中日沿海部分魚類DNA條形碼研究
2021-03-18 01:53:22黃志基張玉寧
海洋漁業 2021年1期
關鍵詞:物種
羅 純,章 群,黃志基,張玉寧
(暨南大學生態學系,廣州 510632)
中國與日本海域相連,是南北暖冷魚類的重要交匯區。一方面,由于同源擴散的結果,兩國沿海部分魚類特別是近海種類呈現高度相似性;另一方面,由于地理環境有所區別,再經過歷史演替,兩國沿海魚類在共性的基礎上也呈現一定差異[1]。近年來,隨著中國沿海經濟不斷發展,海洋漁業資源被過度利用,漁獲物低齡化現象嚴重[2],對海洋魚類資源開展深入研究有助于漁業的長期、可持續性發展。
為了有效保護和管理魚類資源,對魚類進行鑒定分類必不可少。由于傳統分類基于外部形態特征,對分類者的專業知識和經驗要求較高,同時有些物種形態結構易受到地理環境和不同發育階段的影響;還有部分魚類存在雌雄轉換現象,具有性別二態性[3]:如隆頭魚科魚類為世界性分布,多數種存在幼魚期、成魚期及超雄期之間以及雌雄之間的體色和形態的差異[4],給物種分類鑒定造成了一定困難。
DNA條形碼不僅可以識別整條魚,還可以識別魚片、魚鰭和其他難以根據形態學識別的魚體碎片[5]。2005年WARD等[6]利用COI基因對澳大利亞207種魚類進行鑒定,表明COI基因能夠有效區分不同物種。動物線粒體COI基因5′端約650 bp序列已被公認為是有效區分不同物種的DNA條形碼標準區域[7-8]。大量研究也表明,線粒體DNA條形碼技術在解決同種異名、異種同名、發掘隱藏種方面較傳統形態分類具有顯著優勢。
目前,對中日沿海魚類的研究報道多為生態資源調查[1]、遺傳結構與分子系統地理學研究[9-11]等。在分類方面,日本學者河野光久等[12]對日本海西南部山口縣的魚類進行了統計學研究,共確認了39目197科870種。荒見真一郎等[13]運用DNA條形碼技術鑒定了神奈川縣市場31種魚類。李昂等[14]對中國近海和日本近海的白姑魚分類地位和系統進化進行了研究。本研究測定了在日本沿海采集的37種魚類的184條COI基因序列,并結合GenBank下載的序列,共分析了中日沿海共有種327條COI基因序列,以期確定日本及中國沿海魚類的分類地位,為相關海域魚類種質資源保護和可持續利用提供科學指導。
1 材料與方法
1.1 樣品來源及分子實驗
研究樣品于2018年4月—2019年7月采集于日本太平洋側瀨戶內海明石沖(34°38′41″N、134°59′23″E)和日本海側若狹灣舞鶴(35°27′01″N、135°18′52″E)等地,并置于95% 乙醇中保存。依據Fishbase數據庫,并參照《中國魚類系統檢索》[15]、《黃渤海魚類圖志》[16]等資料進行形態鑒定,測序標本和下載序列信息見表1。
取0.2 g魚尾鰭肌肉晾干后,參考樂小亮[17]改進的苯酚/氯仿法提取DNA。參考WARD[18]方法進行PCR 擴增。使用引物FishF1:5′-TCAACCAACCACAAAGACATTGGCAC-3′,FishR1:5′-TAGACTTCTGGGTGGCCAAAGAATCA-3′,反應體系為20μL:上下游引物各1μL,PCR Mix 10 μL,DNA模板1μL,ddH2O 7μL。PCR反應條件為:95℃預變性5 min,95℃變性40 s,55℃退火40 s,72℃延伸50 s,共35個循環,最后72℃延伸10 min。4℃保存。擴增產物經1% 瓊脂糖凝膠電泳檢測后送至北京六合華大基因公司正向測序。
1.2 數據處理
通過Bioedit 7.0.9.0[19]軟件上對測序峰圖進行人工校對。運用MEGA 7.0[20]軟件分析序列特征,計算堿基組成、變異位點、簡約信息位點、單一可變位點等分子多樣性指數;基于Kimura 2-parameter(K2P)替代模型,分別計算種內、屬內種間、屬內、科內屬間遺傳距離,經1 000次重復抽樣(bootstraps)檢測分支置信度構建鄰接(neighbor-joining)樹。采用自動條形碼間隙檢索方法 (automatic barcode gap discovery,ABGD)[21]將全部序列自動分組進行假設種分析,同時利用種內遺傳距離及種間遺傳距離繪制條形碼間隙圖。
2 結果與分析
2.1 COI基因序列特征
參照國際上通用的DNA條形碼長度,截取平均長度為638 bp(552 bp ~652 bp)的5′端COI序列進行分析[22]。在本研究測定序列和GenBank下載的7目24科35屬40種327條序列中,沒有發現堿基插入和缺失現象。共檢測到315個變異位點,其中簡約信息位點有308個,單一可變位點有7個。堿基T、C、A、G的平均含量分別為29.4%、28.5%、23.2%、18.8%,A+T含量(52.7%)高于G+C含量(47.3%),表現出明顯的堿基組成偏向性,該結果與動物COI基因堿基組成中普遍存在的AT含量高于GC含量的特征一致[18]。
2.2 各階元平均遺傳距離概況
各分類階元的K2P遺傳距離如表2和圖1所示。40種魚類種內遺傳距離為0~3.91%,平均遺傳距離為0.42%;屬內種間遺傳距離為0.32%~17.64%,平均遺傳距離為6.25%;屬內種間平均遺傳距離是種內平均遺傳距離的14.88倍,符合HEBERT[22]提出的物種鑒別“十倍法則”,即種間遺傳距離遠大于種內遺傳距離的10倍以上。圖1顯示:種內遺傳距離98%集中在0~2%區間;屬內種間遺傳距離集中在2%~18%區間內,二者形成了明顯的條形碼間隙。
2.3 條形碼分子系統樹分析
從圖2鄰接樹看,327條序列形成了41分支,平均遺傳距離分支間21.81%(1.97% ~28.29%)是分支內0.30%(0~0.81%)的73倍(圖3),40種魚類有36種形成了單系分支,支持其物種有效性。白姑魚(Pennahia argentata)形成2個分支,中國和日本全部個體各自聚類成一支,分支間遺傳距離為3.28%。花鱸(Lateolabraxspp.)形成2個自展支持率為100%的分支,日本和中國個體分別成支,分支間遺傳距離為6.98%。赫氏高眼鰈(Cleisthenes herzensteini)和松木高眼鰈(C.pinetorum)混雜分布在同一分支上,分支內的遺傳距離為0.23%。在ABGD分析中(圖4),當種內先驗遺傳距離P為0.021 5% 時,40種魚類被劃分為40個假設種。瓦氏(Callionymus valenciennei)和飾鰭斜棘(Repomucenus ornatipinnis)被歸為一個假設種,其余種的劃分與鄰接樹形成的分支相一致。

