vvi-miR172s及其靶基因響應赤霉素調控葡萄果實發育的作用分析
2021-03-25 08:44:04宣旭嫻盛子璐解振強黃雨晴鞏培杰張川鄭婷王晨房經貴
中國農業科學 2021年6期
宣旭嫻,盛子璐,解振強,黃雨晴,鞏培杰,張川,鄭婷,王晨,房經貴
vvi-miR172s及其靶基因響應赤霉素調控葡萄果實發育的作用分析
宣旭嫻1,盛子璐1,解振強2,黃雨晴1,鞏培杰1,張川1,鄭婷1,王晨1*,房經貴1
1南京農業大學園藝學院,南京 210095;2江蘇農林職業技術學院,江蘇句容 212499
【】miR172是植物生長發育的重要調節因子,闡釋vvi-miR172s及其靶基因應答赤霉素在葡萄果實不同組織發育過程中的作用,從miRNA角度認識GA調控葡萄果實發育的作用機制。以‘白羅莎里奧’葡萄為材料,以miR-RACE和PCR技術克隆vvi-miR172a/b/c/d的成熟體和前體序列,由psRNA Target軟件預測vvi-miR172s的靶基因;利用RLM-RACE技術驗證vvi-miR172s剪切靶基因的裂解作用及其作用位點;采用生物信息學軟件對靶基因、靶蛋白進行系統進化、序列結構分析及亞細胞定位預測;通過在線軟件PLANTCARE進行啟動子作用元件分析;通過qRT-PCR和芯片數據分析vvi-miR172s和靶基因在不同器官、不同發育階段的基因表達圖譜,以及應答GA3在果實不同組織中的時空表達模式。克隆鑒定了vvi-miR172a/b/c/d的成熟體和前體序列,預測到和四個靶基因,驗證到裂解產物及其裂解位點,證明它們為vvi-miR172s的真實靶基因。序列結構分析顯示,均含有10個外顯子和9個內含子,其motif元件的種類和數量相似,均含有兩個排列順序相近的AP2蛋白結構域,表明其結構和功能具有一定的保守性;且、和與楊樹的親緣關系較近,與大豆具有較高的同源性;靶基因蛋白二級結構均為-螺旋,可進一步折疊為穩定的三級結構;4個靶蛋白亞細胞定位主要位于細胞核內。啟動子作用元件分析發現vvi-miR172c和中含有赤霉素響應元件,表明它們可能響應赤霉素調控葡萄生長發育;芯片數據分析顯示vvi-miR172c和靶基因的表達模式具有組織或器官特異性,且它們之間呈現一定的負相關,表明它們之間存在負調控作用;RT-qPCR結果顯示,隨著葡萄果實的發育,果皮中vvi-miR172a/b/d呈下降的表達趨勢,而靶基因的表達模式相反,表明vvi-miR172a/b/d對負調控;果肉中vvi-miR172d的表達水平降低,而靶基因的表達水平增加,表明vvi-miR172d與呈負相關性。另外,GA3處理改變了vvi-miR172s的靶模式,增強了果肉和果皮組織中vvi-miR172d與的負相關性,同時誘導了vvi-miR172c對的負調控作用。均為葡萄miR172a/b/c/d的真實靶基因,vvi-miR172家族可能通過vvi-miR172a/b/d和vvi-miR172d分別介導靶基因和調控果皮和果肉組織的發育過程,而vvi-miR172c和vvi-miR172d及其靶基因可能是GA調控葡萄果皮與果肉組織發育的主要作用因子。
葡萄;果實發育;赤霉素;miR172;靶基因
0 引言
【研究意義】葡萄(L.)是世界四大水果之一,在我國廣泛栽培。生產上,通常利用GA3調控葡萄花果發育,提高果實品質從而增加其經濟效益。miR172是一個重要的保守miRNA家族,其家族成員的功能具有多樣性,在植物成花作用[1]、種胚發育[2]以及非生物脅迫的響應[3]等方面發揮重要作用。近期研究發現,miR172可能響應GA參與葡萄花果發育的調控[4],但其對葡萄果實不同組織的調控作用尚不清楚。從miR172角度研究GA調控葡萄果實發育的作用機制,為深入認識vvi-miR172s的功能及其應答GA的分子調控機制提供重要信息。【前人研究進展】miR172廣泛存在于植物中,不同物種中具有多個,可以在不同的組織和不同的發育階段形成保守的成熟miR172序列。模式植物擬南芥中5個加工后形成3個miR172成熟體[5];楊樹有9個和4個miR172成員[6];水稻含4個和2個miR172[7];油菜含有4個miR172成員[1];玉米中僅包括一個miR172成員[8]。擬南芥中miR172的過表達會導致非正常花的發育,還會使植物提早開花[9];miR172對擬南芥果實(長角果)的生長具有促進作用;miR172的過表達會使蘋果果實顯著變小[10];麻瘋樹種JcmiR172a的過表達增強木質部的發育,減少韌皮部的發育[7];據報道,miR172調控小麥穗型和脫粒率研究中發現miR172的過表達會誘導穗的加長和降低脫粒率[10]。另有研究發現,OsmiR172b是調控水稻種子發育表型最顯著的成員[2],對種子胚發育具有負調節作用。這些結果表明,miR172家族在不同植物中的功能具有多樣性。成熟miR172可以通過序列互補靶向AP2基因家族的mRNA,并且可以通過抑制翻譯或引發靶mRNA的降解來抑制靶基因的表達。目前,在擬南芥[5]、水稻[3]、油菜[1]等植物中均發現AP2家族轉錄因子是miR172的靶基因。miR172及其靶基因在開花時間和花器官分化中起關鍵作用。在擬南芥中,除外的所有miR172靶基因過量表達均延遲開花[11-13]。相反,花椰菜中雙重突變體,四重突變體和六倍體突變體均導致提早開花,且開花時間隨突變基因數量的增加而延長[14-15]。另有研究發現AP2類轉錄因子還參與番茄果實的轉色[16]、成熟[17]及類胡蘿卜素[18]積累過程,且擬南芥中、[19]可調控種子的發育。