微生物響應PBAT-PLA生物降解膜袋工業需氧堆肥降解機制
2021-03-17 07:48:18馬昳超章若紅朱佳歡杜歡政
農業工程學報 2021年24期
關鍵詞:生物
馬昳超,劉 峻,章若紅,朱佳歡,杜歡政,陸 莎
微生物響應PBAT-PLA生物降解膜袋工業需氧堆肥降解機制
馬昳超1,2,劉 峻1※,章若紅1,朱佳歡1,杜歡政2,陸 莎2
(1. 上海市質量監督檢驗技術研究院,上海 201114;2. 同濟大學環境科學與工程學院,上海 200086)
塑料污染已對全球環境造成嚴重威脅,生物降解塑料的推廣使用及其工業堆肥是治理塑料污染的有效途徑之一。該研究根據GB/T 19277.1—2011,在(58±2)℃的特殊高溫條件下,對PBAT-PLA生物降解膜袋進行有氧堆肥降解,并選擇微晶纖維素作為對照。通過對堆肥中的微生物進行16S/18S高通量測序,分析降解過程中細菌/真菌的群落種類和數量變化,包括物種多樣性、物種組成、物種差異分析、樣本比較分析,并結合掃描電鏡下的微觀形貌,深入探尋可降解塑料膜袋在工業需氧堆肥過程中的微生物響應降解機制。結果表明:微晶纖維素和生物降解膜袋在降解活躍期(第140天取樣),其所在堆肥中大量存在的優勢菌屬為(球桿菌屬,放線菌綱),分別占比20.25%和39.44%。與同樣條件下不含降解材料的對照組堆肥相比,微晶纖維素/生物降解膜袋工業需氧堆肥降解過程中顯著增長的4種菌屬中有3種屬于放線菌,說明放線菌對聚酯物的解聚以及纖維素的降解具有積極的作用。試驗結果也表明了聚酯和纖維素的完整生物降解過程不依賴單一菌種,而是微生物協同作用的結果。
微生物;塑料;降解;生物降解膜袋;工業需氧堆肥;高通量測序
0 引 言
自20世紀50年代以來,全球共生產了83億t塑料,其中63億t成為塑料垃圾。目前全球年均使用5 000億個塑料袋,并造成至少800萬t塑料進入海洋[1]。2020年1月19日中國公布《關于進一步加強塑料污染治理的意見》,要求積極推廣可循環、易回收、可降解的替代產品,規范塑料廢棄物的回收利用。降解產品,特別是生物降解塑料制品的使用是解決部分一次性塑料制品污染的有效方案。為了完善生物降解塑料制品的檢測標準并實施監管,中國針對不同降解環境,也發布了一系列的可降解檢測相關國標。其中使用較為廣泛的是標準《受控堆肥條件下材料最終需氧生物分解能力的測定—采用測定釋放的二氧化碳的方法第1部分:通用方法:GB/T 19277.1—2011》[2]。通過受控堆肥處理固體廢物是處理和回收有機廢物材料的一種有價值的方法[3]。可生物降解塑料的堆肥是廢品回收的一種形式,可減少對填埋場日益增長的需求,但其降解過程會對堆肥產生一定的影響,且其降解產物會影響食物鏈[4]。因此,研究生物降解塑料的降解機制及其環境微生物響應具有重要的意義。
生物降解塑料是指在細菌、真菌、藻類等自然界存在的微生物作用下發生生化、物理作用而降解或分解的材料[5]。理想的生物降解塑料在廢棄后最終可被環境微生物完全分解并生成CO2和水,進入自然生物圈的物質循環系統,不再對生態環境造成危害[3]。近年來,生物降解塑料的降解機理一直是國內外的研究熱點[6]。Haider等[7]指出生物降解塑料的降解過程主要包括生物腐蝕,胞外酶的解聚合,微生物同化降解及礦化。Gu等[8]總結了幾種聚合物材料降解中涉及的機理和微生物,長鏈聚合物由于分子較大,難以通過微生物細胞膜。在降解過程中,來自微生物的胞外酶分解長鏈聚合物,產生短鏈或更小分子的聚合物(例如低聚物、二聚體和單體)的過程叫做解聚合。解聚合過程使聚合物分子變小,部分小分子化合物可以由細胞膜的主動運輸等吸收作用進入胞內。這些短鏈分子可以作為碳源供細胞生長代謝,并最終以CO2、H2O或CH4等代謝物的形式排放到胞外。Weng等[9]將聚乳酸(Polylactic Acid,PLA)、聚己二酸/對苯二甲酸丁二醇酯(Poly Butylene Adipate-co-terephthalate,PBAT)和PBAT/PLA薄膜樣品埋在真實的土壤環境中。定期從土壤中采集殘留降解樣品,并通過掃描電鏡、差示掃描量熱法、熱重分析、紅外和元素分析的表征探尋降解機理。Bonilla等[10]采用OECD 301d標準,對殼聚糖、PBAT和高密度聚乙烯(High Density Polyethylene,HDPE)聚合物進行了生物降解試驗。并通過記錄照片,紅外,掃描電鏡,彈性模量和拉伸強度等指標研究其降解規律與機理。Zhang等[11]發現PLA-PBAT農膜在不同地區土壤中的降解程度不同,同時可降解塑料農膜的使用會改變土壤微生物群落。生物降解的過程復雜而多樣化,受溫度、濕度、菌群種類等因素影響。然而對目前最常使用的受控工業需氧堆肥(58±2)℃特殊條件下的降解機制研究尚為空白。
