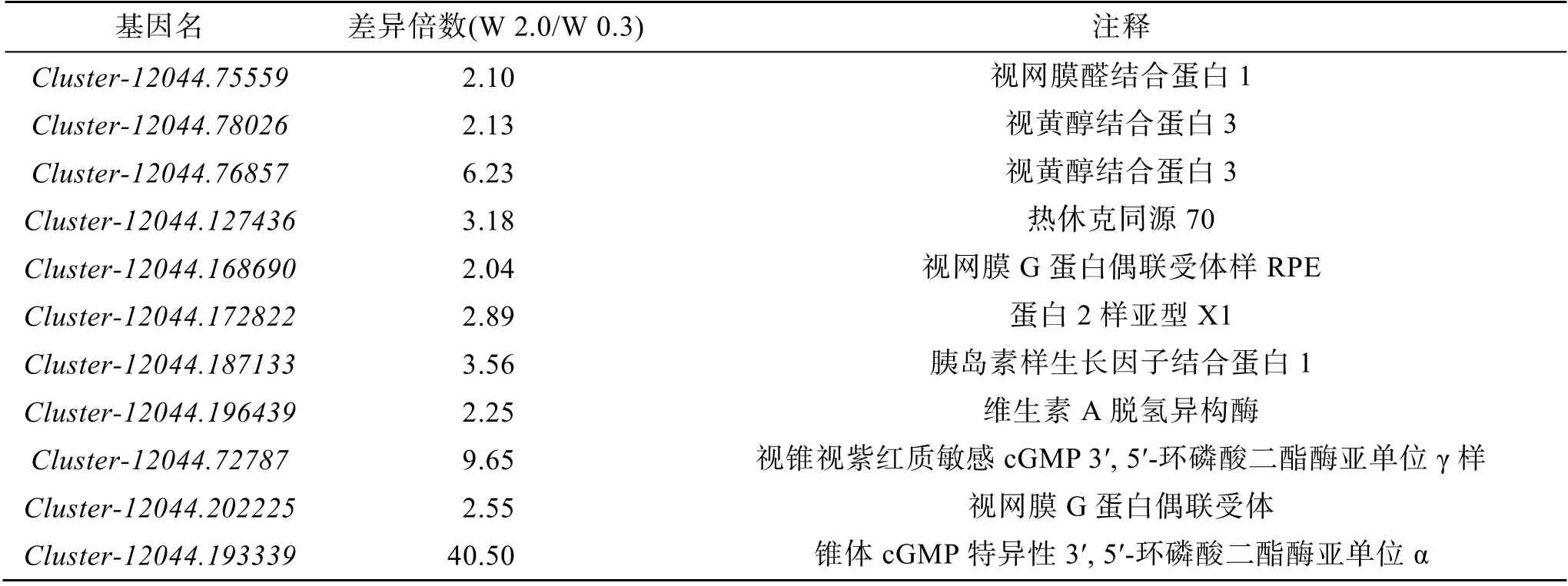
續表
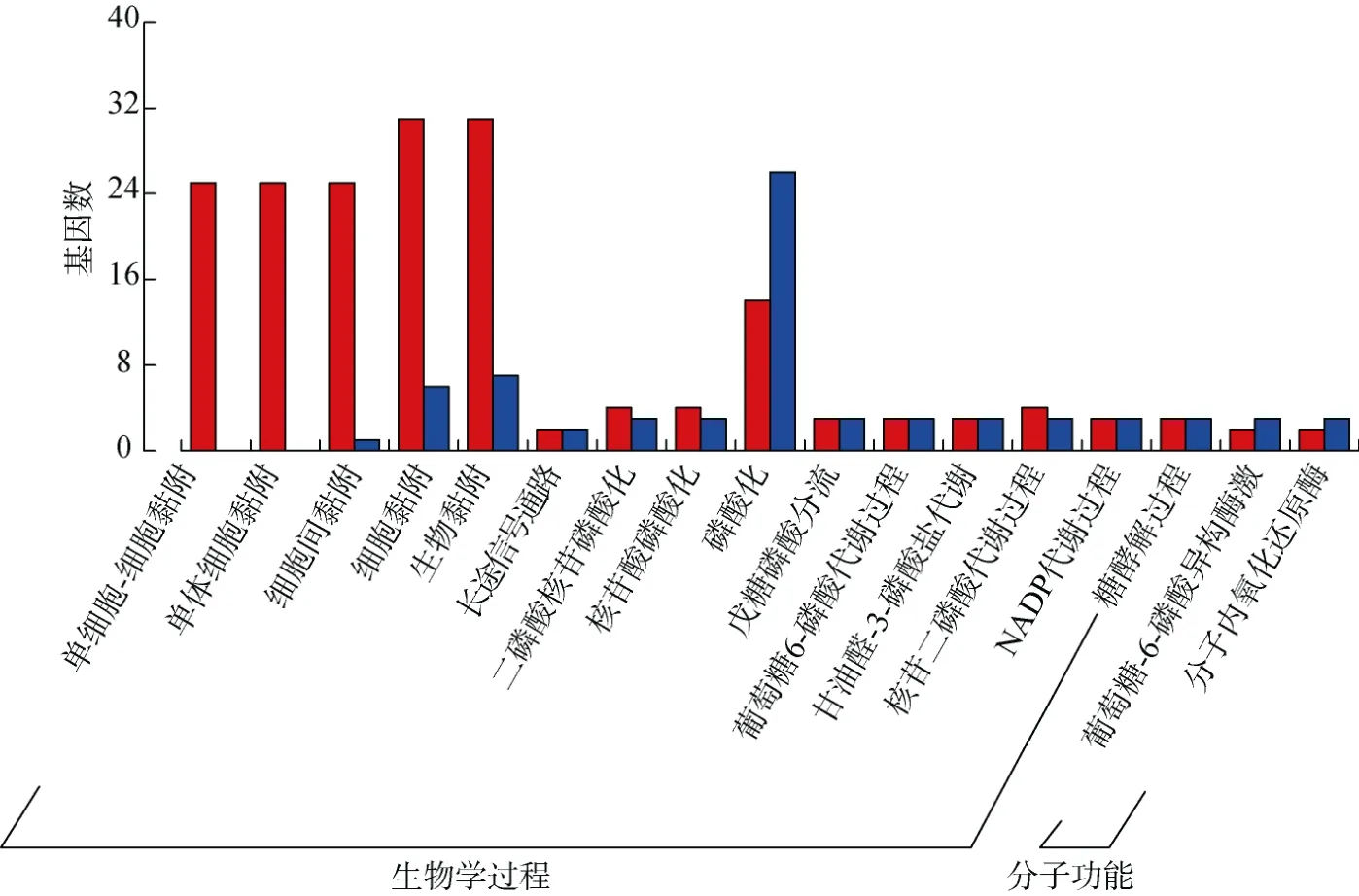
圖3 W 0.3 與W 2.0 組歐洲舌齒鱸眼內差異表達基因的GO 富集分析Fig. 3 GO enrichment analysis of differentially expressed genes in the eye of D. labrax between the W 0.3 and W 2.0 groups
KEGG富集分析結果如圖4所示, 差異基因富集在碳代謝(carbon metabolism), 乙醛酸鹽和二羧酸鹽代謝(glyoxylate and dicarboxylate metabolism), RNA轉運(RNA transport), 氧化磷酸化(oxidative phosphorylation), 三羧酸循環 TCA 循環(citrate cycle TCA cycle), 淀粉和蔗糖的代謝(starch and sucrose metabolism), 軍團桿菌病(legionellosis), 磷酸戊糖途徑(pentose phosphate pathway)等20 個通路上。在這些通路中, RNA 轉運、核糖體、氧化磷酸化和碳代謝上差異表達基因的分布相對較多。
2.4 qPCR 結果
對得到的6 個差異表達基因進行驗證, 結果顯示在W 2.0 組稚魚眼內MIP、HSC70、 RBP3、RPE65、EF1A 和GPI 基因的表達都顯著高于W 0.3 組稚魚(P<0.05, 圖5), 這與轉錄組結果一致。
3 討論
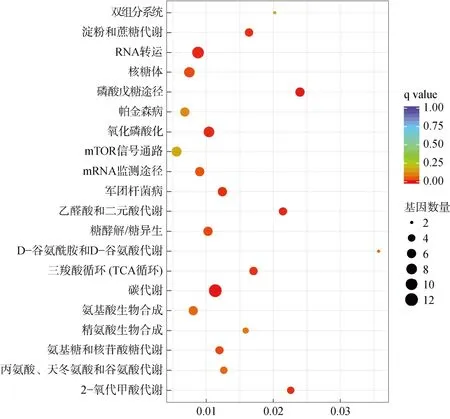