此外,AP2/ERF類轉錄因子還被視為植物激素信號連接的關鍵調節器,可參與赤霉素、生長素、細胞分裂素、茉莉酸、乙烯、脫落酸等多種激素信號轉導途徑[20]。【本研究切入點】赤霉素(GA)是調控葡萄果實發育如無核膨大等的關鍵生長調節劑,但其相關分子機制尚不甚清晰[21]。如上所述,miR172和AP2的研究主要集中在植物開花時間和花器官的發育方面。在擬南芥[22-23]和大豆[24]的研究中已證明AP2類轉錄因子與種子發育相關,但其參與果皮、果肉發育的研究甚少,更未見miR172通過應答GA靶向AP2類轉錄因子參與葡萄果皮、果肉等膨大發育關鍵組織調控的報道。【擬解決的關鍵問題】本試驗以miRBase數據庫為基礎,鑒定葡萄中miR172c家族成員及其靶基因,系統分析miR172成員及其靶基因的染色體定位、進化關系、基因結構、保守基序和上游順式作用元件;采用實時熒光定量PCR(qRT-PCR)技術,結合芯片數據,明確vvi-miR172成員及其靶基因應答GA3在葡萄果實不同組織中的表達特性,為深入研究其在果皮、果肉組織發育過程中的調控機制提供重要參考。
1 材料與方法
試驗于2017年在江蘇省句容農博園葡萄基地進行。
1.1 材料與處理
本試驗以6年生優質‘白羅莎里奧’葡萄為試驗材料,于盛花前10 d用50mg·L-1的GA3浸蘸花穗30s,以清水處理為對照。每組處理隨機選定12株長勢較為一致的植株,分別于幼果期(花后10 d,10 DAF)、硬核期(35 DAF)、第2次膨大期(60 DAF)、近成熟期(85 DAF)隨機采集生長較為一致的葡萄果實,將果皮、果肉分離,用液氮速凍并保存于-80℃冰箱中。
1.2 葡萄miR172s成熟體序列及前體序列的克隆鑒定
在miRBase數據庫(http://www.mirbase.org/)搜索并下載葡萄miR172s(vvi-miR172a/b/c/d)前體及成熟體序列。基于成熟體序列,參考Wang等[25]的方法,設計特異性引物,利用miR-RACE技術在‘白羅莎里奧’果實組織中克隆vvi-miR172a/b/c/d的成熟體序列,測序后鑒定其精確序列。在miRBase數據庫下載水稻(Japonica)、玉米()、擬南芥()、大豆()、楊樹()、煙草()、馬鈴薯()和番茄()miR172家族的成熟體序列,與葡萄miR172s(vvi-miR172a/b/c/d)進行序列比對。利用下載的vvi-miR172a/b/c/d前體序列,分別向上、向下擴展約200 bp,預測葡萄miR172a/b/c/d的前體基因序列,并設計特異性引物進行PCR擴增(表1)。反應體系為50 μL:上、下游引物各2 μL,cDNA 2 μL,10×PCR buffer(Mg2+plus)5 μL,dNTP Mixture 4 μL,Ex-Taq酶0.50 μL,ddH2O 34.5 μL。反應程序:95℃ 5 min;95℃ 30 s,59℃ 30 s,72℃ 30 s,35個循環;72℃ 10 min。擴增產物用1.5%的瓊脂糖凝膠電泳檢測正確后回收目的片段,連接至pMD19T載體進行TA克隆,克隆后的純化產物由上海生工生物工程股份有限公司進行測序。
1.3 葡萄miR172s靶基因預測及生物信息學分析
基于鑒定的vvi-miR172a/b/c/d成熟體序列,由在線軟件psRNA Target(http://plantgrn.noble.org/psRNATarget/)進行靶基因預測,運行參數均為默認值。根據預測的結果,從葡萄基因數據庫CRIBI(http://genomes.cribi.unipd.it/grape/index.php)獲得靶基因的功能注釋及基因序列。運用在線工具MG2C(http://mg2c.iask.in/mg2c_v2.0/)繪制染色體定位圖;利用Gene Structure Display Server(GSDS)(http:// gsds.cbi.pku.edu.cn/index.php)進行基因序列結構分析,繪制基因結構示意圖;利用DNAMAN 6.0和Clustal X 2.1軟件進行多序列比對;使用MEGA37.0軟件,選用鄰接法(Neighbor-Joining)構建系統進化樹;采用在線工具MEME(http://meme-suite.org/tools/ meme)鑒定蛋白保守基序,基序數量設置為15,其余參數為默認值;使用在線數據庫CDD(http://www. ncbi.nlm.nih.gov/cdd)進行保守結構域分析,采用IBS軟件作圖[26];利用在線軟件PRABI(http://www.prabi. fr/)進行蛋白質二級結構進行預測;根據在線軟件SWISS-MODEL(https://swissmodel.expasy.org/interactive)進行蛋白質三級結構分析;運用在線軟件BUSCA(http://busca.biocomp.unibo.it)進行亞細胞定位預測。
1.4 葡萄miR172s及其靶基因啟動子作用元件分析
利用葡萄基因組數據庫CRIBI(http://genomes.cribi.unipd.it/grape/index.php)和Grape Genome Browse(http://www.genoscope.cns.fr/externe/GenomeBrowser/Vitis/)在線軟件,確定vvi-miR172a/b/c/d前體基因上游1 500 bp及4個靶基因轉錄起始位點(ATG)上游1 500 bp序列,利用在線軟件PLANTCARE (http:// bioinformatics.psb.ugent.be/webtools/plantcare/html/)進行啟動子順式作用元件分析。
