兩個番茄品種葉綠體超微結構及光合生理對弱光脅迫的響應
2021-03-26 00:05:50孟憲敏季延海孫旺旺武占會儲昭勝劉明池
中國農業科學 2021年5期
孟憲敏,季延海,2,孫旺旺,武占會,2,儲昭勝,劉明池,2
兩個番茄品種葉綠體超微結構及光合生理對弱光脅迫的響應
孟憲敏1,季延海1,2,孫旺旺3,武占會1,2,儲昭勝1,劉明池1,2
1北京市農林科學院蔬菜研究中心,北京 100097;2農業農村部都市農業(華北)重點實驗室,北京 100097;3北京市農林科學院北京農業生物技術研究中心,北京 100097
【】研究弱光下兩個番茄品種葉綠體超微結構及光合生理特性,揭示不同番茄品種對弱光響應的差異,挖掘番茄耐弱光潛力。以本課題組篩選出的荷蘭耐弱光品種‘佳西娜’和國內不耐弱光品種‘京丹1號’為試材,設置正常光照(CK,光強300—350 μmol·m-2·s-1),50%正常光照(弱光,光強70—80 μmol·m-2·s-1)處理15 d,觀測植株葉綠素熒光成像,測定葉片葉綠素熒光參數和光合參數,并通過掃描電鏡和透射電鏡觀察葉片氣孔形態和葉綠體結構。與對照相比,弱光使2個番茄品種葉片的凈光合速率(n)、光化學淬滅系數(P_Lss)、葉綠素(a+b)含量(chl.(a+b))、超氧化物歧化酶(SOD)活性、氣孔密度和氣孔規則分布的空間尺度降低,還導致植株干重與壯苗指數降低,葉片光穩態非化學熒光淬滅(_Lss)、丙二醛(MDA)含量與植株株高、最大節間距提高,而‘京丹1號’的變化幅度顯著大于‘佳西娜’,且‘佳西娜’的實際光量子效率(_Lss)、實際光化學效率(即PSⅡ)及葉綠體結構基本沒有改變,其葉片n顯著高于‘京丹1號’。另外,弱光下,‘佳西娜’通過降低氣孔間L(d)值來改善氣孔的規則分布,并提高了葉綠素b(chl.b)含量,降低了chl.a/b,而‘京丹1號’chl.b含量卻有所降低,chl.a/b提高,氣孔規則分布的空間尺度與規則程度降低,葉片受弱光影響較嚴重。