圖4 W 0.3 與W 2.0 組歐洲舌齒鱸眼內差異表達基因的KEGG 富集分析Fig. 4 KEGG enrichment analysis of differentially expressed genes in the eye of D. labrax between the W 0.3 and W 2.0 groups
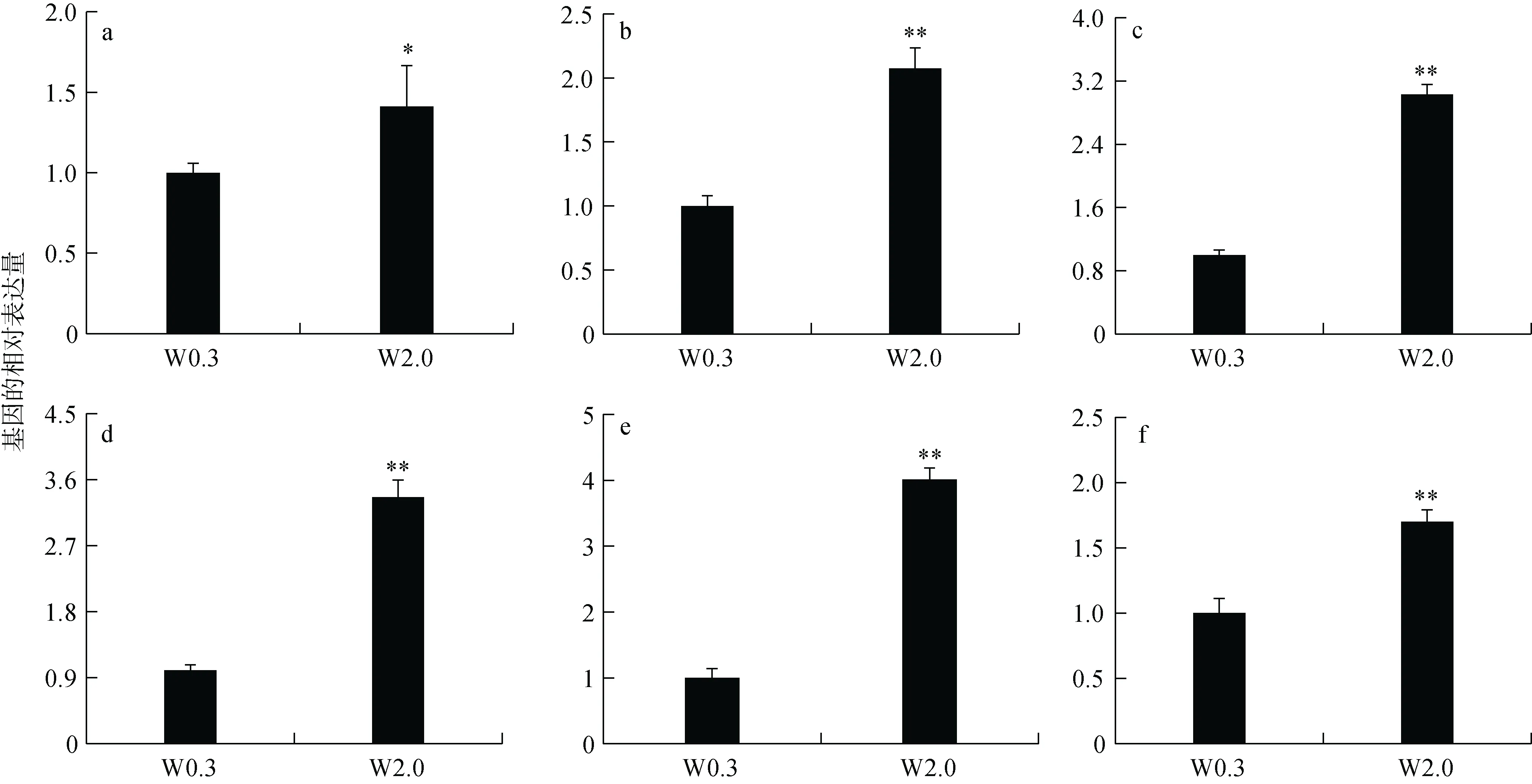
圖5 W 0.3 與W 2.0 組歐洲舌齒鱸眼內6 個基因表達Fig. 5 Expression of six genes in the eye of D. labrax between the W 0.3 and W 2.0 groups
許多海洋魚類靠視覺覓食, 并且需要一個最低光照強度閾值; 但是對高光強影響稚魚視覺, 進而影響其攝食和生長, 目前關于這方面的認識仍然不足。前人的研究主要集中在光照條件對眼睛, 尤其是視網膜的組織學上的觀察, 而對眼內基因的相關性表達產生的影響的報道少之又少。Bayarri 等人研究發現影響歐洲舌齒鱸眼睛視網膜敏感性光閾值為0.06 W/m2[24], Villamizar 等人設置光照強度為0.42 W/m2,使歐洲舌齒鱸眼睛和血漿中褪黑激素含量產生變化[25]。在本研究中, 我們的研究任務是查明當光照強度增加時, 歐洲舌齒鱸稚魚的生長、存活和視覺是否受到影響。因此我們將稚魚分別暴露于0.3 W/m2、1.0 W/m2和2.0 W/m2這3 個光照強度下, 形成3 個處理組。實驗結束時, 在光照強度為2.0 W/m2下飼養的稚魚的平均體長、濕重均顯著小于1.0 或0.3 W/m2下飼養的稚魚(P<0.05), 而1.0 和0.3 W/m2光照強度下飼養的稚魚平均體長無顯著差異(P>0.05), 3 個處理組之間的存活率無顯著性差異(P>0.05)。前人研究證明, 漠斑牙鲆(Paralichthys lethostigma)暴露于100 lx的光照強度以下時生長和存活率都有所降低[26]。