氮營養鹽對極北海帶幼苗生長和生理生化特性的影響
2021-03-25 01:47:16袁艷敏劉福利梁洲瑞
水產科學 2021年2期
袁艷敏,劉福利,2,梁洲瑞,2,王 瀟,3
(1.中國水產科學研究院 黃海水產研究所,農業農村部海洋漁業資源可持續利用重點開放試驗室,青島 266071;2.青島海洋科學與技術國家試驗室,海洋漁業科學與食物產出過程功能試驗室,青島 266000;3.上海海洋大學 水產與生命學院,上海 201306)
近年來,海水富營養化和重金屬污染問題日益嚴重,對海水養殖業和人類健康造成極大威脅。海藻是海洋生態系統的初級生產者,利用海藻的吸收、富集和降解作用可以去除污水中的營養物質、重金屬和有機毒物,具有顯著的生態環境改善效果[1]。極北海帶(Laminariahyperborea)耐低光能力極強,其光補償點可低至4.6 μmol/(m2·s)[2],適于在較深的水域生長。當前我國正在積極推進海洋牧場建設,海藻場是決定海洋牧場效益的關鍵組成因素。由于我國近海海水透明度普遍較低,投放的人工魚礁水層較深,因光照限制很難增殖起規模化的“海藻森林”,而極北海帶擁有“個體大、多年生、耐低光”三大特征,可作為我國構建海藻場的優良藻種。
氮、磷營養鹽在大型海藻生長和發育過程中扮演著重要角色。研究發現,水體中缺乏氮、磷營養鹽時可嚴重影響海帶(Saccharinajaponica)配子體的形成時間和雌雄分化,在添加氮、磷營養鹽的人工海水中胚孢子大部分能正常形成配子體,且氮、磷的配比對海帶的發育也有一定影響[3]。孫菊燕[4]研究發現,在過高和過低氮、磷劑量下細基江蘺(Gracilariatenuistipitata)藻體內藻紅素、葉綠素a、蛋白質含量降低,生長速率明顯減慢。硝態氮質量濃度的增加可使龍須菜(Gracilariopsislemaneiformis)藻體的色素和可溶性蛋白含量及營養鹽吸收速率均顯著提高,即在海水富營養化條件下,龍須菜的生長、生化成分和營養鹽的吸收都會受到影響[5]。