綜上,‘佳西娜’捕光能力較強于‘京丹1號’,并將光合色素吸收的較多光能用于光化學傳遞,減少熱耗散且提高了PSⅡ實際光化學速率及光能轉化率,以維持其光合系統的運轉,光合能力及產出比‘京丹1號’稍高,耐弱光能力強。2個番茄品種對弱光的響應差異主要表現在chl.b含量、氣孔空間分布格局、葉綠體結構、SOD活性與光合熒光特性不同,使得荷蘭耐弱光品種‘佳西娜’在弱光下能維持高效率的光合作用。
番茄;弱光;光合熒光特性;氣孔;葉綠體
0 引言
【研究意義】番茄(L.)喜溫喜光,是全世界設施栽培中的主要作物之一,冬春季廣泛栽培于中國北方的單坡日光溫室[1]。但日光溫室內由于墻體、骨架或保溫被等遮擋易形成弱光及低溫環境,成為冬春茬產量和品質降低的阻礙[2-3]。研究認為秋冬季番茄開花結果期,對自然環境光強要求約為2 160 μmol·m-2·s-1,日光溫室內植株冠層上方平均日輻射量860 μmol·m-2·s-1,光照時長也低于室外,遠不能滿足番茄正常生長需求[4]。然而荷蘭全年光照嚴重不足,尤其冬至前后最短光照時長僅為5—6 h,一天光照累積量最低為150—200 J·cm-2[5],但荷蘭番茄、黃瓜產量分別可達800和1 000 t·hm-2,是中國平均水平的3—4倍[6]。經過筆者對國內外番茄品種的多年研究和生產實踐表明,除先進的補光技術外,品種的耐弱光差異是荷蘭番茄高產的重要因素之一。因此,深入研究我國與荷蘭番茄品種在弱光下的響應差異,對指導中國日光溫室番茄品種選擇具有重要意義。【前人研究進展】研究表明,弱光環境導致植株株高增加,可能出現徒長趨勢,影響葉綠體結構發育[7-8],葉片凈光合速率n[9-12]、葉綠素含量和PSⅡ實際量子效率(PSⅡ)下降[3],光合產物合成和輸出減少[13-14],而膜脂過氧化程度較高,最終導致產量和品質降低,但因品種耐受性不同導致降低幅度有顯著差異[3]。