本研究根據GB/T 19277.1—2011,在受控工業需氧堆肥條件下,對微晶纖維素(作為參比)、生物降解膜袋實施降解。通過對堆肥中的細菌/真菌進行16S/18S高通量測序,來分析降解過程中細菌/真菌的群落變化,包括物種多樣性、物種形成、物種差異分析、樣本比較分析,并結合掃描電鏡下的微觀形貌分析,深入探尋可降解塑料膜袋的微生物響應降解機理。為今后優化工業堆肥技術,提升可降解塑料檢測標準打下理論基礎,更為今后生物降解塑料膜袋大面積推廣,及其堆肥產品在新型功能農業領域應用的可行性研究提供科學依據。
1 材料與方法
1.1 試驗材料
生物降解膜袋(無印良品)主要基材為65% PBAT、5% PLA,并含有30%滑石粉。生物降解率測試所用參比物為微晶纖維素,購于國藥試劑(品牌sigma-aldrich,產品號S3504,CAS 9004-34-6,粒徑20m,色譜級)。有機堆肥購于上海質檢院合作堆肥廠(堆肥粒徑<0.5 cm,碳氮比10~40,干固體比例68.7%,揮發性固體比例27.75%)。
1.2 受控工業堆肥需氧生物分解率測試
材料在受控工業堆肥條件下最終需氧的生物分解能力依據國標GB/T 19277.1—2011測定,采用測定釋放CO2的方法,在上海市質量監督檢驗技術研究院進行測試。降解測試設備包含生物降解反應系統與紅外在線式氣體分析系統兩大模塊,由SQI搭建。在工業需氧堆肥(58±2)℃條件下,按6:1把干基質量為300 g的培養土和干基質量為50 g的PBAT-PLA膜袋或微晶纖維素放入2 L降解反應容器,測定材料最終需氧生物分解能力[12],每個樣品平行測試3次。微晶纖維素作為參比材料,可驗證堆肥環境的降解適宜性。由于原堆肥中本身含有機碳源,因此還需進行對照組堆肥降解測試,用以扣除堆肥中自帶碳源的影響。定期關注降解反應容器中堆肥混合物的濕度,若容器頂部無冷凝水,則需要從瓶口加水調節水份,使堆肥含水量保持在約50%。使用紅外在線式氣體分析系統對反應容器中產生的CO2進行連續監測,并累計所產生的CO2量。由式(1)可得,降解材料實際產生的CO2量與該材料理論CO2的產生量之比為生物分解百分率D(%)。
式中(CO2)T為每個含有降解材料混合物的降解測試容器累計釋放出的CO2量,g;(CO2)B為3個對照組測試容器累計放出的CO2量平均值,g;ThCO2為降解材料產生的CO2理論釋放量,g。
1.3 環境微生物群落多樣性測序及交互式分析
1.3.1 堆肥樣品取樣與DNA抽提
選取生物降解率曲線趨于平穩前的最后時間點,此時生物分解率測試已基本完成,而微生物所需的碳源依然充足,仍處于降解活躍期。分別提取原樣品、對照組、纖維素堆肥以及生物降解膜袋堆肥,標記為堆肥A、B、C、D,詳細的堆肥樣品信息見表1,每個樣品做3個平行試驗。取樣前將反應器充分混勻,使用滅菌藥匙取1~2 g堆肥樣品,裝入無菌容器,干冰冷藏條件下送檢。DNA抽提選用土壤DNA抽提試劑盒(FastDNA? Spin Kit for Soil 116560-200,美國MP)。

表1 堆肥樣品信息
1.3.2 16S rDNA和18S rDNA高通量測序
16S rDNA和18S rDNA高通量測序由上海美吉生物有限公司使用MiSeq PE300平臺(Illumina,美國)進行。16S rDNA使用引物338F(5′-ACTCCTACGGGAGGCAGCAG- 3′)和806R(5′-GGACTACHVGGGTWTCTAAT-3′)擴增每個樣本中細菌16S rRNA的基因[13]。使用引物SSU0817F(5′-TTAGCATGGAATAATRRAATAGGA-3′)和1196R(5′-TCTGGACCTGGTGAGTTTCC-3′)擴增真菌18S rRNA基因[14]。
1.3.3 生物信息分析
原始序列使用USEARCH 7.1進行質量控制程序[15]。使用 UPARSE 7.0.1090軟件將具有97%或更高相似性的序列聚類為操作分類單元(OUT)[16]。使用包含細菌和真菌rRNA序列的SILVA數據庫(版本138)對序列進行分類[17]。使用美吉生信云平臺從Alpha多樣性水平評估微生物多樣性,包括豐富度(Chao,Ace)、多樣性(Shannon,Simpson)和測序覆蓋率(Coverage)。進一步分析屬水平下堆肥中細菌/真菌組成,使用Student’s T檢驗比較兩組間的差異,并使用Beta多樣性(主坐標分析)比較堆肥樣品的差異性[15]。
1.4 掃描電鏡