海藻的分布、生長、發育和繁殖等生命過程均會受到溫度、光照、營養鹽等環境因子的影響,尤其是海藻幼苗對各種環境因子變化相對敏感,探究幼苗生長發育的最適環境條件,對開展海藻的栽培意義重大。極北海帶幼苗在早期對環境條件(溫度、光照、營養鹽)的需求和響應的研究較少,目前尚未見硝態氮對極北海帶幼苗生長、生理生化影響的報道。筆者以配子體克隆培育的極北海帶幼苗為試驗對象,通過分析氮營養鹽的不同水平對極北海帶生長和生化特性的影響,研究適宜幼苗生長的最優營養鹽條件,以期為極北海帶苗種生產、人工養殖及海藻場建設提供理論參考。
1 材料與方法
1.1 試驗材料
試驗用極北海帶配子體來源于法國布列塔尼半島海區,其幼苗通過配子體克隆技術培育約2個月獲得。幼苗培養條件為:溫度13 ℃,光合有效輻射為40~50 μmol/(m2·s),光周期12L∶12D,天然海水經過濾、高壓滅菌冷卻后添加營養鹽(PO43--P:0.4 mg/L,NO3--N:4 mg/L)作為培養液。
1.2 試驗方法
選取藻體長度為3~5 cm、無褶皺損傷、無病害的健康幼苗作為試驗材料。NO3--N質量濃度梯度為0、0.5、2、4、6、8、10 mg/L,PO43--P:0.4 mg/L。其他培養條件:溫度9 ℃,光合有效輻射40 μmol/(m2·s),光周期12L∶12D。每個處理組至少3個平行。
1.2.1 不同氮營養鹽質量濃度下極北海帶幼苗相對生長速率的測定
挑選健康的極北海帶幼苗,稱量藻體鮮質量,各氮營養鹽質量濃度處理組平均體長為(3.0±0.1)~(5.0±0.1) cm、平均鮮質量為(1.0±0.05) g。每隔3 d換水一次,10 d后稱量鮮質量,相對生長速率(RRG)按下式計算:
RRG=[ln (mt/m0)/t]×100%
式中,m0為初始藻體的鮮質量(g),mt為試驗結束時藻體的鮮質量(g),t為試驗持續的時間(d)。
1.2.2 生化指標的測定
極北海帶藻體鮮質量測定結束后,立刻用液氮將不同處理組的極北海帶幼苗速凍并儲存在超低溫冰箱中。本試驗測定的生化指標包括:色素(葉綠素a和類胡蘿卜素)、可溶性蛋白質、超氧化物歧化酶、過氧化物酶、過氧化氫酶、丙二醛、超氧陰離子。
根據文獻[6-7]中的方法,測定葉綠素a和類胡蘿卜素的含量,其他指標的測定均采用酶聯免疫吸附分析法。酶聯免疫分析檢測試劑盒購自上海酶聯生物科技有限公司,嚴格按照試劑盒說明書進行各個指標的測定。超氧化物歧化酶、過氧化物酶和過氧化氫酶的活性單位以U/mg表示[8]。
1.3 數據處理