·續表1·
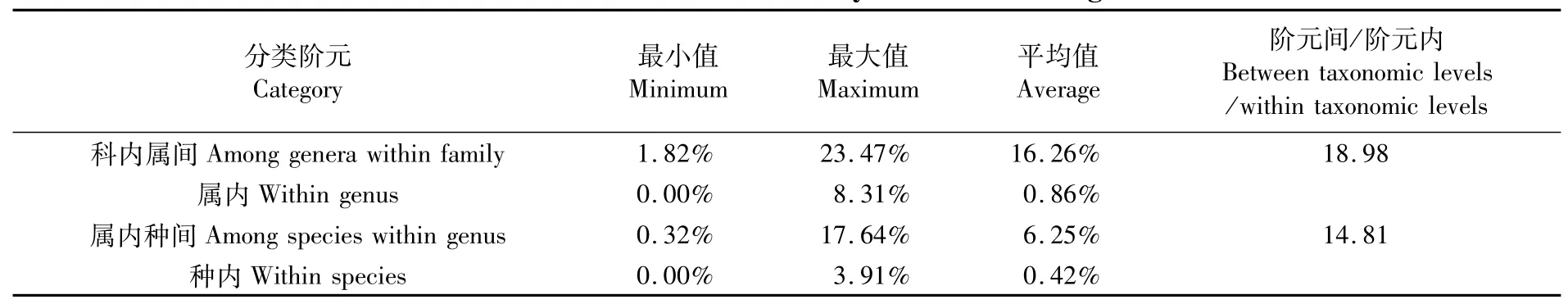
表2 不同分類階元的遺傳距離Tab.2 Genetic distance summary of different categories