研究表明,葉片氣體交換參數不僅與氣孔密度及大小有關,也與氣孔的空間分布格局有關[15]。Zheng等[16]發現氣孔在葉片上空間分布規則時,最小鄰域距離L(d)值較小,這一分析結果是通過Ripley’s K函數將葉片上可利用所有單點距離的二階矩陣,來探究這些點在不同尺度上二維分布格局[17]。氣孔在葉片上空間分布的規律在小尺度范圍內呈規則分布,而大尺度范圍內隨機分布。【本研究切入點】前人對弱光的研究多采用白熾燈、日光燈或農藝鈉燈等,此類光源能效大、成效低,也并不完全符合植物生長的最佳光譜成分,而日漸興起的發光二極管(LED)作為日光溫室的重要補光光源之一,正在發揮更大優勢[18-19]。目前,針對品種耐弱光差異機理的研究較少,而且對不同基因型品種間光合生理特征及氣孔、葉綠體結構的系統性研究較薄弱,也少見關于弱光下番茄葉片氣孔分布的報道。【擬解決的關鍵問題】本試驗采用LED燈作為光源進行弱光模擬試驗,通過對番茄幼苗生長、葉片光合特性、氣孔特征及分布、葉綠體超微結構等的研究,探明中國和荷蘭品種間耐弱光性差異的機理。
1 材料與方法
試驗于2018年在北京市農林科學院蔬菜研究中心人工氣候室(LPH-1-CT)進行。
1.1 試驗材料
以北京市農林科學院蔬菜研究中心選育的不耐弱光櫻桃番茄品種‘京丹1號’和荷蘭瑞克斯旺公司選育的較耐弱光溫室品種‘佳西娜’為試材。
1.2 試驗設計
采用72孔穴盤于2018年2月播種,所用基質為草炭﹕蛭石=2﹕1,于3葉一心時,挑選長勢一致的健壯番茄植株隨機分組,移至人工氣候室開始試驗。2個品種均設對照與弱光處理。以LED燈(PHILIPS)為光源,光質為紅藍光2﹕1[20],以正常光強300—350 μmol·m-2·s-1為對照,50%正常光照下光強70—80 μmol·m-2·s-1為弱光。共4個處理,各設3次重復,每個重復50株。光周期12 h,晝夜溫度25℃/15℃,空氣相對濕度60%—70%。