1.5 芯片數據分析
從NCBI Genesis Expression Omnibus(GEO)下載GSE36128系列(https://www.ncbi.nlm.nih.govlgeo/ query/acc.cgi?acc=GSE36128)中葡萄不同組織、不同發育階段的芯片表達譜[27]以及GSE59802系列(https:// www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE59802)中70個小RNA文庫數據。兩者表達數據都基于歐亞種葡萄品種‘Corvina’(clone 48)[28],將葡萄不同組織、不同發育階段中每個基因的平均表達值進行歸一化,按log2的值計算vvi-miR172s和靶基因的表達數據。
1.6 vvi-miR172家族成員及其靶基因的表達特性分析
采用CTAB法提取4個不同發育時期葡萄果皮、果肉中的RNA,利用Primer 3 Input(http://bioinfo.ut. ee/primer3-0.4.0/)設計引物(表1),以反轉錄后的cDNA為模板,以為內參基因,對vvi-miR172s前體基因及4個靶基因進行qRT-PCR檢測。擴增體系及程序參考TaKaRa公司的SYBR? Premix Ex TaqTM Ⅱ試劑盒說明書,設置3次生物學重復。采用2-ΔΔCt計算各基因的相對表達量,采用SPSS軟件對數據進行統計及差異顯著性分析(<0.05)。

表1 引物序列及用途
下劃線部分AAGCTT和GGATCC分別指dIII和H I酶切位點
The underlined sequences AAGCTT and GGATCC in the primers represent restriction enzyme sites ofdIII andH I, respectively
2 結果
2.1 赤霉素處理對‘白羅莎里奧’葡萄果實生長發育的影響
GA3處理后,果實橫徑無明顯變化,果實縱徑在花后10、35及60 d顯著增長,且各時期沒有明顯的果核出現,僅在種子區域留下一條木質化的細線;而未經GA3處理的對照組種子正常發育,在花后35 d可觀察到果核,說明GA3處理可以高效誘導‘白羅莎里奧’葡萄產生無核果實。另外,也觀察到外源GA3處理誘導葡萄無核果實的同時,促進了無籽果實的膨大,使其發育成正常商品果大小,甚至更大(圖1),表明GA3在果實發育過程中起著重要調控作用。

圖1 GA3和CK處理下‘白羅莎里奧’葡萄果實生長發育情況
2.2 vvi-miR172家族成員成熟體及前體序列的克隆鑒定
為確定vvi-miR172家族成員在‘白羅莎里奧’葡萄中的真實序列,對其成熟體序列和前體序列進行克隆鑒定。通過miR-RACE獲得vvi-miR172a/b/c/d的成熟體序列,克隆所得的序列與miRBase中的同源序列一致。克隆鑒定的vvi-miR172a/b/c/d前體基因分別為507、495、546和507 bp(圖2-A)。染色體定位結果顯示vvi-miR172家族成員分布在葡萄3條染色體上,vvi-miR172b和vvi-miR172c位于Chr13上,而vvi-miR172a和vvi-miR172d分別位于Chr6和Chr8上(圖2-B)。4種miRNA前體結構顯示出一定差異,但均可形成穩定的莖環結構,且成熟序列均位于莖環結構的3′臂上(圖2-C),表明vvi-miR172a/b/c/d在前體基因上的位置高度保守。多序列比對結果顯示,除vvi- miR172d含23個堿基外,vvi-miR172a、vvi-miR172b以及vvi-miR172c均為21個堿基,且vvi-miR172a和vvi-miR172b僅存在1個堿基的差異(圖2-D)。vvi-miR172四個成熟體序列存在相對保守的區域,介于4—19 bp和21—22 bp,表明miR172成熟體高度保守。vvi-miR172c與ptr-miR172g和sly-miR172d的成熟體序列相同,表明其可能具有相似的生物學功能。
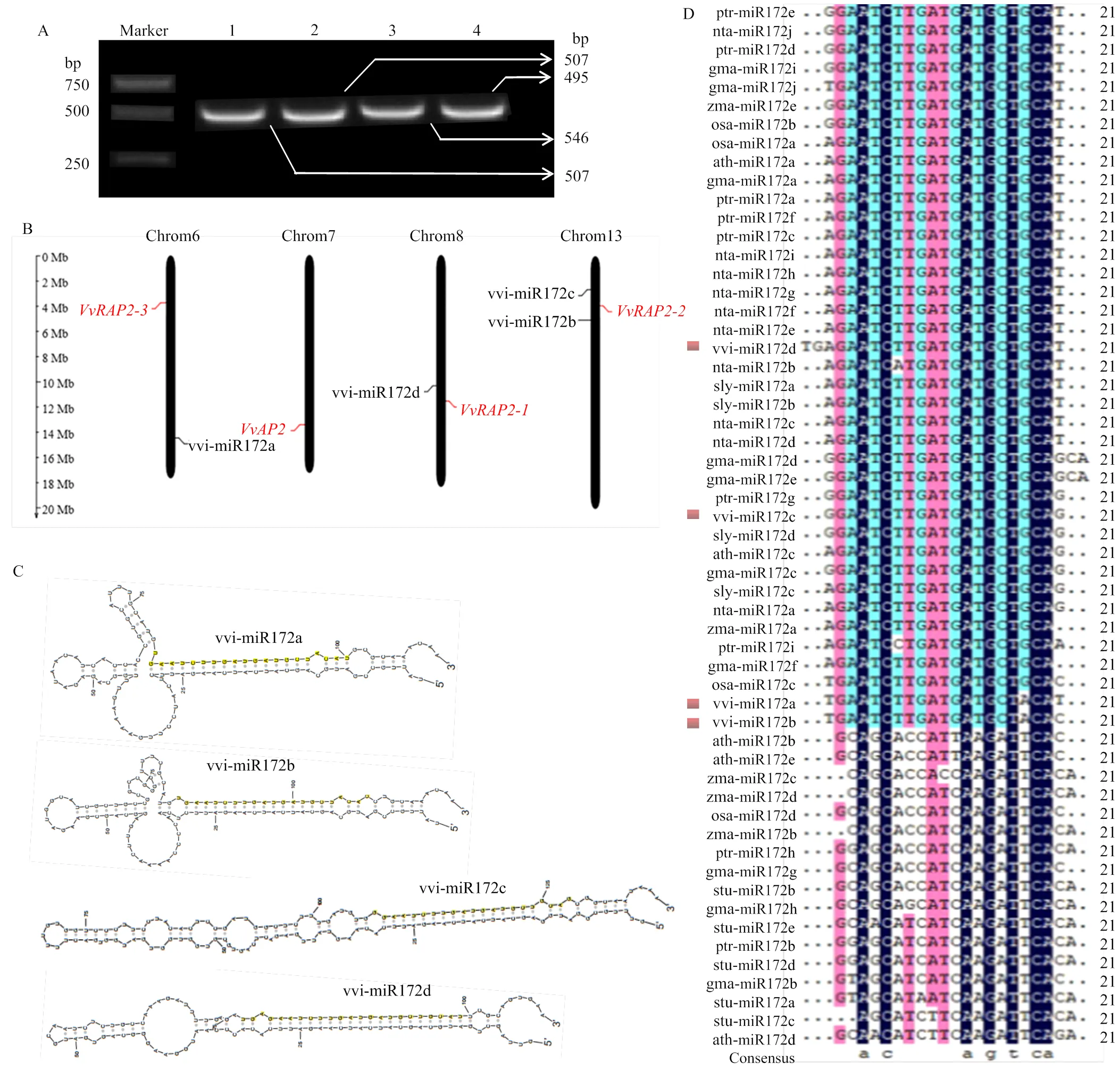
A:vvi-miR172a/b/c/d 前體基因的PCR克隆產物;1:vvi-miR172a;2:vvi-miR172b;3:vvi-miR172c;4:vvi-miR172d;B:vvi-miR172s及其靶基因在染色體上的定位;C:vvi-miR172s前體的二級莖環結構;D:不同物種miR172成熟體序列比對
2.3 vvi-miR172s靶基因的鑒定
2.3.1 vvi-miR172s靶基因的預測及匹配程度分析 根據所獲得的vvi-miR172a/b/c/d成熟體序列,利用在線軟件psRNA Target預測到4條靶基因、、和,分別分布于葡萄基因組的4條染色體上(Chr7、Chr8、Chr13、Chr6)(圖2-B),分析發現vvi-miR172a/b/c/d的靶區都在其靶基因的CDS區,且均通過裂解作用于靶基因(表2)。對vvi-miR172a/b/c/d與其靶基因的匹配程度進一步分析(圖3),發現vvi-miR172a、vvi-miR172b與預測所得靶基因的匹配程度相同,錯配率均為2.5;vvi-miR172c與和的匹配程度最高,錯配率為0.5,與和的錯配率為1.0;vvi-miR172d與的錯配率最高為2.5,與的錯配率分別為0.5、1.5和2.0(表2),說明vvi-miR172家族成員與靶基因間的作用強度存在差異。