采用SPSS 18.0統計軟件進行單因素方差分析、Duncan多重比較及Pearson相關性分析(P<0.05為差異顯著),用Excel軟件繪制圖形。
2 結 果
2.1 不同氮質量濃度下極北海帶幼苗的相對生長速率
氮營養鹽水平對極北海帶相對生長速率有顯著影響(圖1)。2~8 mg/L是極北海帶生長的適宜氮質量濃度,相對生長速率顯著高于其他氮質量濃度,分別為8.50%/d~9.48%/d,該質量濃度范圍內的相對生長速率無顯著差異。氮質量濃度為0.5、10 mg/L時,極北海帶相對生長速率分別為5.87%/d、4.75%/d,顯著低于最適宜質量濃度,但顯著高于氮質量濃度0 mg/L(相對生長速率為3.54%/d),說明過高或過低的氮質量濃度均會限制極北海帶的相對生長速率。試驗周期內,在不同氮質量濃度下極北海帶藻體顏色和形態無顯著變化。
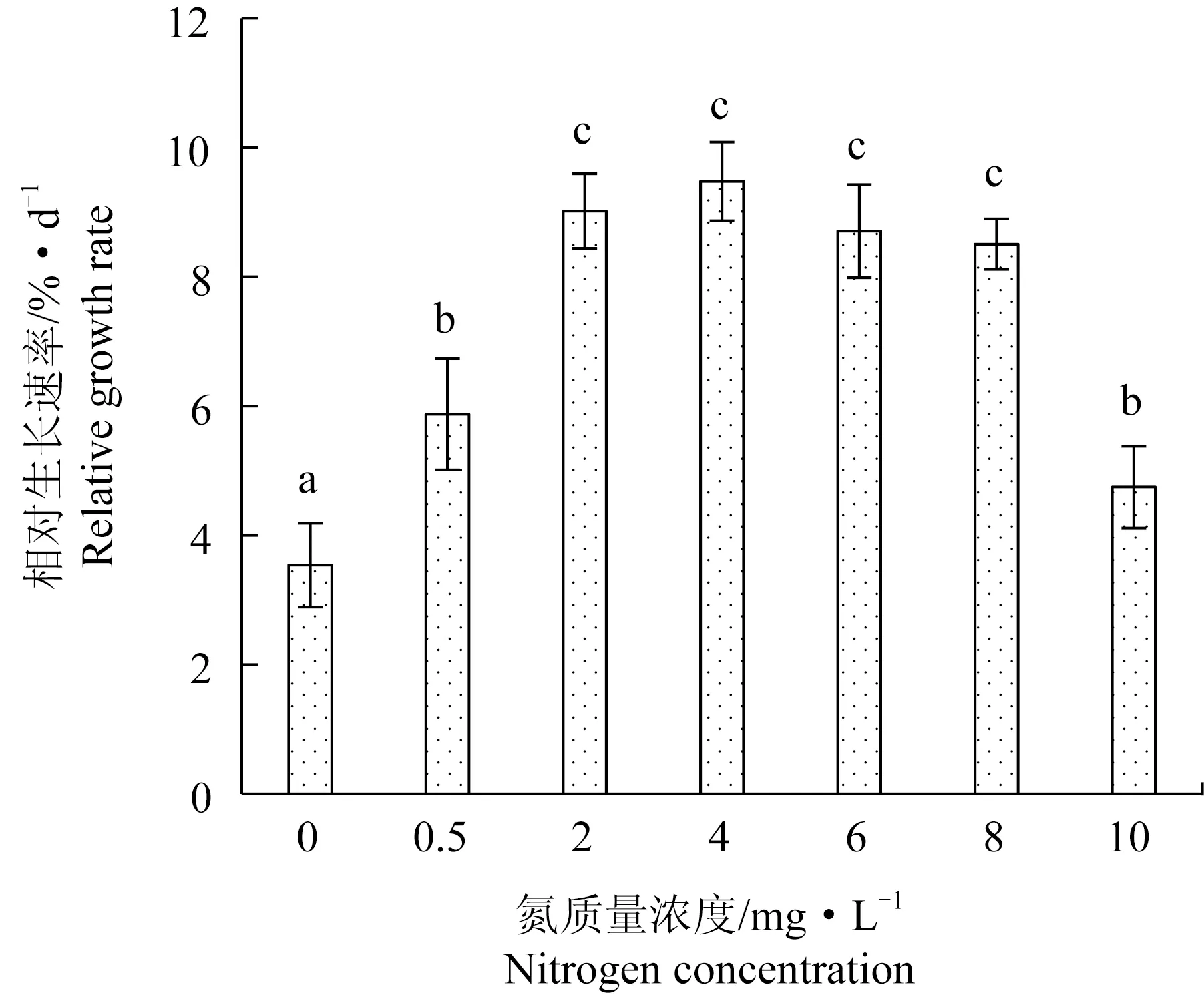
圖1 不同氮質量濃度下處理10 d后極北海帶幼苗的相對生長速率Fig.1 The relative growth rates of young kelp L.hyperborea seedlings exposed to different nitrate nitrogen concentration for 10 days柱狀圖上的不同字母表示在0.05水平具有顯著性差異(P<0.05),下同.The means different letters on the histogram are significant difference at the level of 0.05 (P<0.05),et sequentia.
2.2 不同氮質量濃度下極北海帶幼苗的丙二醛和超氧陰離子含量
方差分析結果表明,氮質量濃度的變化對極北海帶幼苗的丙二醛含量有顯著影響。培養液中氮質量濃度為0和0.5 mg/L時,極北海帶幼苗丙二醛含量較高,顯著高于氮質量濃度2~10 mg/L。氮質量濃度為8 mg/L時,丙二醛含量最低,其他各組間均無顯著差異(圖2)。Pearson相關性分析表明,丙二醛含量與相對生長速率呈顯著負相關(P<0.05)。
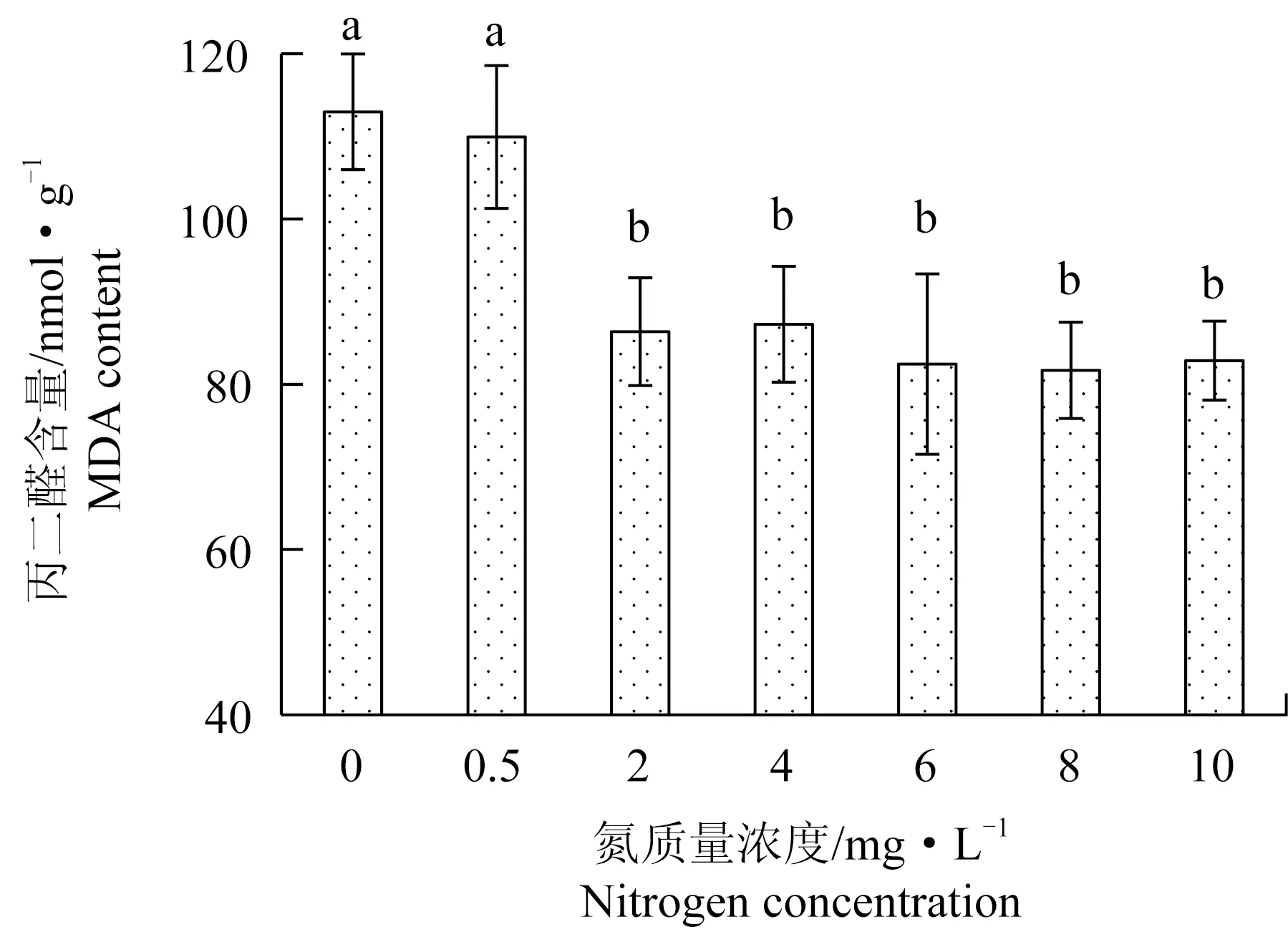