苗期統一水肥管理,營養液采用北京市農林科學院蔬菜研究中心地下水改良番茄專用配方:5 mmol?L-1KNO3、5 mmol?L-1Ca(NO3)2·4H2O、2 mmol?L-1MgSO4、1 mmol?L-1KH2PO4、50 μmol?L-1EDTA- NaFe、50 μmol?L-1H3BO3、10 μmol?L-1MnSO4·4H2O、0.8 μmol?L-1ZnSO4·7H2O、0.4 μmol?L-1CuSO4·5H2O、0.02 μmol?L-1(NH4)6Mo7O24·4H2O)[21-22],85%磷酸調節營養液EC值(2.0±0.2)mS·cm-1,pH 6.5。為保證植株生長條件和脅迫程度的一致性,同一光強處理的架子上分別放置2個品種的番茄幼苗。處理12 h觀測熒光成像,處理15 d測定生長、光合生理指標,觀察葉片氣孔、葉綠體超微結構。各指標均重復測定3次,取平均值。
1.3 測定項目及方法
1.3.1 生長指標 每處理選取代表性植株12株,用鋼尺測定株高、最大節間距;游標卡尺測定莖粗;直尺測定葉長、葉寬,并計算葉面積;烘干稱重法測定植株干重,計算壯苗指數。
1.3.2 酶活性 參考李合生[23]的方法測定超氧化物歧化酶(SOD)活性;參照Cakmak等[24]的方法測定丙二醛(MDA)含量。
1.3.3 葉片氣體交換參數和光合色素含量 于8:30 —11:00采用LI-6400XT光合儀(美國LI-COR)測定第3功能葉片的凈光合速率(n)、氣孔導度(s)、胞間CO2濃度(i)及蒸騰速率(r)。采用6400-02B紅藍光源,對照光強800 μmol·m-2·s-1,弱光處理光強為100 μmol·m-2·s-1,CO2濃度(400±2)μmol·mol-1。將已測定光合參數的葉片摘下,參考Arnon[25]的95%乙醇浸提法測定并計算總葉綠素chl.(a+b)、葉綠素a、葉綠素b含量及葉綠素a/葉綠素b(chl.a/b)。