掃描電子顯微鏡圖像由掃描電鏡(FEI,FP Quanta 250)拍攝。工業堆肥降解后的微晶纖維素與堆肥充分混合,難以直接分離。取纖維素和堆肥混合樣品置于去離子水中,攪勻并靜置,降解后的微晶纖維素,由于分子鍵的斷裂,分子量減小,因此會漂浮于水面。靜置后取上層液體,滴于鋁箔紙,放于通風櫥中自然風干后待測。夾取降解后堆肥中的生物膜袋碎片,使用去離子水漂洗去除膜袋表面堆肥后,將膜袋碎片鋪于鋁箔紙上,然后放入通風櫥自然風干。經過預處理的微晶纖維素和生物膜袋樣品分別鍍金,在20.00 kV,放大3 000倍條件下進行掃描電鏡觀察[18]。
2 結果與討論
2.1 受控需氧工業堆肥條件下膜袋生物分解率
生物降解膜袋樣品與微晶纖維素的生物分解百分率曲線如圖1所示。試驗初期,纖維素的生物分解速度較PBAT-PLA生物降解膜袋的分解速度更快。第60天時,纖維素的生物分解率達到76.6%±2.9%,隨后其降解速度放緩。在第90 天時,膜袋和纖維素的生物分解率分別為81.1%±4.1%和80.0%±2.7%,隨后兩者的分解率開始趨于一致,PBAT-PLA膜袋的分解率略高于纖維素。第120 天時,PBAT-PLA膜袋和微晶纖維素的生物分解率分別為91.4%±3.4%和87.5%±2.0%,繼續培養至第150 天,分別達到98.8%±1.8%和95.1%±2.2%。因此,本研究選擇在第140 天根據表1分別提取堆肥樣品,進行16S rDNA和18S rDNA高通量測序,此時膜袋和纖維素的生物分解率已接近最高值,但微生物生存所需碳源還沒有完全耗盡,微生物依然保持活性,因此對這一時間段的微生物群落進行分析對研究工業堆肥中的生物降解塑料降解機制具有重要價值。
2.2 堆肥中微生物群落多樣性分析
Alpha多樣性反映了堆肥反應體系內微生物群落的多樣性,常用的度量標準有Shannon、Simpson、Chao、Ace、Coverage,堆肥中細菌和真菌的Alpha多樣性指數見表2。所有樣品的群落覆蓋度(coverage)均在0.999以上,表明樣品測序結果能夠反映實際情況。Shannon和Simpson指數可表示堆肥樣品中微生物群落的多樣性。Shannon值越大,群落多樣性越高;Simpson 指數值越大,群落多樣性越低[19]。由表2中的Shannon和Simpson指數可見,堆肥B的細菌多樣性顯著低于原堆肥A。這是因為工業堆肥(58±2)℃的特殊環境下,只有部分嗜熱菌能夠生存。同時由于降解周期長達150 d,堆肥中的能量來源逐漸被消耗,造成一部分細菌的死亡。而堆肥C和D中,由于加入了纖維素或生物降解購物袋,為對其降解起作用的細菌提供了碳源,使這類細菌大量繁殖,因此堆肥C和D的細菌多樣性顯著高于堆肥B。如表2所示,4個堆肥樣品中的真菌多樣性變化不顯著,這可能是因為在(58±2)℃的特殊環境下,僅部分嗜熱真菌能夠生存。而在堆肥A中,雖然是室溫環境,但堆肥濕度僅約18%,干燥的環境不適宜多數真菌生存,因此4個堆肥樣品的生物多樣性沒有顯著改變。Ace和Chao指數用來估計群落中含有OTU數目,兩者算法不同,都體現了堆肥樣品的微生物群落豐度。堆肥樣品中細菌豐度的變化規律與多樣性類似。由于工業堆肥(58±2)℃的特殊環境,造成真菌多樣性的變化不大,而試驗結果進一步顯示真菌豐度受堆肥中溫度和碳源的影響,其中堆肥B(對照組)顯示出了最低的真菌豐度。
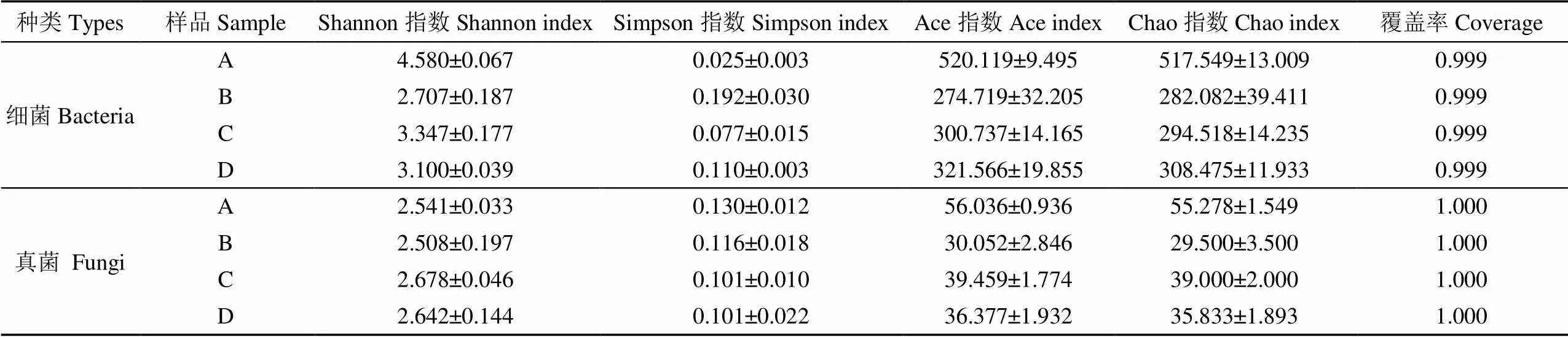
表2 堆肥中細菌/真菌Alpha多樣性指數
2.3 堆肥中微生物的物種組成
進一步對堆肥樣品的物種組成相似性及重疊情況進行研究,可以分析得到4組堆肥樣品中所共有和獨有的物種數目(圖2)。在細菌屬水平上,與堆肥A相比,堆肥B中由于溫度升高,碳源不足,241種細菌消失,并出現6種新的細菌。而由于碳源的添加,原本在堆肥B中消失的細菌,又在堆肥C和D中分別出現了46和38種,其中19種是共有細菌。4組堆肥中真菌的物種數目整體偏低,這與上述Shannon和Simpson指數所體現出的真菌多樣性結果一致。堆肥D中顯示出了3種獨有真菌。