圖2 極北海帶幼苗在不同氮質量濃度下的丙二醛含量Fig.2 MDA content of young kelp L.hyperborea seedlings exposed to different nitrate nitrogen concentrations
氮質量濃度變化對極北海帶幼苗的超氧陰離子含量有顯著影響。氮質量濃度在4~10 mg/L時,超氧陰離子含量無顯著性差異且含量較低,而在低氮質量濃度(0~0.5 mg/L)下,超氧陰離子含量顯著升高(圖3)。Pearson相關性分析表明,超氧陰離子含量與丙二醛含量呈極顯著正相關(P<0.01),說明超氧陰離子的積累,引起細胞膜脂質過氧化,進而導致丙二醛含量上升。
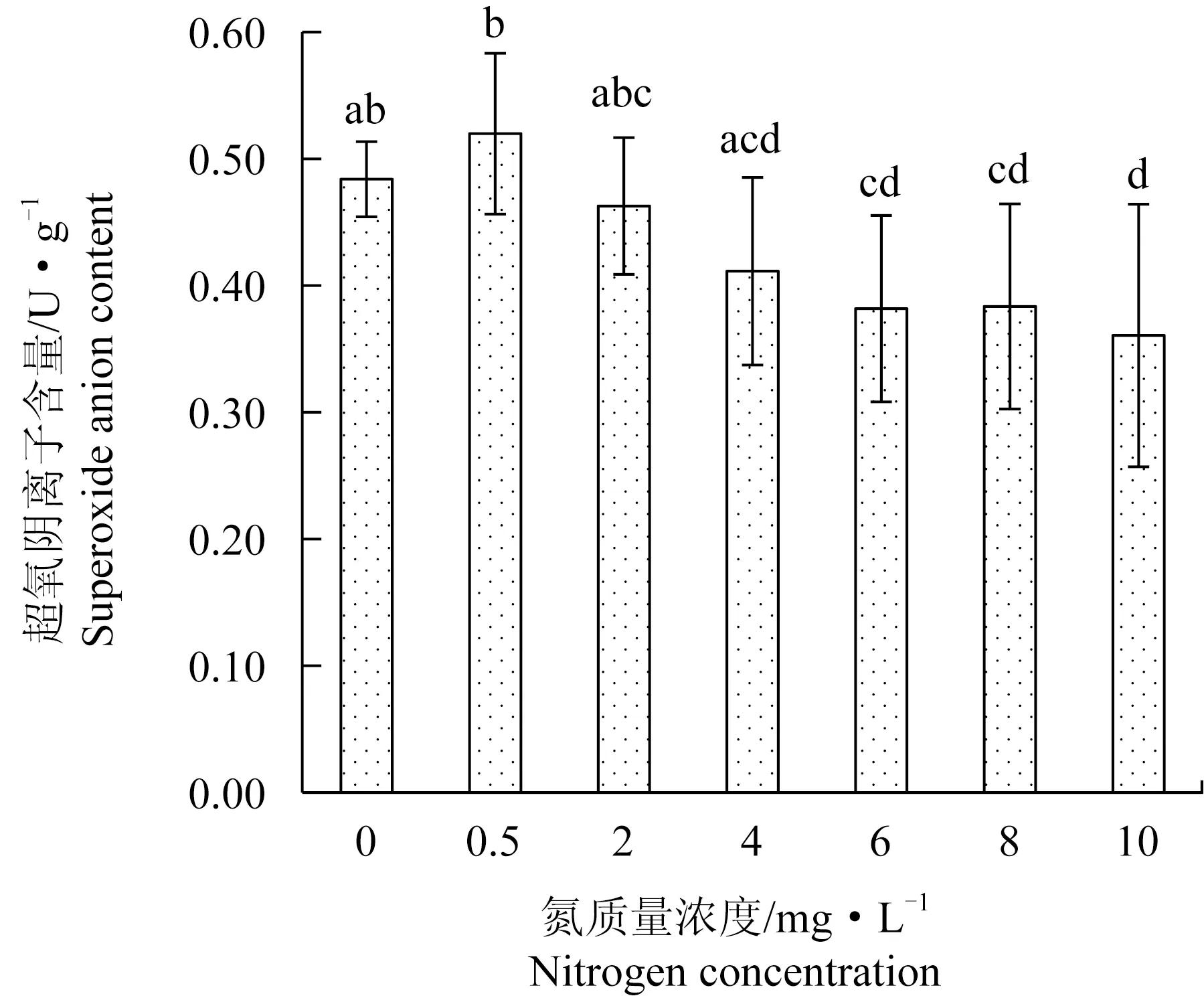
圖3 極北海帶幼苗在不同氮質量濃度下的超氧陰離子含量Fig.3 Superoxide anion content of young kelp L.hyperborea seedlings exposed to different nitrate nitrogen concentration
2.3 不同氮質量濃度下極北海帶幼苗的可溶性蛋白含量
極北海帶幼苗可溶性蛋白含量隨著氮質量濃度的增加呈升高趨勢。可溶性蛋白含量最高值位于氮質量濃度為6、8 mg/L時,分別為14.63、14.57 mg/g,均高于氮質量濃度為4、10 mg/L時的含量,但無顯著性差異。0~2 mg/L氮質量濃度組的可溶性蛋白含量顯著低于氮質量濃度為4~10 mg/L組,且可溶性蛋白含量隨著氮質量濃度的增加顯著升高(圖4)。Pearson相關性分析表明,可溶性蛋白含量與相對生長速率呈極顯著正相關(P<0.01)。
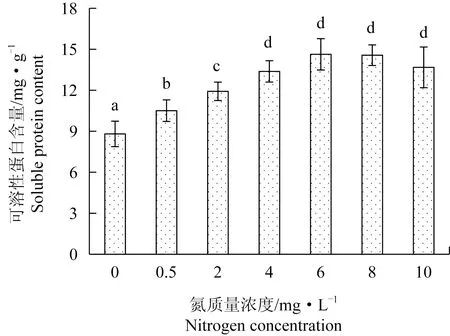
圖4 極北海帶幼苗在不同氮質量濃度下的可溶性蛋白含量Fig.4 Soluble protein content of young kelp L.hyperborea seedlings exposed to different nitrate nitrogen concentration
2.4 不同氮質量濃度下極北海帶幼苗的抗氧化酶(超氧化物歧化酶、過氧化物酶和過氧化氫酶)活性