金頭鯛(Sparus aurata)在中等強度下生長較好, 可能是由于這種光強下會刺激視網膜上的色素, 增加其辨別獵物的能力[27]。加州鱸魚(yellow perch)的反應距離,隨著光強的降低而逐漸降低, 當光強度下降到小于2 lx 時, 最大平均反應距離顯著下降, 在黑暗中最小反應距離減小[28]。另在禽類中, 光照強度在1~65 lx,對雞的生長沒有影響, 而在130~290 lx 的光照下會抑制雞的生長[29]。由此可見, 因不同物種的生活環境不同, 其對光強的適應范圍各有不同, 過低或者是過高的光強都會對其生長產生影響。因此, 在養殖生產中, 了解魚類生長所需的合理光照強度至關重要。
一般認為大部分海水魚是通過視覺來攝食, 在W 2.0 組稚魚的生長較差, 可能是因為2.0 W/m2對魚類來說屬于高光強, 高光強可能會影響其視覺, 進而對其生長產生影響。與高等脊椎動物相比, 多數魚類的眼睛無眼瞼, 不能調節瞳孔大小來控制入射視網膜的光強度, 所以在高光強下魚類的視網膜更容易受到傷害[30]。研究表明, 將大西洋鱈、大西洋鮭和歐洲舌齒鱸置于24 h 高光強照明條件下, 它們的視網膜會受到損傷[15]。我們前期研究中發現, 24 h 連續照明條件下, 歐洲舌齒鱸稚魚的視網膜各層厚度會受到影響, 且感光層均會受到不同程度的損傷, 相比W 0.3 組, W 2.0 組稚魚的視網膜感光層受損更為嚴重, 除視桿細胞的損傷外, 我們還發現視錐細胞外節出現縮短的現象, 在腫脹的內節里出現細胞核溶解, 這是一種細胞壞死的標志[22]。研究發現, 光誘導的光感受器細胞損傷是從外節末端開始的, 這表明外節層是最先遭受損傷的, 損傷的嚴重程度和膜盤的更新速度有關[31]。趙穎熙研究發現, 高光強可以引起豚鼠眼球眼軸增長減緩, 對視網膜光感受器造成輕微損傷[32]。然而, 即使在哺乳動物中, 光照對視網膜造成的損傷的分子機制依舊不是十分清楚。