圖1 不同分類階元的遺傳距離圖Fig.1 Distribution chart of genetic distance for 40 species based on the K 2P model
本研究采取同一物種內多個個體比較分析的策略,計算每一種魚類種內不同樣本間的遺傳距離,結合各種間遺傳距離繪制40種魚類條形碼間隙圖,以獲得種內與種間遺傳距離的差異度。圖5顯示松木高眼鰈(9)和赫氏高眼鰈(10)最大種內遺傳距離未能與最小種間遺傳距離很好的分離,其余98%的魚類均能夠形成良好的條形碼間隙。
3 討論
3.1 花鱸與白姑魚分類地位的探討
YOKOGAWAL等[23]研究發現,日本花鱸(L.japonicus)和中國花鱸(L.maculatus)在形態和遺傳上具有顯著差異,前者側線鱗片數76 ~92,鰓耙數24 ~30,后者側線鱗片數66 ~82,鰓耙數19 ~24;且基于同工酶等位基因頻率計算的遺傳距離為0.174,應為2個不同的物種。中山耕至[24]研究發現,中國花鱸和日本花鱸細胞色素類型與分布不同。更新世冰期海平面下降,日本海和中國東海成為2個獨立的避難所,使分布其中的鱸魚逐漸分化[25]。在本研究中,花鱸種內平均遺傳距離為3.91%,大于2%的物種遺傳閾值;在鄰接樹上形成中國群體和日本群體2個分支,分別對應于中國花鱸和日本花鱸,分支間平均遺傳距離(6.98%)是分支內平均遺傳距離(0.49%)的14.2倍,符合“10倍法則”;支持將二者作為2個物種的分類處理。
在鄰接樹上,中國和日本白姑魚各自獨立成支,平均遺傳距離分支間(3.28%)是分支內(0.29%)的11.3倍;ABGD也將白姑魚分為2組。目前,國內外學者已從生物學特性等方面對其進行了研究,韓志強等[26-27]通過白姑魚線粒體DNA控制區和Cytb基因序列和AFLP技術對白姑魚系統地理分布模式的研究,表明中日白姑魚組群遺傳分化顯著;李昂[28]通過對中日白姑魚的耳石形態和線粒體基因組全序列分析發現,二者存在一定程度的分化,認為邊緣海的隔離是造成白姑魚分化的原因,海洋洋流、海水溫度以及東海深海溝是維持當前分化的主要因素[27]。本研究中,根據物種鑒定的種間遺傳距離是種內遺傳距離的10倍和種內遺傳距離一般小于2%法則,推測中國沿海和日本的白姑魚是兩個亞種乃至不同種。

圖2 基于線粒體COI基因序列的40種魚類鄰接樹Fig.2 Neighbor-joining tree based on m tDNA COI sequences of 40 fish species

圖3 41個分支的分支內(左)和分支間(右)遺傳距離Fig.3 Genetic distance w ithin(left)and among(right)41 clades

圖4 中日沿海部分魚類的ABGD分析圖Fig.4 Automatic barcode gap discovery analysis of some coastal fishes in China and Japan
3.2 高眼鰈屬分類地位的探討
赫氏高眼鰈和松木高眼鰈混雜在遺傳距離為0.23%的分支內,種間平均遺傳距離(0.32%)和種內遺傳距離(0.23%),均在一般物種的種內遺傳范圍內。由于沒有赫氏高眼鰈的憑證標本,推測可能存在鑒定錯誤的情況,但不排除存在譜系揀選尚未完成的可能性。
3.3 瓦氏與飾鰭斜棘分類地位的探討

圖5 40種魚類DNA條形碼間隙圖Fig.5 DNA barcoding gap for 40 fish species
3.4 花鰭海豬魚的分類探討
本研究中,花鰭海豬魚形態差異較大,其種內平均遺傳距離為0.56%,且能形成單系分支。花鰭海豬魚幼體表現為雌性,成體后轉換為雄性,且雌雄個體之間存在體色和形態的差異[33-34],易被誤認為不同的物種。福井行雄等[35]研究發現,雌性個體體色偏黃,體側中央有一細的明顯褐色縱帶,自吻端經眼徑直達尾鰭基,此縱帶在胸鰭上方及尾鰭基處黑色加深;而雄性個體在胸鰭基底后方上具一大黑斑,且背鰭、臀鰭和尾鰭有紅色斑紋。而本研究中花鰭海豬魚的形態差異就是雌雄個體的差異。事實上,大部分隆頭科魚類存在性別二態性,了解雌雄之間的差異對于種類鑒定、形態研究以及判斷雌、雄在繁殖過程中的不同角色有著重要的意義。
綜上所述,基于COI基因的DNA條形碼技術能夠對日本大部分海洋魚類進行有效的區分,說明線粒體COI基因作為日本海洋魚類DNA條形碼適用性廣,對于目前研究尚不深入的非經濟魚類,尤其是易引起誤鑒的雌雄異形的魚類,展開線粒體COI基因的研究工作尤為重要。本研究中,由于西北太平洋獨特的地理構造和復雜的洋流體系,分布于中日沿海的魚類呈現多樣的分布格局。對于中日海洋魚類來說,絕大多數種類并沒有顯著的遺傳分化,這可能與黑潮和親潮的輸送作用有關。而中日白姑魚達到亞種乃至2個不同種的水平、日本花鱸和中國花鱸間的遺傳分化,應是與具體類群的獨特生理生態習性和邊緣海的隔離有關。由于可參考的魚類檢索工具書的欠缺,樣品的地理分布廣泛,難以獲得不同區域樣品進行近緣種的比較,導致存在物種分類鑒定錯誤的可能,如松木高眼鰈和赫氏高眼鰈可能因錯誤鑒定而出現聚在同一分支上的情況。另外,母系遺傳的線粒體COI基因不能反映雙親遺傳的核基因信息,難以區分物種的雜交滲透狀況。因此,準確的物種鑒定不僅需要DNA條形碼進行輔助研究,在今后的工作中仍需補充其同屬其他物種,同時結合形態測量、核基因標記以及生物學研究,以進一步明確日本與中國海洋魚類的分類地位。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