氮質量濃度變化對極北海帶幼苗超氧化物歧化酶、過氧化物酶和過氧化氫酶活性均有顯著影響,即適宜的氮質量濃度下活性低,高氮或低氮質量濃度下活性高。氮質量濃度為6 mg/L時極北海帶幼苗超氧化物歧化酶活性最低,顯著低于氮質量濃度0、0.5、8、10 mg/L,與氮質量濃度為2、4 mg/L時無顯著差異。氮質量濃度為10 mg/L時超氧化物歧化酶活性顯著高于0.5、2、4 mg/L。氮質量濃度為6 mg/L時極北海帶幼苗的過氧化物酶活性最低,顯著低于氮質量濃度0、0.5、10 mg/L。氮質量濃度為0 mg/L時過氧化物酶活性最高,顯著高于其他各組。氮質量濃度為10 mg/L時過氧化物酶活性顯著高于0.5 mg/L,其他各組間過氧化物酶活性無顯著性差異。極北海帶幼苗的過氧化氫酶活性在氮質量濃度為2、4、6、8 mg/L時均較低,其中,氮質量濃度為4、6、8 mg/L時極北海帶幼苗過氧化氫酶活性顯著低于氮質量濃度0、0.5、10 mg/L。氮質量濃度為2 mg/L時顯著低于10 mg/L,其他各組間無顯著性差異(圖5)。Pearson相關性分析表明,過氧化物酶與超氧化物歧化酶、過氧化氫酶均呈顯著性正相關(P<0.05),說明三者對氮質量濃度的變化具有相似的響應趨勢。

圖5 極北海帶幼苗在不同氮質量濃度下的超氧化物歧化酶、過氧化物酶、過氧化氫酶活性Fig.5 Specific activities of SOD,POD and CAT of young kelp L.hyperborea seedlings exposed to different nitrate nitrogen concentrations柱狀圖上的字母下標為相同數字的作為同一組,進行多重比較,同一組的不同字母表示在P<0.05水平具有顯著性差異,下同.The letters with the same number subscript on the histogram indicate the same group,and the different letters in the same group show significant difference at the level of P<0.05,et sequentia.