1.3.4 葉綠體超微結構 于上午8:30取第3功能葉片中部靠葉脈處附近2 mm×2 mm小塊,用2.5%戊二醛固定,pH 7.2磷酸緩沖液沖洗,1%鋨酸固定,乙醇-丙醇梯度脫水,Epon-812環氧樹脂包埋,LKB-5型超薄切片機切片,醋酸鈾-檸檬酸鉛雙重染色,透射電鏡選取15—20個視野并拍照。
1.3.5 氣孔特征及空間分布格局分析 取樣方式同1.3.4,經沖洗及梯度脫水,CO2臨界點干燥。把葉片粘到掃描樣品臺上,離子濺射儀噴涂鍍膜,掃描電鏡觀測15—20個視野并拍照。Auto CAD 2009測量單個氣孔的長度和寬度,計算周長(P)、面積(A)及形狀指數(S)。S是通過計算單一氣孔形狀與相同面積圓之間的偏離程度來測量其形狀的復雜程度[26]。
假定每個氣孔都是葉片表面上分布的單點,氣孔最中間位置為該單點的位置。取葉片遠軸面中部,采用透明指甲油印跡法制作臨時裝片,于LEICA CTR 4000下觀察,隨機選5個視野,各拍3張照片。各處理選5張照片,計算氣孔密度。利用Arc GIS 10.2將照片于相同的地理坐標、投影坐標系下進行數字化處理,再運用多距離空間聚類分析工具(Ripley’s K函數)對表征氣孔分布狀況的點進行空間分析[16-17],為了確定95%的可信任區間,采用蒙特卡洛算法模擬隨機分布點1 000次,分析結果用L(d)值表示。
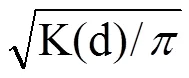
式中,L(d)表示最小鄰域距離,K(d)為任何個體在一定空間尺度內對其他個體的期望值,d為空間尺度。
1.3.6 熒光參數及成像 處理12 h,采用Fluor Cam開放式葉綠素熒光成像系統(北京Eco Tech)測定第3功能葉片葉綠素熒光參數,以顏色的形式對參數大小進行注釋,相應數據可直接從Report窗口導出。待測葉片經暗適應20 min,設定快門Shutter=1,敏感度Sensitivity=2.8,光照Act2=19,Super=20,采用protocols程序測定葉綠素熒光參數。