表2 葡萄vvi-miR172s及靶基因信息

圖3 vvi-miR172s與靶基因的匹配程度
2.3.2 vvi-miR172s靶基因驗證 利用RLM-RACE技術對vvi-miR172s剪切靶基因的作用位點進行驗證,發現預測的4條靶基因(和)均被驗證到裂解產物,表明它們為vvi- miR172s的真實靶基因(圖4);同時發現vvi-miR172s在和上均只有一個酶切位點,4個靶基因的裂解位點位于vvi- miR172a/b 5′端的第9位堿基(U)和第10位堿基(A)之間,vvi-miR172c/d 5′端的第10位堿基(U)和第11位堿基(A)之間,表明miRNA切割位點的保守性。此外,切割頻率最低,表明其與vvi-miR172a/ b/c/d間的作用強度較弱(圖4)。

虛線代表裂解位點;數字代表mRNA片段5′末端克隆的頻率
2.3.3 靶基因序列結構及其進化保守性分析 基于預測的4條靶基因cDNA編碼的氨基酸序列,利用MEGA7.0軟件對同源比對結果進行進化分析(圖5-A),發現比對結果可分為3組,其中水稻成員和玉米成員分別聚集為一組;、和屬于同一組,而在另一組。且和與楊樹的親緣關系較近,與大豆具有較高的同源性。利用在線軟件GSDS對靶基因序列結構進行分析(圖5-B),發現4個靶基因所含的內含子和外顯子數量相同,均含有10個外顯子和9個內含子。其中,與其他3個靶基因的外顯子長度相似,但內含子長度不同。通過MEME在線軟件預測motif元件(圖5-C),發現各物種內蛋白基序存在一定差異,但各類元件的種類和數量較為穩定,表明AP2及RAP2具有一定的功能保守性。
2.3.4 靶蛋白的結構分析與亞細胞定位預測 利用在線軟件PRABI對vvi-miR172s的靶蛋白進行二級結構預測,發現四者二級結構的表現形式均為-螺旋(圖6-A)。利用SWISS-MODEL進行三維結構同源建模,結果表明其蛋白三維空間排列所形成的蛋白質分子構象具有多樣性[26],其中和的蛋白構象更相似(圖6-B),表明其可能具有功能的相似性。利用在線軟件CDD對靶基因蛋白及其同源序列進行蛋白結構域預測,發現各序列均含有兩個排列順序相近的AP2蛋白結構域(圖6-C),由位置和數量相似的氨基酸編碼。對靶蛋白及同源序列進行亞細胞定位預測(圖6-D),發現4條靶蛋白均位于細胞核中,而和編碼的蛋白還存在于線粒體中。
2.4 vvi-miR172s及其靶基因潛在功能分析
2.4.1 vvi-miR172s前體及靶基因的啟動子作用元件分析 通過分析vvi-miR172s前體基因及其靶基因啟動子作用元件,發現其作用元件大致可分為4類:光信號響應(Gap-box、Box4、GAT-motif等)、激素相關響應(ABRE、TGACG-motif、TATC-box等)、脅迫相關響應(MBS、ARE、GC-motif等)和晝夜周期節律(circadian等)相關元件。葡萄vvi-miR172家族成員及靶基因中光信號響應元件數量最多,作用位點最豐富;激素和脅迫相關響應作用元件數量次之;晝夜周期節律相關元件數量最少(圖7-A)。此外,vvi-miR172s及其靶基因啟動子作用元件的數量和類型也各不相同。在元件數量上,最多,次之,而vvi-miR172a最少;在作用元件類型上,除含有晝夜周期節律相關元件外,其他基因只含有3種作用元件(圖7-A)。為進一步認識vvi-miR172s及其靶基因應答激素反應的潛在機制,分析發現所有vvi-miR172家族成員及靶基因均含有激素響應作用元件,主要包括赤霉素(gibberellin,GA)、生長素(auxin,IAA)、水楊酸(salicylicacid,SA)、脫落酸(abscisic acid,ABA)及茉莉酸甲酯(methyljasmonate,MeJA)響應元件(圖7-B)。但與激素相關的順式作用元件的組成和數量在不同的vvi-miR172成員及其靶基因間均表現出一定的差異,其中,GA響應元件僅存在于和vvi-miR172c中;IAA響應元件僅存在于和中;ABA響應元件存在于4個靶基因中;MeJA和SA響應元件分布較為廣泛(圖7-B)。上述作用元件分析表明vvi-miR172s及其靶基因不僅能響應光、脅迫和晝夜周期節律等外界環境信號,還可能通過參與不同激素信號途徑調控果實的生長發育。
2.4.2 vvi-miR172s及靶基因在葡萄不同器官組織中的時空表達模式分析 為認識vvi-miR172家族及靶基因在葡萄不同器官不同組織的時空表達特征,本研究基于葡萄54個器官組織的轉錄組數據[27]和70個miRNA測序數據[28]對vvi-miR172s及靶基因的表達模式進行分析,發現vvi-miR172c及靶基因在13個不同組織或器官中均有表達。vvi-miR172c在花器官(心皮、花粉、雄蕊和花序)中的表達水平相對較高,在運輸器官(穗軸和莖)和生殖器官(果實和種子)的表達相對較低;相比之下,靶基因在所有組織中具有較高的表達水平,和在不同組織中的表達水平較低(圖8-A)。進一步分析vvi-miR172c及其靶基因在花、果實、莖、葉基本組織中的表達,發現vvi-miR172c在莖、葉和果實中低表達,而靶基因和呈現相反的表達模式(圖8-B),表明vvi-miR172家族可能主要通過vvi-miR172c介導靶基因和調控葡萄莖、葉、果實的發育過程。