2.5 不同氮質量濃度下極北海帶幼苗的色素(葉綠素a和類胡蘿卜素)含量
氮質量濃度對極北海帶幼苗葉綠素a和類胡蘿卜素含量有顯著的影響。極北海帶幼苗葉綠素a的含量隨氮質量濃度的增加呈現先升后降的趨勢,氮質量濃度為4、6 mg/L時極北海帶葉綠素a含量無顯著性差異,均顯著高于其他質量濃度組。氮質量濃度為10 mg/L時極北海帶葉綠素a含量最低,顯著低于2~8 mg/L質量濃度組,其他各組間無顯著性差異。極北海帶幼苗類胡蘿卜素含量在氮質量濃度為4 mg/L時最高,顯著高于0、10 mg/L組,極北海帶幼苗類胡蘿卜素含量在氮質量濃度10 mg/L時最低,顯著低于除對照外的其他氮質量濃度組,其他各組間無顯著性差異(圖6)。
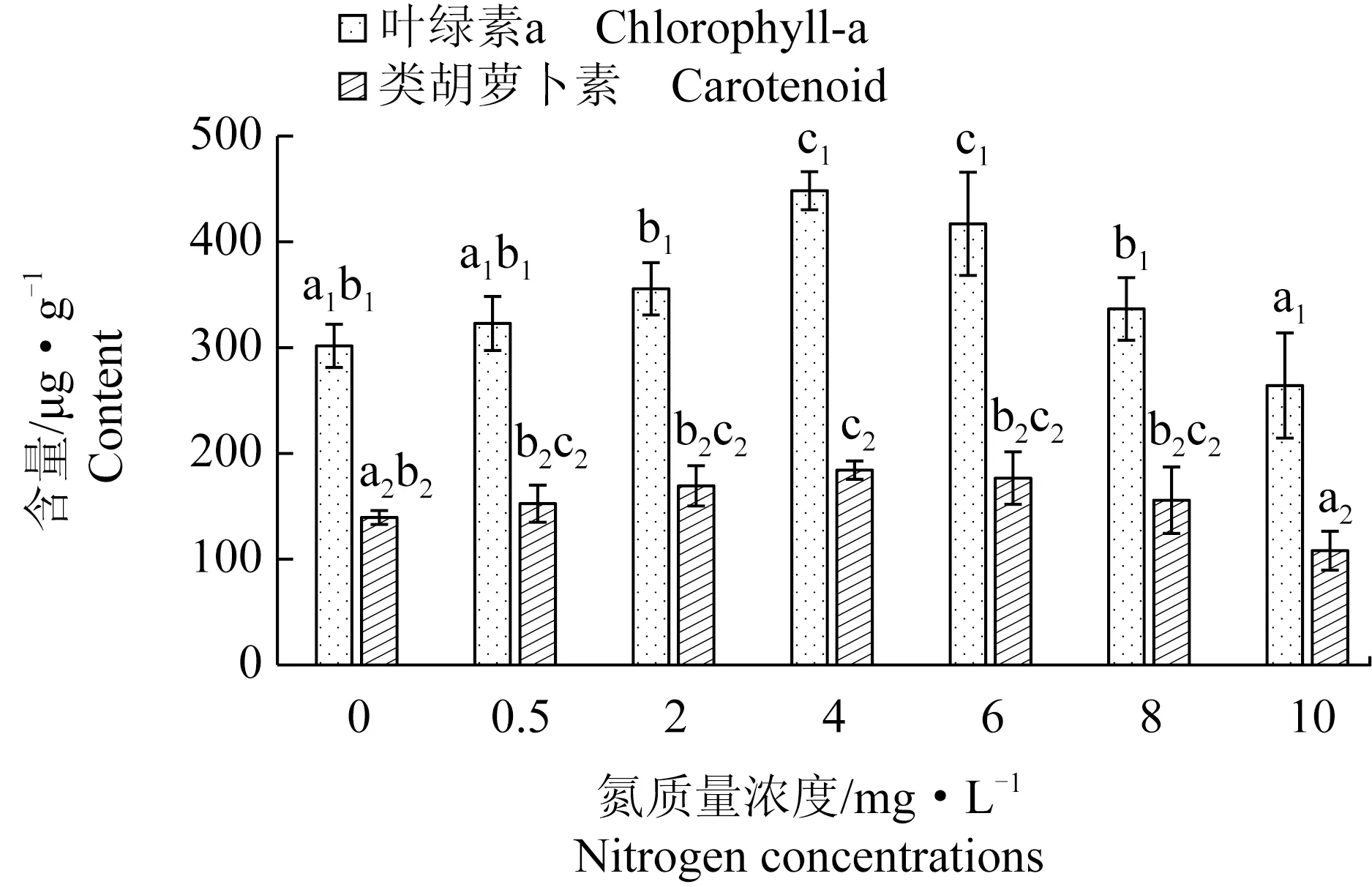
圖6 極北海帶幼苗在不同氮質量濃度下的葉綠素a和類胡蘿卜素含量Fig.6 Chlorophyll-a and carotenoid contents of young kelp L.hyperborea seedlings exposed to different nitrate nitrogen concentrations
3 討 論
極北海帶自然分布于大西洋東北部沿岸,其分布、生長、發育及繁殖等生命過程受到溫度、光照度、營養鹽含量等環境因子的影響,尤其是幼苗階段對各種環境因子變化相對敏感[9]。筆者主要討論氮營養鹽含量對極北海帶幼苗生長和生化成分的影響,為探究極北海帶的苗種繁育、人工養殖和對富營養化水域的調控能力提供參考。
3.1 氮營養鹽質量濃度對極北海帶幼苗生長的影響
氮、磷營養鹽在大型海藻的生長發育過程中具有重要的作用,研究發現,氮是藻類光合器官的重要組成成分,是藻體生長的營養基礎,氮營養鹽的增加能夠為以氮為基本元素的各種蛋白和化合物的合成提供原料,氮的加富必然促進藻體內氮代謝反應,從而導致氮同化物的積累,在氮充足的條件下,酶類、氨基酸、葉綠素、藻膽蛋白可大量合成而作為藻體的氮庫[10-13]。硝態氮增加可促進龍須菜的生長速率,提高色素和可溶性蛋白含量[7]。水體營養水平較高時,龍須菜色素、蛋白質及總氮含量均較高,且生長速度快[14]。海水總無機氮含量過低、氮磷比過高以及氨氮在總無機氮中所占比例較低等條件下,均可導致芋根江蘺(G.blodgettii)藻體營養不良、生長緩慢[15]。海帶幼孢子體生長的適宜氮營養鹽質量濃度為4~8 mg/L[16]。在本試驗中,氮質量濃度4~6 mg/L時極北海帶幼苗的葉綠素a含量顯著高于其他各質量濃度,氮質量濃度2~8 mg/L時幼苗的相對生長速率均較高,氮質量濃度4~8 mg/L時可溶性蛋白含量顯著高于其他質量濃度組且與相對生長速率呈極顯著正相關,因此,可以推斷極北海帶生長最適宜的氮質量濃度為2~8 mg/L,這與Kain[9]報道的歐洲沿岸野生極北海帶適宜氮含量以及我國養殖海帶適宜的氮含量范圍基本相同。