1.4 數據分析
用Microsoft Excel 2010及SPSS17.0軟件進行數據處理與分析。
2 結果
2.1 番茄葉片葉綠體結構對弱光的響應
圖1顯示,正常光照下,2個番茄品種葉片的葉綠體結構良好,呈長橢圓形或梭形,基粒片層垛疊緊致,基質片層結構清晰,葉綠體被膜清晰、完整,有少量嗜鋨顆粒。弱光下,‘佳西娜’的葉綠體有輕微膨脹,基粒片層垛疊整齊有序,結構清晰,只有少數葉綠體內含淀粉粒;‘京丹1號’的葉綠體趨于圓形或短粗形,發生嚴重腫脹,葉綠體膜嚴重溶解,片層結構數量減少并且變得混亂,基粒片層部分解體,模糊不清,葉綠體與細胞壁脫離而游離分布在細胞內,產生淀粉粒較多并有嗜鋨顆粒積累。
2.2 番茄葉片氣孔特征及空間分布格局對弱光的響應
弱光顯著降低了氣孔密度及寬度(表1)。‘佳西娜’和‘京丹1號’的氣孔密度分別比其對照降低47.39%、51.24%(<0.05),氣孔寬度分別比其對照降低19.77%、40.33%;弱光下,‘京丹1號’的氣孔面積、周長、長度及形狀指數最低(>0.05)(圖2、表1)。
對葉片上單個氣孔的空間分布格局分析,結果如圖3。氣孔在小尺度范圍內呈規則分布,在大尺度范圍內隨機分布。2個品種的葉片氣孔空間分布格局與單個氣孔間L(d)值對弱光響應有差異,即對照葉片的氣孔由規則分布轉變為隨機的空間尺度約為300 μm,而弱光下空間尺度減小為250 μm。弱光下‘京丹1號’葉片單個氣孔間L(d)比其對照提高5.95%,‘佳西娜’的L(d)值最小,比其對照降低34.78%,說明弱光下‘佳西娜’的氣孔在葉片上呈現更規則的空間分布格局。
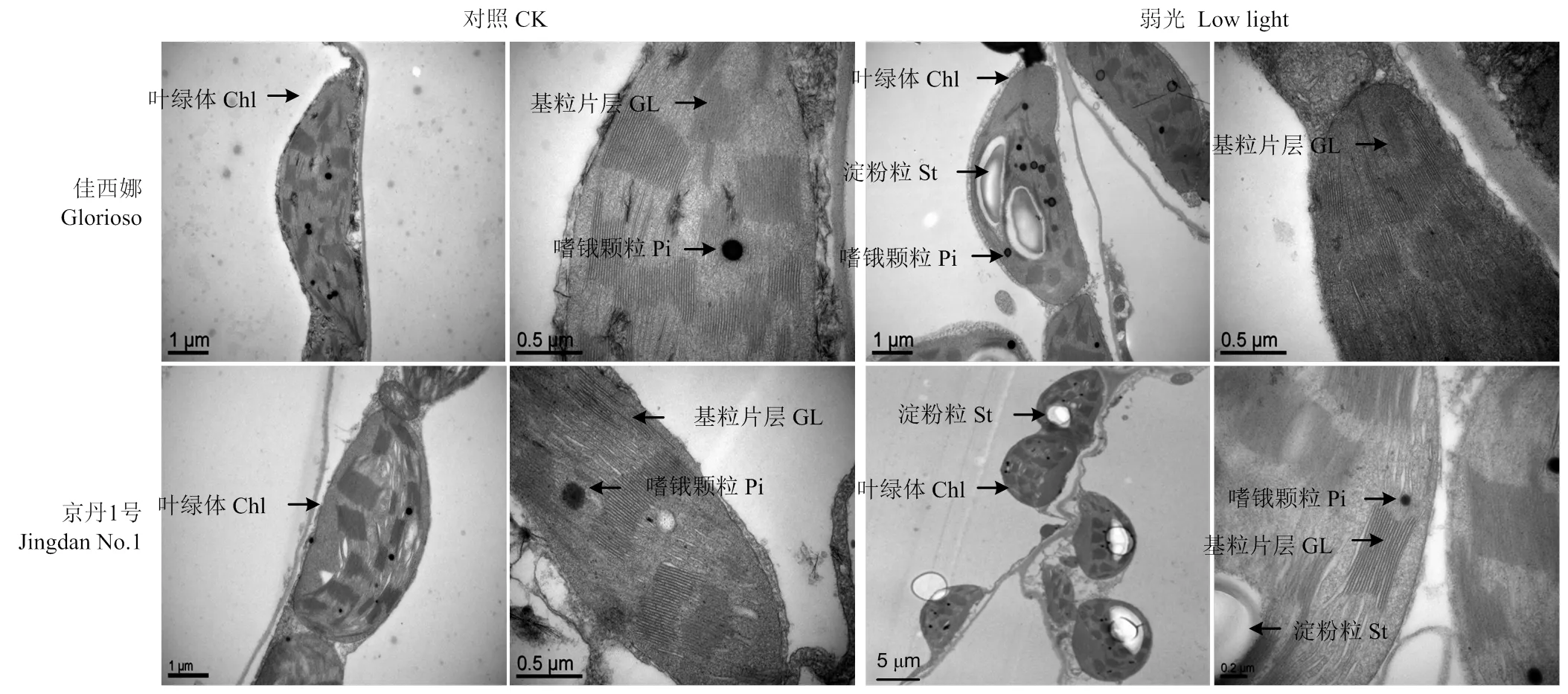
圖1 弱光對葉綠體超微結構的影響
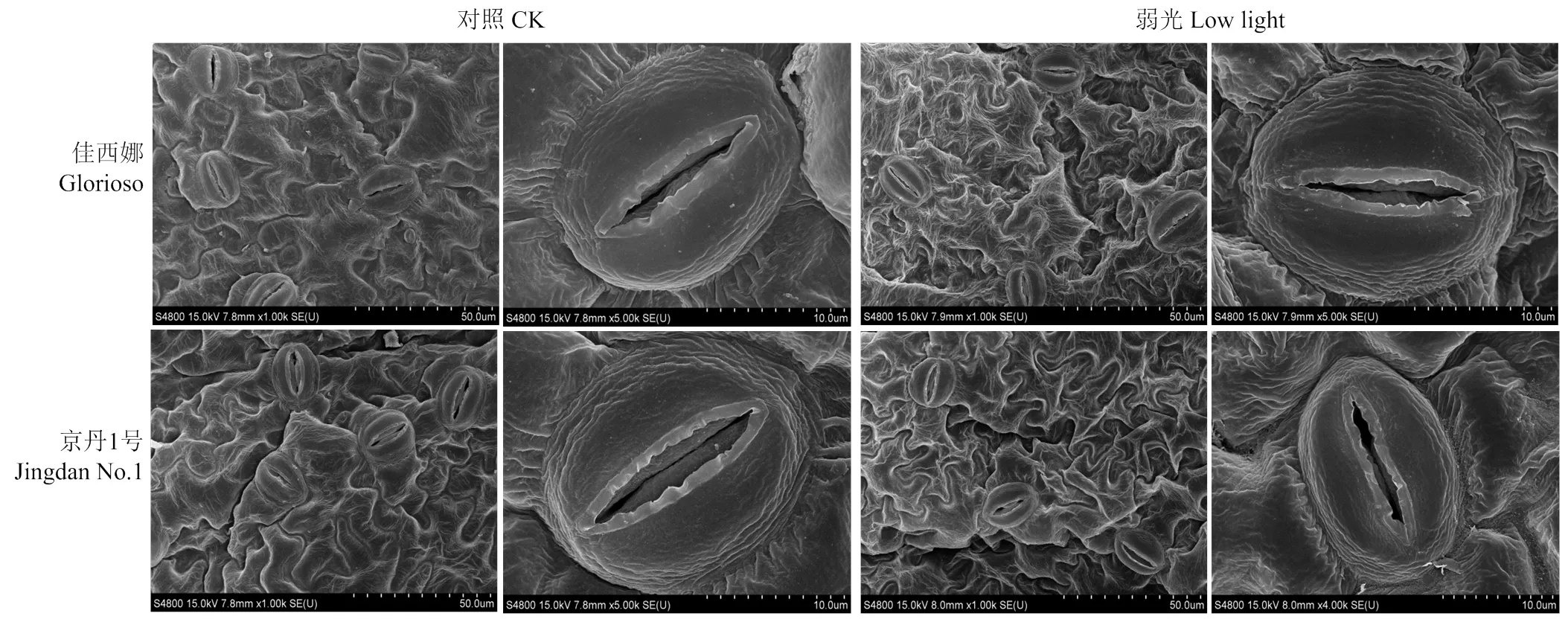