2.4.3 vvi-miR172s及靶基因在果實不同組織中的時空表達圖譜分析 vvi-miR172a/b/d在其發育前期表達量高,在發育后期表達量低(圖9-A),其中,vvi-miR172a和vvi-miR172d呈現出相似的表達模式,在幼果期(10 DAF)表達量最高,在近成熟期(85 DAF)表達量最低(圖9-A),而vvi-miR172b在硬核期(35 DAF)果皮中有最高的表達,表明前兩個成員可能與幼果期果皮發育相關,而后者可能與硬核期果皮發育相關性更強。而vvi-miR172c在果皮發育的各個階段都表現出較低的表達水平(圖9-A)。相比之下,靶基因在果皮發育中呈現幼果期較高表達,而近成熟期最高表達的趨勢,與vvi-miR172a/b/d的表達趨勢相反;與vvi-miR172c呈相反的表達趨勢,從幼果果皮到成熟期逐漸降低(圖9-A)。這些結果表明,vvi-miR172s家族的不同成員可能分別參與葡萄果皮發育不同時期的調控

A:靶基因的系統進化;B:靶基因基因結構;C:靶基因保守基序

A:靶蛋白的二級結構預測;B:靶蛋白的三級結構預測;C:靶蛋白保守結構域分析;D:靶蛋白的亞細胞定位預測

A:vvi-miR172s及其靶基因啟動子不同類型順式作用元件匯總;B:vvi-miR172s及其靶基因激素相關順式作用元件
如圖9-B所示,與果皮中不同,果肉中vvi- miR172a/b具有相同的表達模式,均在果實第二次膨大期(60 DAF)表達水平最高,其他時期具有較低的表達;vvi-miR172d隨著果肉發育逐漸降低,在幼果果肉中有最高表達,成熟期表達最低,而vvi- miR172c在整個果肉發育期的表達均較低。對比相關靶基因發現,在果肉不同發育時期均呈現較高表達,與vvi-miR172c呈相反表達趨勢,而在果肉發育過程中表達逐漸升高,相反于vvi-miR172d的表達趨勢,表明vvi-miR172c/d可能負調控其靶基因介導葡萄果肉發育,而vvi-miR172a/b可能與葡萄果實膨大發育相關。
對比vvi-miR172s和靶基因的表達模式發現,葡萄果皮中,vvi-miR172a/b/d與靶基因呈較顯著的負相關,與其他靶基因的相關性較差;而在葡萄果肉中,vvi-miR172d顯著負調控的表達(圖9-B),表明vvi-miR172s及其靶基因的表達存在時空特異性。且在果實不同組織和不同發育時期,vvi-miR172家族成員對靶基因的調控作用存在差異,果皮組織中vvi-miR172家族可能主要通過vvi-miR172a/b/d介導起調控作用,而果肉中vvi-miR172d靶向的調控作用更強。
2.4.4 vvi-miR172s及靶基因在果實不同組織發育過程中應答GA3的模式分析 赤霉素處理可誘導葡萄產生無核果實,為認識vvi-miR172s及其靶基因在葡萄果實發育過程中應答GA3的模式,分析其在GA3處理下的表達水平。果皮中外源GA3的施用使vvi-miR172c在果實第二次膨大期(60 DAF)的表達水平顯著上調,vvi-miR172a/b/d在幼果期和硬核期的表達量顯著下降(圖10-A)。GA3處理顯著下調了靶基因的表達,但的表達在果實第二次膨大期顯著上調。
相關性分析顯示,GA3處理增強了果皮組織中vvi-miR172d對的負調控作用(圖11-A)。GA3處理使果肉中vvi-miR172d在各個發育階段呈下降趨勢,但顯著增加vvi-miR172c在果實第二次膨大期(60 DAF)的表達。靶基因在60 DAF顯著下調,的表達在GA3處理下顯著上調(圖10-B)。對比赤霉素處理后vvi-miR172s和靶基因表達水平的相關性,發現在果肉中GA3同樣增強了果實發育前期vvi-miR172d與的負相關性,同時誘導了vvi-miR172c對的負調控作用(圖11-B),表明vvi-miR172s及其靶基因可能存在不同的GA3應答模式參與葡萄果皮和果肉的發育過程,其中vvi-miR172c可能通過應答GA3介導相應靶基因參與葡萄果實第二次膨大發育的調控,而vvi-miR172d可能響應GA3介導其靶基因調控葡萄幼果期與硬核期果實的發育。

A:vvi-miR172c及其靶基因在葡萄不同器官、組織和發育階段的表達譜;B:vvi-miR172c及其靶基因在葡萄不同組織的表達水平。Stamen:雄蕊;Bud-AB:萌發后的芽;Inflorescence-WD:發育良好的花序;Flower-FB:始花期;Flower-F:盛花期;Carpel:心皮;Pollen:花粉;Rachis-PFS:坐果后的穗軸;Stem-G:綠色的莖;Stem-W:木質化的莖;Seed-G:綠果的種子

*、**分別代表差異顯著(<0.05)和極顯著(<0.01)。下同
*,** represented significant difference (<0.05) and extremely significant difference (<0.01), respectively. The same as below
圖9 vvi-miR172a/b/c/d及其靶基因在葡萄果皮(A)和果肉(B)中的表達水平
Fig. 9 Expression levels of vvi-miR172a/b/c/d and its target genes in berry pericarp (A) and flesh (B) of grape