3.2 氮營養鹽質量濃度對極北海帶幼苗生化成分的影響
植物體內可溶性蛋白質大多是酶類,參與細胞的各種代謝活動,其含量是反映植物體總代謝的一個重要指標[17]。超氧陰離子是活性氧的一種,植物體內活性氧的積累會導致氧化脅迫,對細胞的傷害之一是啟動膜脂過氧化連鎖反應,使維持細胞區域化的膜系統受損或瓦解[18]。膜脂過氧化即非脂性自由基如超氧陰離子對類脂中不飽和脂肪酸引起的一系列自由基反應,一般被認為是活性氧對細胞膜系統傷害的原初機理,丙二醛(MDA)是膜脂過氧化的主要產物之一,常被作為判斷膜脂過氧化的一種主要指標[18]。弱光、高光脅迫下極北海帶幼苗體內超氧陰離子和丙二醛含量均高于適宜光照時的含量[19]。黃鶴忠等[20]發現,細基江蘺繁枝變型(G.tenuistipitatavar.liui)在低硝態氮和高硝態氮的營養鹽水平下丙二醛含量顯著高于中等硝態氮含量組,表明過高或過低的硝態氮含量均會對江蘺的生理生長產生不利影響。本試驗中,在低氮營養鹽質量濃度0、0.5 mg/L時極北海帶幼苗的超氧陰離子和丙二醛含量均高于其他質量濃度組且與丙二醛含量呈極顯著正相關,丙二醛含量與相對生長速率呈顯著負相關。因此,低營養鹽含量時極北海帶可能處于非最佳生理狀態,導致體內超氧陰離子積累,使細胞處于氧化脅迫狀態,膜脂過氧化對細胞膜造成了一定的傷害,進而導致極北海帶的相對生長速率降低。
活性氧對細胞膜系統的損傷不可修復,植物體的抗氧化系統使活性氧含量處于不斷產生和清除的動態平衡中。植物本身的抗氧化系統主要包括酶促抗氧化系統(超氧化物歧化酶、過氧化氫酶、過氧化物酶、谷胱甘肽還原酶等)和非酶促抗氧化系統(抗壞血酸與類胡蘿卜素以及一些含巰基的低分子化合物等),遭受環境脅迫時,活性氧代謝平衡被破壞,酶促和非酶促抗氧化系統共同作用,清除積累的活性氧,減少對細胞的損害[21-22]。試驗表明,不同氮、磷質量濃度對海藻的超氧化物歧化酶和過氧化物酶等抗氧化酶活性、膜脂過氧化產物等均產生顯著影響。龍須菜藻體的超氧化物歧化酶、過氧化物酶和過氧化氫酶活性隨氮磷質量濃度的降低而逐漸升高,高氮質量濃度下長時間培養時酶活性降低,抗氧化系統作用降低,藻體可能會面臨死亡[23]。