在本研究中, 以高通量測序為研究手段, 構建了W 0.3 和W 2.0 兩組稚魚的眼轉錄組文庫, 并篩選不同處理組之間的差異表達基因, 以期從分子水平來解釋光照強度對歐洲舌齒鱸視覺的影響。結果表明, 共獲得在W 2.0 和W 0.3 組稚魚眼內差異表達的基因368 個, 與W 0.3 組相比, W 2.0 組中234 個基因上調表達, 134 個基因下調表達。在這些差異表達基因中, 我們發現, MIP、RBP、RPE65、HSC70、EF1A和GPI 6 個基因均在W 2.0 組高表達。這說明, 這些基因的上調可能是對光照強度的響應, 進而影響歐洲舌齒鱸稚魚的視覺。
其中我們篩選得到的MIP 為水通道蛋白的一種,主要在晶狀體纖維細胞及視網膜中表達[33]。截至目前, 已經在植物、原生生物和脊椎動物中發現超過200 種水通道蛋白[33]。眾所周知, 人體內大部分器官,如肝、腎、眼等依賴于滲透壓調節來維持各自的生理功能, 所以水分對傳遞視覺信息, 視網膜成像起著至關重要的作用[33]。MIP 同時也具有結構性鏈接功能, 它與晶狀體正常代謝及透明度的維持密切相關。近年來的研究發現, MIP 也與白內障的發生密切相關[33]。而白內障是由于晶狀體病變而引起的疾病,晶狀體在正常狀態下是透明的, 因不同原因引起的晶狀體蛋白變性、水腫、纖維之間出現空泡, 上皮細胞增生等, 從而使晶狀體透明性減弱, 阻礙光線入射眼內, 形成白內障, 進而影響視力[34-35]。HSC70 是1999 年Ballinger 等[36]發現的具有輔助伴侶分子和泛素連接酶功能的蛋白質, 可以通過其氨基端連接熱休克蛋白調節蛋白質的重新折疊, 同時通過其羧基端連接蛋白酶體促進底物的降解[37]。HSC70 在所有的器官內幾乎均有表達, 但在代謝效率較高亦或是蛋白質更替較快的器官或組織中, 如骨骼肌、心臟和腦中高表達, 在胰腺、肺、肝、胎盤和腎臟中的表達水平相對較低[37]。在正常情況下, HSC70 參與維持蛋白質的空間構象, 起到細胞骨架等基本功能。