圖10 GA3處理下vvi-miR172a/b/c/d及其靶基因在葡萄果皮(A)和果肉(B)中的表達水平

3 討論
MicroRNAs是一類內源性的非編碼小RNA,在基因表達的轉錄后水平起重要的調控作用。其中,高度保守的miR156和miR172被認為是植物營養生長階段變化的主要調控因子[8]。而miR172最早是在擬南芥中通過小RNA測序獲得,由于其具有高度保守性,目前已在許多植物中鑒定到miR172的存在。本研究發現葡萄中miR172有4個成員(miR172a/b/c/d),和水稻、油菜中的成員數量相同,擬南芥miR172有5個成員,麻瘋樹有2個成員,表明miR172家族成員在不同物種中存在很大的差異。miR172是第一個主要通過抑制翻譯來調節其靶標的植物miRNA,但在葡萄中vvi-miR172s通過裂解作用介導其靶基因的表達。miR172s的靶基因主要為AP2轉錄因子基因家族成員,AP2/ERF蛋白是一個轉錄因子大家族,在生物和非生物脅迫以及發育階段起關鍵的作用,目前已在擬南芥[29]、水稻[30]、大麥、小麥[31]、油菜[1]、楊樹[32]和花生[33]中鑒定到其存在。MiR156-miR172-AP2被認為是一種調控營養生長向生殖生長轉變的途徑,進而影響植物開花時間和花的發育[34]。AP2蛋白含有一到兩個AP2結構域,AP2域包括60—70個高度保守的氨基酸(aa)和兩個保守的YRG和RAYD motif元件,分別在AP2域的N和C末端[1]。本研究中葡萄miR172s的4條靶基因(、和),均含有兩個AP2蛋白結構域,但不同物種中靶基因存在差異,油菜和擬南芥中分別含有19和6個AP2類靶基因。靶基因的進化分析顯示,相比于擬南芥和水稻,葡萄中miR172靶基因的親緣關系較遠,推測葡萄中miR172靶基因的進化程度較大,其功能可能更加多樣化。
MicroRNA及其靶基因在不同組織中的表達具有時空特異性。Jofuku等[35]指出,與其他花同源基因不同,AP2在擬南芥的非花組織(莖和葉)和花組織(萼片、花瓣、雄蕊、心皮、發育中的胚珠和花序分生組織)中均有表達。GIL-HUMANES等[31]研究了AP2類基因在小麥和大麥不同發育階段根、莖、幼葉和穗中的表達,發現AP2類基因在所有組織中都有表達,在穗發育的早期達到高峰,并隨著穗的成熟而逐漸減少。油菜中的研究表明大部分在花器官中高表達,推測其在花發育過程中具有特殊的功能;且、和在晚花材料中的表達水平均高于早花材料,表明它們可能具有抑花作用[1]。本試驗的熒光定量分析顯示,vvi-miR172s及其靶基因在果實發育過程中的表達存在顯著差異,vvi-miR172c在果皮和果肉中的表達水平較低,在擬南芥中,miR172a/b/c的表達在生殖生長階段升高,而miR172d/e基因的表達很低,并且不會隨著植物的發育而改變[34];在水稻中,miR172c在幼苗中表達,但在發育中的谷物中不表達[36],表明在不同物種中miR172c的表達存在顯著差異。本研究發現,外源GA3增強了果皮和果肉組織中vvi-miR172d對的負調控作用,同時誘導了果肉組織中vvi-miR172c對//的負調控作用。該研究結果初步表明vvi-miR172c/d可能介導調控葡萄果實的發育,為深入研究vvi-miR172s及其靶基因響應GA信號調控葡萄果實發育的分子機制奠定了重要的基礎。
4 結論
葡萄miR172家族含有vvi-miR172a/b/c/d四個成員,均可通過裂解、四條靶基因參與調控葡萄果皮、果肉發育。其中,vvi-miR172a/b/d和vvi-miR172c可能分別介導靶基因和在果皮和果肉組織發育中起著更為重要的作用,而vvi-miR172c可能主要響應GA3信號介導調控葡萄果實膨大期果皮、果肉的發育。
[1] Wang T Y, Ping X K, Cao Y R, Jian H J, GA3o Y M, Wang J, Tan Y C, Xu X F, Lu K, Li J N, Liu L Z. Genome-wide exploration and characterization of miR172/euAP2 genes inL. for likely role in flower organ development. BMC Plant Biology, 2019, 19(1): 336.
[2] XU Z S, CHEN M, LI L C, MA Y Z. Functions and application of the AP2/ERF transcription factor family in crop improvement. Journal of Integrative Plant Biology, 2011, 53(7): 570-585.
[3] LI J, LUAN Y S, ZHAI J M, LIU P, XIA X Y. Bioinformatic analysis of functional characteristics of miR172 family in tomato. Journal of Northeast Agricultural University (English edition), 2013, 20(4): 21-29.
[4] 王夢琦, 解振強, 孫欣, 李曉鵬, 朱旭東, 王晨, 房經貴. 葡萄miR159及其靶基因VvGAMYB在花發育過程中的作用分析. 園藝學報, 2017, 44(6):1061-1072.