在最適宜的營養鹽條件下,芋根江蘺在生化組分(光合色素及可溶性蛋白)和抗氧化能力等方面都表現較好,過低或過高質量濃度的無機氮均使得丙二醛、過氧化物酶活性升高,影響藻體生長[15]。逐漸增加水體中硝態氮含量,細基江蘺繁枝變型的硝酸還原酶、超氧化物歧化酶和過氧化物酶的活性均顯著呈現先增后降的趨勢,且與丙二醛呈相反的趨勢[20]。本試驗中,在低氮質量濃度(0~0.5 mg/L)時,極北海帶幼苗的超氧陰離子水平明顯升高,此時超氧化物歧化酶、過氧化物酶和過氧化氫酶的活性均升高,尤其是過氧化物酶活性,與氮質量濃度為0 mg/L時相比升高較明顯,表明這3種抗氧化酶尤其是過氧化物酶在清除藻體內自由基時起著關鍵作用。而在低氮處理組中,丙二醛有較明顯的積累,表明在低氮營養鹽水平下抗氧化酶的作用可能受到限制。在高氮環境(10 mg/L)時,極北海帶幼苗的超氧陰離子并未表現出較高的水平,同時丙二醛也無明顯的積累,這可能與3種抗氧化酶活性在高氮環境時均明顯升高有關。由此推測,幼苗雖然均處于非最佳生理狀態,但高氮環境和低氮環境對極北海帶幼苗的抗氧化酶系統的影響并不相同。
3.3 氮營養鹽質量濃度對極北海帶幼苗色素含量的影響
類胡蘿卜素是非抗氧化系統重要的組成部分,在所有的單線態氧猝滅系統中,類胡蘿卜素具有最重要的生物學意義。類胡蘿卜素存在于葉綠體內,一方面阻止激發態葉綠素分子的激發從反應中心向外傳遞;另一方面,它也保護葉綠素分子免遭光氧化損傷[24]。本試驗脅迫條件為高氮和低氮劑量時,類胡蘿卜素含量均無顯著升高,表明其對氮脅迫無顯著響應。幼苗的葉綠素a含量在4~6 mg/L時顯著高于其他各劑量組,因此極北海帶生長最適宜的氮質量濃度為4~6 mg/L。在非最適的氮環境(高氮和低氮)時,幼苗葉綠素a含量降低,對藻體的光合作用產生影響,從而使幼苗生長速率降低。
猜你喜歡
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
小讀者(2021年2期)2021-03-29 05:03:48
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
中國生殖健康(2019年2期)2019-08-23 08:12:08
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
產品可靠性報告(2017年7期)2017-09-05 09:49:12
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