圖3展示了屬水平下堆肥中細菌和真菌的組成。在屬水平上,4種堆肥里排名前十的菌群有,SBR1031,n_S0134,,,,,,和。相較于室溫存放的原堆肥,工業堆肥條件下的堆肥B,C,D樣品中,在屬水平上出現了2種獨有菌群,分別是(各占1.63%,5.47%和4.82%)和(各占7.21%,0.70%和2.69%)。此外,和的占比顯著提高。在堆肥B,C,D中,最主要的優勢細菌是,其占比從原堆肥A中的0.52%分別提高到40.69%,20.25%和39.44%。(球桿菌屬)屬于(球桿菌科),(放線菌綱)[20]。Ma等[21]研究了污泥堆肥過程中微生物演替的變化,研究發現在整個堆肥時間演變的過程中,都是優勢菌群之一。Storey等[22]分別將用乳品廢水污泥和硝酸銨鈣作為氮源加入用來生產堆肥。研究結果發現,在兩種堆肥中,都是堆肥成熟階段最豐富的菌屬,此結果與本研究一致。除以外,優勢菌群中的,和均屬于放線菌[20]。放線菌在堆肥過程中非常重要,因為它們能夠降解纖維素和幾丁質,大多數報道的-葡萄糖苷酶序列與放線菌門有關,這是堆肥成熟度的典型指標[23-24]。而優勢菌群中的其他菌屬所屬的門,如,,等,都是堆肥中常見的細菌類別[25-26]。
相較于堆肥A,在真菌屬水平上堆肥B,C,D中(ABCD分別為28.87%,5.43%,5.68%,8.82%)和(ABCD分別為11.08%,2.18%,4.30%,2.06%)的豐度顯著降低,而(ABCD樣品中分別為1.13%,8.24%,20.96%,18.29%),(ABCD樣品中分別為1.61%,21.89%,9.34%,10.12%),(ABCD樣品中分別為0.14,10.76%,8.98%,12.42%)和(ABCD樣品中分別為0.21%,6.43%,5.31%,4.64%)的豐度顯著增高。說明這些真菌屬均受工業堆肥過程影響。
2.4 堆肥樣本物種差異分析
堆肥B作為工業堆肥環境下的對照組樣品,其溫濕度等環境條件與堆肥C,D保持一致,僅缺少額外添加的碳源(微晶纖維素/生物降解購物袋)。因此我們使用Student’s T檢驗,分別比較堆肥B,C和堆肥B,D中占比排名前15位的細菌及真菌物種組成差異,堆肥C、D中顯著增加的微生物可被認為是參與工業堆肥降解作用的關鍵菌。結果表明,屬水平下堆肥C中沒有顯著增多的真菌,而顯著增多的細菌有0134(=0.011 7),(=0.007 1)和(=0.0158)。0134屬于(門),僅與土壤環境相關,被報道不利于固氮和固碳[27-28]。屬于(腈基降解菌亞綱),與同屬(放線菌綱)。
相較于堆肥B,堆肥D中的屬細菌(=0.043 4)和屬真菌(=0.0469)的含量顯著增加。屬下的一些菌株,被報道具有酯酶和脂肪酶活性,如從植物根際土壤中分離出的的酯酶和脂肪酶活性被報道呈陽性[29],的酯酶(C4)活性呈陽性[30],的酯酶(C4)、酯酶/脂肪酶(C8)也被報道呈陽性[31]。PBAT和PLA均是通過酯鍵縮合而成,其降解過程需要通過酯鍵的斷裂來完成。此外,嗜熱真菌被認為是堆肥材料中的重要微生物群,因為它們與頑固底物(如纖維素、半纖維素和木質素等)的降解有關,其中是堆肥中常見的一種[21,32]。
進一步對堆肥C和D中的細菌在屬水平上進行比較。堆肥C中的0134,和的占比相較于堆肥D顯著更高。結合上述分析可知,和(屬放線菌綱)對微晶纖維素的降解具有特定積極作用。在真菌屬水平上,堆肥D中的相較于堆肥C具有更高占比,然而對比堆肥B,在堆肥C、D中的物種占比都呈下降趨勢,說明該真菌屬在添加外源纖維素/生物降解塑料的工業堆肥過程中受到了抑制。其中微晶纖維素在降解過程中產生的中間產物,對有更強的抑制作用。在自然界中,真菌被認為是纖維素的主要降解微生物。雖然放線菌對纖維素的降解能力不及真菌,但由于工業堆肥(58±2)℃的特殊高溫環境不適宜多數真菌生存,而放線菌能夠形成孢子,較真菌耐高溫。因此在高溫條件下,放線菌對纖維素和PBAT-PLA膜袋的降解都起了重要作用[25,33]。
綜合上述分析我們可以發現,堆肥C,D中大量存在的優勢菌主要是放線菌,且較對照組B具有顯著增長優勢。對比堆肥C、D,和對微晶纖維素的降解具有特定積極作用。由于工業堆肥(58±2)℃的特殊高溫環境,PBAT-PLA膜袋與微晶纖維素的關鍵降解微生物相似,均屬放線菌綱,在屬水平上略有差異。放線菌對聚酯物的解聚以及纖維素的降解具有積極的作用,但有研究報道,它們不能代謝一些所形成的產物[6,34-35]。因此,對試驗結果進行分析可表明完整的聚酯和纖維素的生物降解是多種微生物協同作用的結果。
2.5 堆肥樣本比較分析
主坐標分析(Principal Co-ordinates Analysis,PCoA)(圖4)直觀地展示了屬水平下堆肥樣本之間細菌/真菌的差異性。在細菌PCoA圖中,堆肥A和堆肥B處在不同的象限,細菌群落差異很大。堆肥C和D由于添加了碳源(微晶纖維素/生物降解膜袋),因此與堆肥B也有差異,但堆肥C和堆肥D的細菌群落差異較小。4種堆肥在屬水平下的真菌群落差異也顯示了類似的規律。結果與上文的指標保持了較好的一致性。
2.6 掃描電鏡圖像