圖2 葉片單個氣孔的掃描電子顯微鏡照片
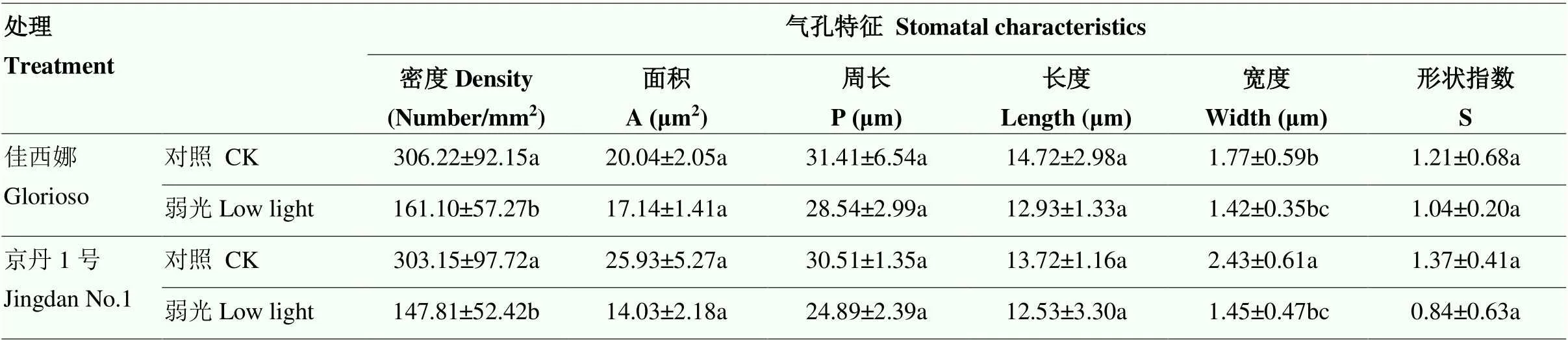
表1 弱光對葉片氣孔特征的影響
不同小寫字母表示處理間差異顯著(<0.05)。下同
Different lower-case letters indicate significant differences among treatments (<0.05). The same as below
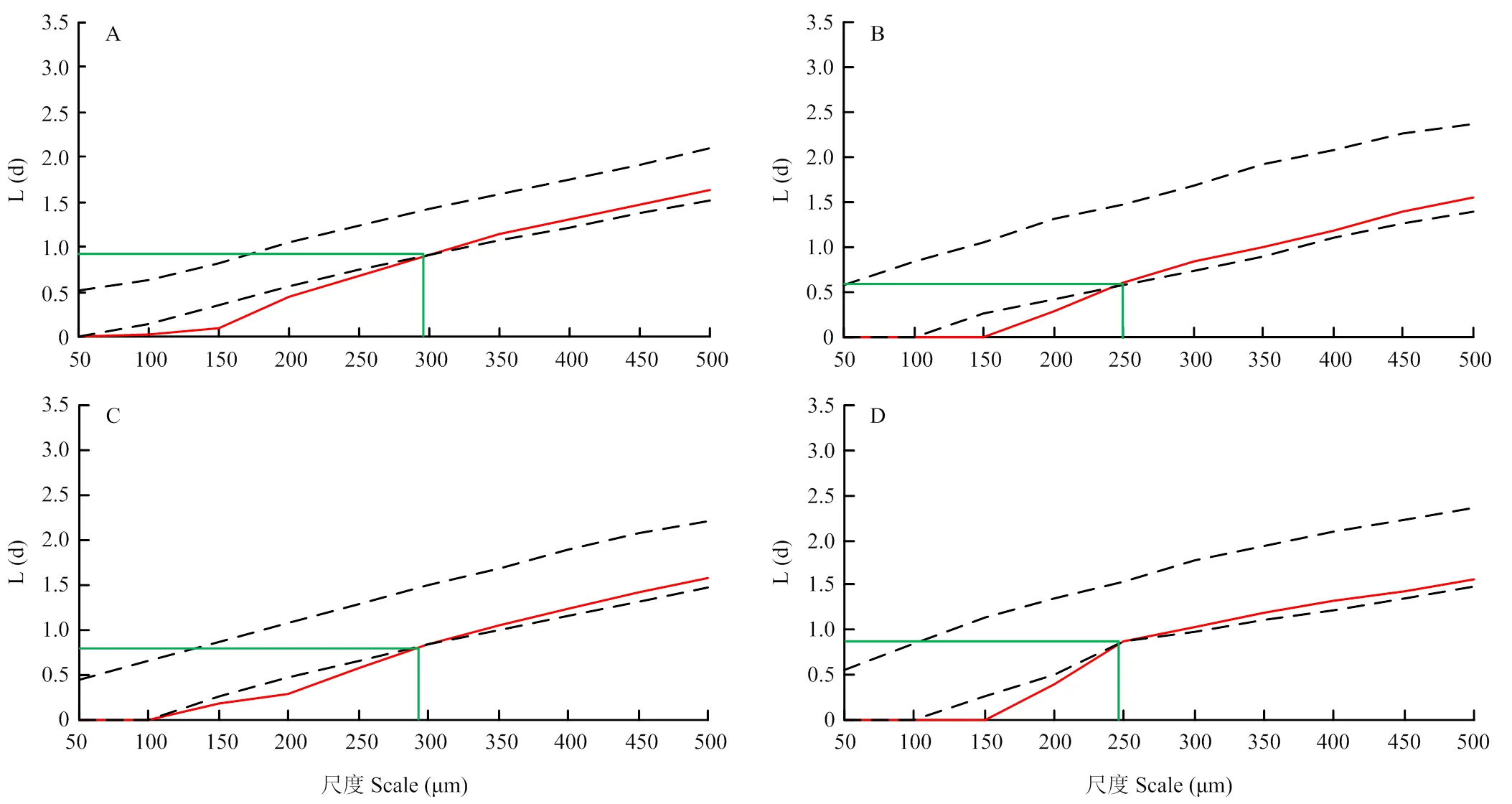
A、B分別為‘佳西娜’的對照和弱光處理,C、D分別為‘京丹1號’的對照和弱光處理。圖中上下兩條虛線代表95%置信區間的上下邊界,紅線代表K(d)值,綠線代表葉片上氣孔分布由規則變為隨機時,所對應的空間尺度和L(d)值。L(d)表示最小鄰域距離,當L(d)值小于95%置信區間時,氣孔在該尺度為規則分布,且L(d)的最小值越小,則氣孔空間分布越規則
2.3 番茄葉片SOD活性及MDA含量對弱光的響應
由圖4看出,弱光下2個番茄品種葉片SOD活性均顯著低于同期對照處理,2個品種正常光照處理下(CK)差異不顯著,弱光處理下‘佳西娜’SOD含量顯著高于‘京丹1號’。弱光下2個番茄品種MDA含量顯著上升,均顯著高于正常光照處理。弱光下,‘佳西娜’和‘京丹1號’MDA分別較正常光照下顯著提高28.88%和60.69%。
2.4 番茄葉片葉綠素含量及氣體交換參數對弱光的響應
弱光處理后,‘佳西娜’番茄葉片Chl.a、Chl.(a+b)含量和Chl.a/b均顯著低于對照植株,分別降低37.81%、27.38%和43.47%;Chl.b含量高于對照植株,但與對照差異并未達顯著性水平。‘京丹1號’的Chl.a、Chl.(a+b)含量均高于對照植株,但均不顯著。弱光處理下,‘佳西娜’的Chl.b含量比‘京丹1號’顯著增加37.50%,Chl.a/b比‘京丹1號’顯著降低25.47%(表2)。