可當細胞面臨脅迫時, 生物體會大量表達HSC70 用于阻止變性蛋白質的的積累, 增加細胞的抗逆和保護機制[38]。維生素A 由視黃醇(維生素A1)和3-脫氫視黃醇(維生素A2)組成, 兩者均為20 碳的白芷酮環多烯烴一元醇, 在動物體內, 其多存在于肝臟中[39]。在前人的研究中發現, 維生素A 是構成視覺細胞內感光物質的成分, 是維持上皮組織健全所必須的物質,其不足時會引起維生素A 缺乏綜合征, 如眼部視網膜血管炎[40]、夜盲癥和干眼癥[41]。在我們篩選的基因中, 我們發現, RBP3 和RPE65 兩者均在W 2.0 組稚魚高表達, 而RBP 是維生素A 的運載蛋白, 在協助維生素發揮生理功能中起著不可替代的作用[42]。綜上, 與W 0.3 組相比, 在W 2.0 組中, 上述基因在眼睛組織中的表達水平上升, 可能是因為該光照條件對稚魚視覺系統造成了脅迫, 其作用機制尚需進一步研究。GPI 又被稱為磷酸葡萄糖異構酶, 在所有真核生物和原核生物的細胞中普遍存在, 是一類具有多功能生物活性的天然蛋白質[43], 它參與糖代謝的糖酵解作用[44]。光電能的轉換是視網膜組織的重要功能之一, 這種轉換過程需消耗大量的能量。這種能量的來源主要是依靠葡萄糖的酵解獲得[45]。因此,W 2.0 組稚魚的光電能的轉換速率可能增加, 進而使葡萄糖-6-磷酸異構酶表達水平升高。
4 結論
本研究以歐洲舌齒鱸稚魚為研究對象, 在白光光照強度為2.0 W/m2、1.0 W/m2和0.3 W/m2的條件下對其進行養殖。實驗結束后比較了3 組稚魚的體長、濕重和存活率, 且構建了W 0.3 和W 2.0 兩組稚魚眼組織的轉錄組文庫并進行了高通量測序。研究發現白光飼養條件下, 高光照強度會抑制歐洲舌齒鱸稚魚的生長, 這可能是由于高光照強度會對其視覺產生影響進而影響其攝食活動和生長。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