研究認為微生物的分解主要發生在有機物表面的生物膜中[6]。因此利用掃描電鏡分別觀察微晶纖維素及生物降解膜袋在工業堆肥前后的表面微觀形態,從而對其工業堆肥降解機制進行一定的研究(圖5)。掃描電鏡結果顯示在微晶纖維素和生物降解膜袋表面都覆蓋了一層生物膜,且構成這一類生物膜的主要是成簇團狀的橢圓形菌。經過與16 s堆肥中細菌組成結果的比對確認,我們可以推斷這些橢圓的菌為(球桿菌)。在膜袋降解所形成的生物膜中,還有少部分條狀菌(推測其為放線菌)和絲狀物(推測其為絲狀真菌)。因此推測PBAT-PLA生物降解膜袋的降解,是微生物(包括真菌,細菌,如放線菌等)群落定植并產生的協同作用效應[36]。
2.7 討論
本研究根據標準GB/T 19277.1—2011,在受控工業需氧堆肥條件下,對微晶纖維素和PBAT-PLA生物降解膜袋降解過程中的微生物響應機制進行研究。第150天時,PBAT-PLA膜袋和微晶纖維素的生物分解率分別為98.8%±1.8%和95.1%±2.2%。故選取第140天的各組堆肥樣品進行16S rDNA和18S rDNA高通量測序,此時的降解過程已處于中后期,微生物群落已相對穩定,并且由于碳源未耗盡,微生物依然保持活性。本研究旨在通過對降解過程中堆肥的細菌/真菌的群落變化分析,分析堆肥中的優勢菌、顯著增長菌,從而為今后篩選降解關鍵作用菌提供重要理論依據,后續有望在堆肥降解過程中,接種該菌,提高該菌濃度,從而加快降解速度,幫助突破現有可降解檢測技術中檢測周期長的技術壁壘,提高檢測效率。同時,通過對PBAT-PLA膜袋(為可降解農用地膜最常用配方)降解所產堆肥的微生物群落進行分析,能夠作為重要參考指標,用以評估生物降解塑料所生產的堆肥對農作物生長、農業生態環境等的影響。因此,本研究為工業堆肥技術的優化和可降解塑料檢測標準的完善奠定了理論基礎。同時,為可降解塑料膜袋的規模化推廣,及其堆肥產品在新型功能農業領域應用的可行性研究提供科學依據。
3 結 論
1)在工業堆肥(58±2)℃的條件下,對照組堆肥相較于原堆肥,細菌多樣性顯著降低,僅部分嗜熱菌繼續生存。而纖維素和生物降解膜袋堆肥,由于提供了額外碳源,細菌多樣性顯著高于堆肥B。4個堆肥樣品中細菌豐度的變化規律與多樣性類似。真菌多樣性因受工業堆肥(58±2)℃這一特殊環境影響并無顯著性變化,而豐度也受堆肥中溫度和碳源的影響,其中對照組堆肥顯示出了最低的真菌豐度。
2)在屬水平上,對照組、纖維素和生物降解膜袋堆肥中最主要的優勢細菌是,其占比從堆肥A中的0.52%分別提高到40.69%,20.25%和39.44%。除以外,優勢菌群中的,和均屬于放線菌。
3)使用Student’s T檢驗,分別比較對照組和纖維素堆肥,以及對照組和生物降解膜袋堆肥的細菌及真菌物種組成差異。屬水平下纖維素堆肥中顯著增多的細菌有0134,和,其中后兩者屬(放線菌綱)。相較于對照組堆肥,生物降解膜袋堆肥在屬水平上顯著增加的細菌是,顯著增加的真菌是。進一步比較纖維素和生物降解膜袋堆肥,發現和(屬放線菌綱)對微晶纖維素的降解具有特定積極作用。在工業堆肥(58±2)℃的特殊高溫環境下,參與PBAT-PLA膜袋與微晶纖維素降解過程的關鍵降解微生物相似,均屬于放線菌綱,僅在屬水平上略有差異。
4)通過掃描電鏡觀察微晶纖維素/生物降解膜袋的微觀形貌,發現其表面覆蓋了一層生物膜,其主要構成為簇團狀的橢圓形菌,推斷其為。
綜合上述分析可以發現,與對照組堆肥相比,纖維素和生物降解膜袋堆肥中的大量存在并顯著增長的優勢菌屬為,屬于放線菌。放線菌對聚酯物的解聚以及纖維素的降解具有積極的作用,但完整的的聚酯和纖維素的生物降解不依賴單一菌種,而是微生物協同作用的結果。
[1] 聯合國向“白色污染”宣戰[N]. 2021-12-30,http: //www. jjckb. cn/2018-12/10/c_137662157. htm?from=singlemessage.
[2] 刁曉倩,翁云宣,宋鑫宇,等. 國內外生物降解塑料產業發展現狀[J]. 中國塑料,2020,34(5):123-135.
Diao Xiaoqian, Weng Yunxuan, Song Xinyu, et al. Development status of biodegradable plastics industry at home and abroad[J]. China Plastics, 2020, 34(5): 123-135. (in Chinese with English abstract)
[3] Shah A A, Hasan F, Hameed A, et al. Biological degradation of plastics: A comprehensive review[J]. Biotechnology Advances, 2008, 26(3): 246-265.