Wang M Q, Xie Z Q, Sun X, Li X P, Zhu X D, Wang C, Fang J G. Function analysis of miR159 and its target genein grape flower development. Acta Horticulturae Sinica, 2017, 44(6): 1061-1072. (in Chinese)
[5] MATHIEU J, YANT L J, MURDTER F, KUTTNER F, SCHMID M. Repression of flowering by the miR172 target SMZ. PLoS Biology,2009, 7(7): e1000148.
[6] ZENG C Y, WANG W Q, ZHENG Y, CHEN X, BO W P, SONG S, ZHANG W X, PENG M. Conservation and divergence of microRNAs and their functions in Euphorbiaceous plants. Nucleic Acids Research,2010, 38(3): 981-995.
[7] TANG M Y, BAI X, NIU L J, CHAI X, CHEN M S, XU Z F. miR172 regulates both vegetative and reproductive development in the perennial woody plant. Plant and Cell Physiology,2018, 59(12): 2549-2563.
[8] ZHU Q H, HELLIWELL C A. Regulation of flowering time and floral patterning by miR172.Journal of Experimental Botany,2011, 62: 487-495.
[9] 侍婷, 高志紅, 章鎮, 莊維兵. MicroRNA參與植物花發育調控的研究進展. 中國農學通報, 2010, 26(13): 267-271.
SHI T, GAO Z H, ZHANG Z, ZHUANG W B. Advance of research on microRNA in flower development regulation. Chinese Agricultural Science Bulletin,2010, 26(13): 267-271. (in Chinese)
[10] YAO JL,TOMES S,XU J,GLEAVE A P. How microRNA172 affects fruit growth in different species is dependent on fruit type. Plant Signaling andBehavior,2016,11(4): e1156833.
[11] RIPOLL J J, BAILRY L J, MAI Q A, WU S L, HON C T, CHAPMAN E J, DITTA G S, YANOFSKY M E, YANOFSKY M F. microRNA regulation of fruit growth. Nature Plants, 2015, 1(4): 15036.
[12] CHEN X M. A microRNA as a translational repressor of APETALA2 inflower development. Science,2004, 303(5666): 2022-2025.
[13] JUNG J H, SEO Y H, SEO P J, REYES J L, JU Y, CHUA N H, PARK C M. The GIGANTEA-regulated microRNA172 mediates photoperiodic flowering independent of CONSTANS in. Plant Cell,2007, 19(9): 2736-2748.
[14] JUNG J H, SEO P J, KANG S K, PARK C M. MiR172 signals are incorporated into the miR156 signaling pathway at the SPL3/4/5 genes indevelopmental transitions. Plant Molecular Biology, 2011, 76(1): 35-45.
[15] YANT L, MATHIEU J, DINH T T, OTT F, LANZ C, WOLLMANN H, CHEN X, SCHMID M. Orchestration of the floral transition and floral development inby the bifunctional transcription factor APETALA2. Plant Cell,2010, 22(7): 2156-2170.
[16] BARTLEY GE, ISHIDA BK. Digital fruit ripening: Data mining in the TIGR tomato gene index. Plant Molecular Biology Reporter, 2002, 20(2): 115-130.
[17] ALBA R, PAYTON P, FEI ZJ, MCQUINN R, DEBBIE P, MARTIN GB, TANKSLEY SD, GIOVANNONI JJ. Transcriptome and selected metabolite analyses reveal multiple points of ethylene control during tomato fruit development. Plant Cell, 2005, 17(11): 2954-2965.
[18] CHUNG MY, VREBALOV J, ALBA R, LEE J, MCQUINN R, CHUNG JD, KLEIN P, GIOVANNONI J. A tomato () APETALA2/ERF gene, SlAP2a, is a negative regulator of fruit ripening. Plant Journal,2010, 64(6): 936-947.
[19] 滕飛. 擬南芥AP2/ERF基因ERF055調控莖端分生組織發育的功能研究[D]. 泰安: 山東農業大學, 2013.
Teng F. Functional study ofAP2/ERF family gene
[20] 張計育, 王慶菊, 郭忠仁. 植物AP2/ERF類轉錄因子研究進展. 遺傳, 2012, 34(7): 835-847.
ZHANG J Y, WANG Q J, GUO Z R. Progresses on plant AP2/ERF transcription factors. Hereditas, 2012, 34(7): 835-847. (in Chinese)
[21] 王文然, 王晨, 解振強, 賈海鋒, 湯崴, 崔夢杰, 房經貴. VvmiR397a及其靶基因VvLACs在葡萄果實發育中的作用分析. 園藝學報, 2018, 45(8): 1441-1455.
WANG W R, WANG C, XIE Z Q, JIA H F, TANG W, CUI M J, FANG J G. Function analysis of VvmiR397a and its target genesin grape berry development. Acta Horticulturae Sinica,2018, 45(8): 1441-1455. (in Chinese)
[22] KUNST L, KLENZ J E, MARTINEZ-ZAPATER J, HAUGHN G W. AP2 gene determines the identity of perianth organs in flowers of. Plant Cell, 1989, 1(12): 1195-1208.
[23] OKAMURO J K, BOER BGW, JOFUKU KD. Regulation of Arabidopsis flower development.Plant Cell, 1993, 5(10): 1183-1193.
[24] NIU X, HELENTJARIS T, BATE N J. Maize ABI4 binds coupling element1 in abscisic acid and sugar response genes. Plant Cell, 2002, 14(10): 2565-2575.
[25] WANG C, ShANGGUAN L F, NICHOLAS K K, WANG X C, HAN J, SONG C N, FANG J G. Characterization of microRNAs identified in a table grapevine cultivar with validation of computationally predicated grapevine miRNAs by miR-RACE. PLoS ONE, 2016(7): e21259.
[26] 張文穎, 王晨, 朱旭東, 馬超, 王文然, 冷祥鵬, 鄭婷, 房經貴. 葡萄全基因組DELLA蛋白基因家族鑒定及其應答外源赤霉素調控葡萄果實發育的特征. 中國農業科學, 2018, 51(16): 3130-3146.
ZHANG W Y, WANG C, ZHU X D, MA C, WANG W R, LENG X P, ZHENG T, FANG J G. Genome-wide identification and expression of DELLA protein gene family during the development of grape berry induced by exogenous GA.Scientia Agricultura Sinica, 2018, 51(16): 3130-3146. (in Chinese)
[27] 朱旭東. 葡萄蔗糖合酶基因VvSS3的初步功能研究[D]. 南京: 南京農業大學, 2017.