[4] Tosin M, Degli-Innocenti F, Bastioli C. Detection of a toxic product released by a polyurethane-containing film using a composting test method based on a mineral bed[J]. Journal of Environmental Polymer Degradation, 1998, 6(2): 79-90.
[5] Sudesh K, Iwata T. Sustainability of biobased and biodegradable plastics[J]. Clean–Soil, Air, Water, 2008, 36(56): 433-442.
[6] Lucas N, Bienaime C, Belloy C, et al. Polymer biodegradation: Mechanisms and estimation techniques: A review[J]. Chemosphere, 2008, 73(4): 429-442.
[7] Haider T P, V?lker C, Kramm J, et al. Plastics of the future? The impact of biodegradable polymers on the environment and on society[J]. Angewandte Chemie International Edition, 2019, 58(1): 50-62.
[8] Gu J D. Microbiological deterioration and degradation of synthetic polymeric materials: Recent research advances[J]. International Biodeterioration and Biodegradation, 2003, 52(2): 69-91.
[9] Weng Y X, Jin Y J, Meng Q Y, et al. Biodegradation behavior of poly (butylene adipate-co-terephthalate)(PBAT), poly (lactic acid)(PLA), and their blend under soil conditions[J]. Polymer Testing, 2013, 32(5): 918-926.
[10] Bonilla J, Paiano R B, Louren?o R V, et al. Biodegradability in aquatic system of thin materials based on chitosan, PBAT and HDPE polymers: Respirometric and physical-chemical analysis[J]. International Journal of Biological Macromolecules, 2020, 164: 1399-1412.
[11] Zhang M, Jia H, Weng Y, et al. Biodegradable PLA/PBAT mulch on microbial community structure in different soils[J]. International Biodeterioration and Biodegradation, 2019, 145: 104817.
[12] 中國國家標準化管理委員會. GBT 19277. 1. 受控堆肥條件下材料最終需氧生物分解能力的測定采用測定釋放的二氧化碳的方法第1部分:通用方法[S]. 北京:中國標準出版社,2011.
[13] Huws S A, Edwards J E, Kim E J, et al. Specificity and sensitivity of eubacterial primers utilized for molecular profiling of bacteria within complex microbial ecosystems[J]. Journal of Microbiological Methods, 2007, 70(3): 565-569.
[14] Rousk J, B??th E, Brookes P C, et al. Soil bacterial and fungal communities across a pH gradient in an arable soil[J]. The ISME Journal, 2010, 4(10): 1340-1351.
[15] Ren S, Lu A, Guo X, et al. Effects of co-composting of lincomycin mycelia dregs with furfural slag on lincomycin degradation, degradation products, antibiotic resistance genes and bacterial community[J]. Bioresource Technology, 2019, 272: 83-91.
[16] Edgar R C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads[J]. Nature Methods, 2013, 10(10): 996.
[17] Chen X, Hou F, Wu Y, et al. Bacterial and fungal community structures in Loess Plateau grasslands with different grazing intensities[J]. Frontiers in Microbiology, 2017, 8: 606.
[18] Oberlintner A, Baji? M, Kal?íková G, et al. Biodegradability study of active chitosan biopolymer films enriched with Quercus polyphenol extract in different soil types[J]. Environmental Technology & Innovation, 2021, 21: 101318.
[19] Huang Y, Pan H, Wang Q, et al. Enrichment of the soil microbial community in the bioremediation of a petroleum-contaminated soil amended with rice straw or sawdust[J]. Chemosphere, 2019, 224: 265-271.
[20] Holt J G. Bergey's Manual of Determinative Bacteriology[M]. Philadelphia, Lippincott Williams and Wilkins, 1993.
[21] Ma C, Hu B, Liu F U Y, et al. Changes in the microbial succession during sewage sludge composting and its correlation with physico-chemical properties[J]. Nature Environment and Pollution Technology, 2020, 19(1): 237-244.
[22] Storey S, Chualain D N, Doyle O, et al. Comparison of bacterial succession in green waste composts amended with inorganic fertiliser and wastewater treatment plant sludge[J]. Bioresource Technology, 2015, 179: 71-77.
[23] Steger K, Sj?gren ? M, Jarvis ?, et al. Development of compost maturity andpopulations during full-scale composting of organic household waste[J]. Journal of Applied Microbiology, 2007, 103(2): 487-498.
[24] Jimenez L, Kulko E, Veloz E, et al. 16S rRNA identification of microorganisms and direct detection of functional genes in waste material generated by an in-vessel rotating compost system[J]. EC Microbiology, 2015, 1(3): 129-142.
[25] Tian W, Sun Q, Xu D, et al. Succession of bacterial communities during composting process as detected by 16S rRNA clone libraries analysis[J]. International Biodeterioration and Biodegradation, 2013, 78: 58-66.
[26] Li J, Chen Y T, Xia Z Y, et al. Changes in bacterial communities during a pilot-scale composting process of dairy manure[J]. Journal of Environmental Engineering, 2020, 146(9): 04020095.
[27] Zhang W, Yu C, Wang X, et al. Retracted: Increased abundance of nitrogen fixing bacteria by higher C/N ratio reduces the total losses of N and C in cattle manure and corn stover mix composting[J]. Waste Management, 2020, 4: 416-425.
[28] Mujaki? I, Andrei A-?, Shabarova T, et al. Common presence of phototrophic gemmatimonadota in temperate freshwater lakes[J]. Msystems, 2021, 6(2): e01241-20.
[29] Yuan L J, Zhang Y Q, Yu L Y, et al.., a novel actinomycete isolated from rhizosphere soil of the plant[J]. International Journal of Systematic and Evolutionary Microbiology, 2010, 60(1): 51-54.
[30] Cao Y R, Jiang Y, Wu J Y, et al. Actinopolymorpha alba sp. nov., isolated from a rhizosphere soil[J]. International Journal of Systematic and Evolutionary Microbiology, 2009, 59(9): 2200-2203.
[31] Wang Y X, Zhang Y Q, Xu L H, et al. Actinopolymorpha rutila sp. nov., isolated from a forest soil[J]. International Journal of Systematic and Evolutionary Microbiology, 2008, 58(10): 2443-2446.
[32] Langarica-Fuentes A, Handley P S, Houlden A, et al. An investigation of the biodiversity of thermophilic and thermotolerant fungal species in composts using culture-based and molecular techniques[J]. Fungal Ecology, 2014, 11: 132-144.
[33] Godden B, Ball A S, Helvenstein P, et al. Towards elucidation of the lignin degradation pathway in actinomycetes[J]. Microbiology, 1992, 138(11): 2441-2448.
[34] Kleeberg I, Hetz C, Kroppenstedt R M, et al. Biodegradation of aliphatic-aromatic copolyesters by Thermomonospora fusca and other thermophilic compost isolates[J]. Applied and Environmental Microbiology, 1998, 64(5): 1731-1735.
[35] Witt U, Einig T, Yamamoto M, et al. Biodegradation of aliphatic–aromatic copolyesters: Evaluation of the final biodegradability and ecotoxicological impact of degradation intermediates[J]. Chemosphere, 2001, 44(2): 289-299.