ZHU X D. The preliminary function analysis of the sucrose synthase gene VvSS3 in[D]. Nanjing: Nanjing Agricultural University, 2017. (in Chinese)
[28] Akullan j b, pinto d l p, bertolinie, fasoli m, zenoni s, tornielli g b, pezzotti m, meyers b c, farina l, pe m e, mica e. miRVine: A microRNA expression atlas of grapevine based on small RNA sequencing. BMC Genomics, 2015, 16: 393.
[29] NAKANO T, SUZUKI K, FUJIMURA T, SHINSHI H. Genome-wide analysis of the ERFgene family inand rice. Plant Physiology, 2006, 140(2):411-432.
[30] TANG M F, LI G S, CHEN M S. The phylogeny and expression pattern of APETALA2-like genes in rice. Journal of Genetics and Genomics,2007, 34(10): 930-938.
[31] GIL-HUMANES J, PISTON F, MARTIN A, BARRO F. Comparative genomic analysis and expression of the APETALA2-like genes from barley, wheat, and barley-wheat amphiploids. BMC Plant Biology, 2009, 9(1): 66-79.
[32] ZHUANG J, CAI B, PENG R H, ZHU B, JIN X F, XUE Y, GAO F, FU X Y, TIAN Y S, ZHAO W, QIAO Y S, ZHANG Z, XIONG A S, YAO Q H. Genome-wide analysis of the AP2/ERF gene family in. Biochemical and Biophysical Research Communications, 2008, 371(3): 468-474.
[33] ZHANG G Y, CHEN M, CHEN X P, XU Z S, GUAN S, LI L C, LI A L, GUO J M, MAO L, MA Y Z. Phylogeny, gene structures, and expression patterns of the ERF gene family in soybean (L.). Journal of Experimental Botany, 2008, 59(15): 4095-4107.
[34] LI J P, CHEN F J, LI Y Q, LI P C, WANG Y Q, MI G H, YUAN L X. ZmRAP2.7, an AP2 transcription factor, is involved inroots development. Frontiers in plant science, 2019, 10: 820.
[35] JOFUKU K D, DEN BOER B D, VAN MONTAGU M, OKAMURO J K. Control offlower and seed development by the homeotic gene APETALA2. Plant Cell,1994, 6(9): 1211-1225.
[36] ZHU QH, HELLIWELLl CA. Regulation of flowering time and floral patterning by miR172. Journal of Experimental Botany, 2011, 62(2):487-495.
Function Analysis of vvi-miR172s and Their Target Genes Response to Gibberellin Regulation of Grape Berry Development
XUAN XuXian1, SHENG ZiLu1, XIE ZhenQiang2, HUANG YuQing1, Gong PeiJie1, ZHANG Chuan1, ZHENG Ting1, WANG Chen1*, FANG JingGui1
1College of Horticulture, Nanjing Agricultural University, Nanjing 210095;2Jiangsu Vocational College of Agriculture and Forestry, Jurong 212499, Jiangsu
【】 miR172 is an important regulator of plant growth and development. The purpose of this research was to clarify the roles and modes response to gibberellin (GA) of the miR172 family and its target genes in development of different tissues of grape berry. 【】The mature and precursor sequences of vvi-miR172a/b/c/d were cloned and identified by miR-RACE and PCR techniques from grapevine cv. Rosario Bianco. The target genes of vvi-miR172s were predicted by psRNA Target software, and the phylogenetic, structure analysis and subcellular localization were performed by bioinformatics tools. RLM-RACE verified the cleavage roles of four target genes by vvi-miR172s. The cis-elements analysis of their promoters was predicted by PLANTCARE software. The qRT-PCR method was used to detect the temporal and spatial expression patterns of vvi-miR172s and target genes in different tissues of grape berry induced by exogenous GA3application.【】The mature and precursor sequences of vvi-miR172a/b/c/d were cloned, and four target genes (,,and) were identified from grape genome. The cleavage sites of vvi-miR172s on target genes were detected by RLM-RACE, which proved that,,andwere the true target genes. Gene structure analysis result showed that all target genes contained 10 exons, 9 introns and 2 AP2 domains. The type and number of motif elements were similar, indicating that gene structures were highly conserved. Phylogenetic analysis showed thatandwere closer to poplar, andhad high homology with soybean. The secondary structure prediction of target proteins indicated that they existed in the form of alpha-helix and further folded into the stable tertiary structure. Subcellular localization results showed that all target proteins were mainly located in the nucleus. Both vvi-miR172c andhad the hormone related cis-elements response to GA3, suggesting that they might be involved in the regulation of grape berry development by responding to corresponding hormones. Microarray data analysis revealed that the expression patterns of vvi-miR172c and target genes were tissue or orGA3n specific. RT-qPCR analysis showed that vvi-miR172a/b/d showed a decrease expression trend in pericarp, whileexhibited a reverse expression trend to the former, indicating that vvi-miR172a/b/d neGA3tively regulatedHowever, in flesh,was contradictory to vvi-miR172d, with the increased expression during grape development, suggesting that there was a significant neGA3tive correlation between vvi-miR172d andInterestingly, GA3treatment promoted the neGA3tive regulation of vvi-miR172d onand induced the neGA3tive regulation of vvi-miR172c on//. 【】andwere the real target genes of vvi-miR172s. Among the vvi-miR172 family, vvi-miR172 a/b/d might mediateregulation of pericarp development, whereas vvi-miR172d might mediateinvolved in the development of grape flesh. vvi-miR172c/d and their target genes might participate in the regulation of grape pericarp and flesh development in response to exogenous GA.
grape; fruit development; gibberellin; miR172; target gene

10.3864/j.issn.0578-1752.2021.06.011
2020-06-01;
2020-11-19
國家重點研發計劃(2018YFD1000106)、江蘇省自然科學基金面上項目(BK20181318)、國家自然科學基金面上項目(31972373)、大學生國家創新創業計劃(S20190012)
宣旭嫻,E-mail:2019804199@njau.edu.cn。通信作者王晨,E-mail:wangchen@njau.edu.cn
(責任編輯 趙伶俐)