[36] Sang B I, Hori K, Tanji Y, et al. Fungal contribution to in situ biodegradation of poly (3-hydroxybutyrate-co-3-hydroxyvalerate) film in soil[J]. Applied Microbiology and Biotechnology, 2002, 58(2): 241-247.
Microorganism responded biodegradation mechanism of PBAT-PLA biodegradable packaging under industrial aerobic composting
Ma Yichao1,2, Liu Jun1※, Zhang Ruohong1, Zhu Jiahuan1, Du Huanzheng2, Lu Sha2
(1.,201114,;2.,200086,)
Plastic pollution has posed a serious threat to the environment in the world. Biodegradable plastics can be widely expected to effectively mitigate plastic pollution. The subsequent industrial composting can also be treated to reduce the growing demand for landfills. In this research, the microorganisms responded to the biodegradation mechanism was proposed for the PBAT-PLA biodegradable packaging under the controlled industrial aerobic composting, according to the national standard (GB/T 19277.1-2011). Microcrystalline cellulose was used as reference material. The compost samples were taken on day 140 when the degradation was in the active period. The initial compost (before degradation), compost without any biodegradable material, compost with microcrystalline cellulose, and compost with PBAT-PLA packaging materials were then labeled as compost A, B, C, and D, respectively. The bacterial/fungal community was analyzed during degradation, including the species diversity, speciation, and species difference. A sample comparison was made through 16S/18S high-throughput sequencing of microorganisms in compost. The microscopic morphologies of PBAT-PLA packaging materials and microcrystalline cellulose were characterized by a Scanning Electron Microscope (SEM), in order to explore the intrinsic microbial response to the degradation mechanism. The results showed that the bacterial diversity of compost B under the industrial composting at 58 ± 2 ℃ was significantly lower than that of initial compost A since only some thermophilic bacteria survived. The bacterial diversities in compost C and D were significantly higher than that of compost B, due to the addition of cellulose or biodegradable packaging materials which provided carbon sources. There was no significant change in the fungal diversity, due to unsuitable for most fungi to survive at high temperatures. The fungal abundance changed, due to the temperature and carbon source in the compost, where the compost B (blank sample) showed the lowest fungal abundance. The dominant bacteria in the compost C and D wereandwhichbelonged to. The student’s T-test was used to compare the composition of bacteria and fungi in the compost C and D with those in the compost B. The significant growing bacteria in the compost C and D, includingand, belonged to. Therefore, there were similar degrading microorganisms of PBAT-PLA membrane bag and microcrystalline cellulose at (58±2) ℃. The SEM images showed that the surface of PBAT-PLA packaging after degradation was covered with a layer of biofilm. The biofilm was mainly composed of the clusters of ellipsoid bacteria, which were inferred to be.presented a positive effect on the depolymerization of polyester and the degradation of cellulose under industrial composting conditions. The complete biodegradation of polyester and cellulose depended on the microbial synergy, rather than a single strain. Therefore, the dominant bacteria and significant growth bacteria of biodegradable materials under industrial composting conditions can be expected to serve as a theoretical basis for screening key biodegradation bacteria in the future. It can also greatly contribute to breaking through the technical barriers of the long detection cycle with the current biodegradation for higher efficiency and speed. The identification of microbial communities can be an important indicator to evaluate the impact of the compost produced by biodegradable plastics on crops growth, and agricultural ecological environment. Consequently, a theoretical foundation can be made to optimize the industrial composting and biodegradable plastic testing standards. The finding can also provide a scientific basis for the large-scale promotion of biodegradable plastic films and the application of the compost produced from biodegradable plastics in advanced (or modern) agriculture
microorganism, plastic, degradation, biodegradable packaging, industrial aerobic composting, high throughput sequencing
2021-09-15
2021-11-30
國家自然科學基金(71974144);國家社科基金重大項目(21ZDA087);上海市質量監督檢驗技術研究院科研項目(KY-2021-2-QH)
馬昳超,博士,研究方向為可降解塑料、生物系統工程、生物質利用。Email:mayc@sqi.org.cn
劉峻,博士,教授級高級工程師,研究方向為食品相關產品安全風險評估。Email:liujun@sqi.org.cn
10.11975/j.issn.1002-6819.2021.24.025
X7
A
1002-6819(2021)-24-0224-08
馬昳超,劉峻,章若紅,等. 微生物響應PBAT-PLA生物降解膜袋工業需氧堆肥降解機制[J]. 農業工程學報,2021,37(24):224-231. doi:10.11975/j.issn.1002-6819.2021.24.025 http://www.tcsae.org
Ma Yichao, Liu Jun, Zhang Ruohong, et al. Microorganism responded biodegradation mechanism of PBAT-PLA biodegradable packaging under industrial aerobic composting[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2021, 37(24): 224-231. (in Chinese with English abstract) doi:10.11975/j.issn.1002-6819.2021.24.025 http://www.tcsae.org
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
小學科學(學生版)(2018年3期)2018-04-18 12:34:19
知識經濟·中國直銷(2017年10期)2017-11-07 02:39:52
知識經濟·中國直銷(2017年3期)2017-04-16 03:08:01
文學少年(有聲彩繪)(2017年2期)2017-03-06 08:10:54