由表3可知,弱光下,2個番茄品種的凈光合速率(n)、氣孔導度(s)、蒸騰速率(r)均低于對照。弱光處理下,‘佳西娜’的s、r與對照差異不顯著,而‘京丹1號’分別比其對照顯著降低40.00%和27.64%。弱光下,‘佳西娜’和‘京丹1號’的n分別比其對照顯著降低33.65%、60.91%。弱光下,‘佳西娜’的胞間CO2濃度(i)較其對照提高7.94%,但未達到顯著性差異,而‘京丹1號’較其對照顯著降低8.52%。

不同小寫字母表示處理間差異顯著(P<0.05)。下同
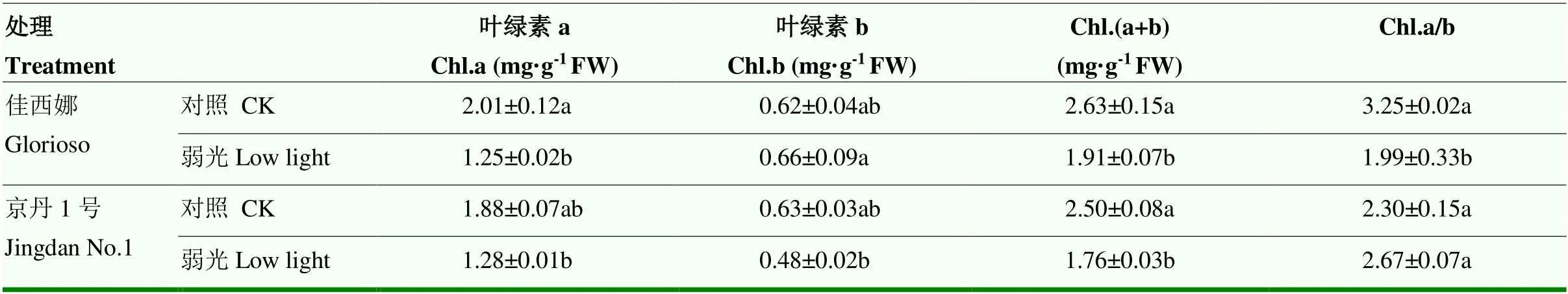
表2 弱光對葉片光合色素含量的影響
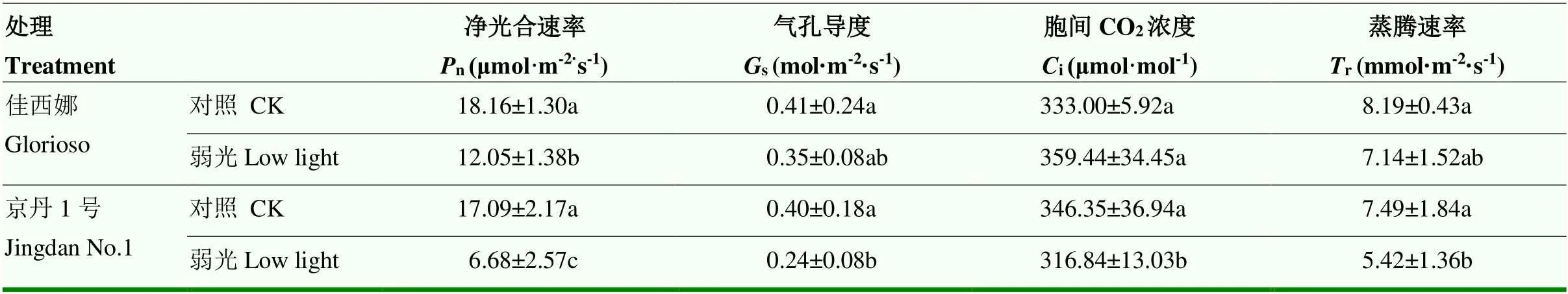
表3 弱光對葉片光合參數的影響
2.5 番茄葉片葉綠素熒光參數與熒光成像對弱光的響應
光照對暗適應過程PSⅡ最大光量子效率_max(即v/m)影響不顯著(圖5-A,B)。弱光下,相比對照植株,從光適應下實際光量子效率_Ln到光穩態下實際光量子效率_Lss,‘佳西娜’的_Ln值降低幅度逐漸減小,而‘京丹1號’的_Ln一直在其對照曲線內側,降低幅度顯著高于‘佳西娜’。弱光下,‘佳西娜’的_Lss和暗弛豫過程中PSⅡ實際光量子效率_Dn基本沒有變化,而‘京丹1號’的_Lss比其對照降低14.29%。
弱光下,2個番茄品種的升高,‘佳西娜’升高幅度小于‘京丹1號’(圖5-C,D),尤其是光適應下非光化熒光淬滅_Ln至光穩態非光化熒光淬滅_Lss。弱光下,‘京丹1號’曲線均位于雷達圖最外側,上升幅度較高;‘佳西娜’和‘京丹1號’的_Lss分別比其對照提高43.90%、55.73%,而光穩態光化學淬滅系數P_Lss分別比其對照降低9.33%、17.77%。光適應下,光化學淬滅P_Ln到暗弛豫過程中,光化學淬滅P_Dn均以‘京丹1號’降低幅度大。
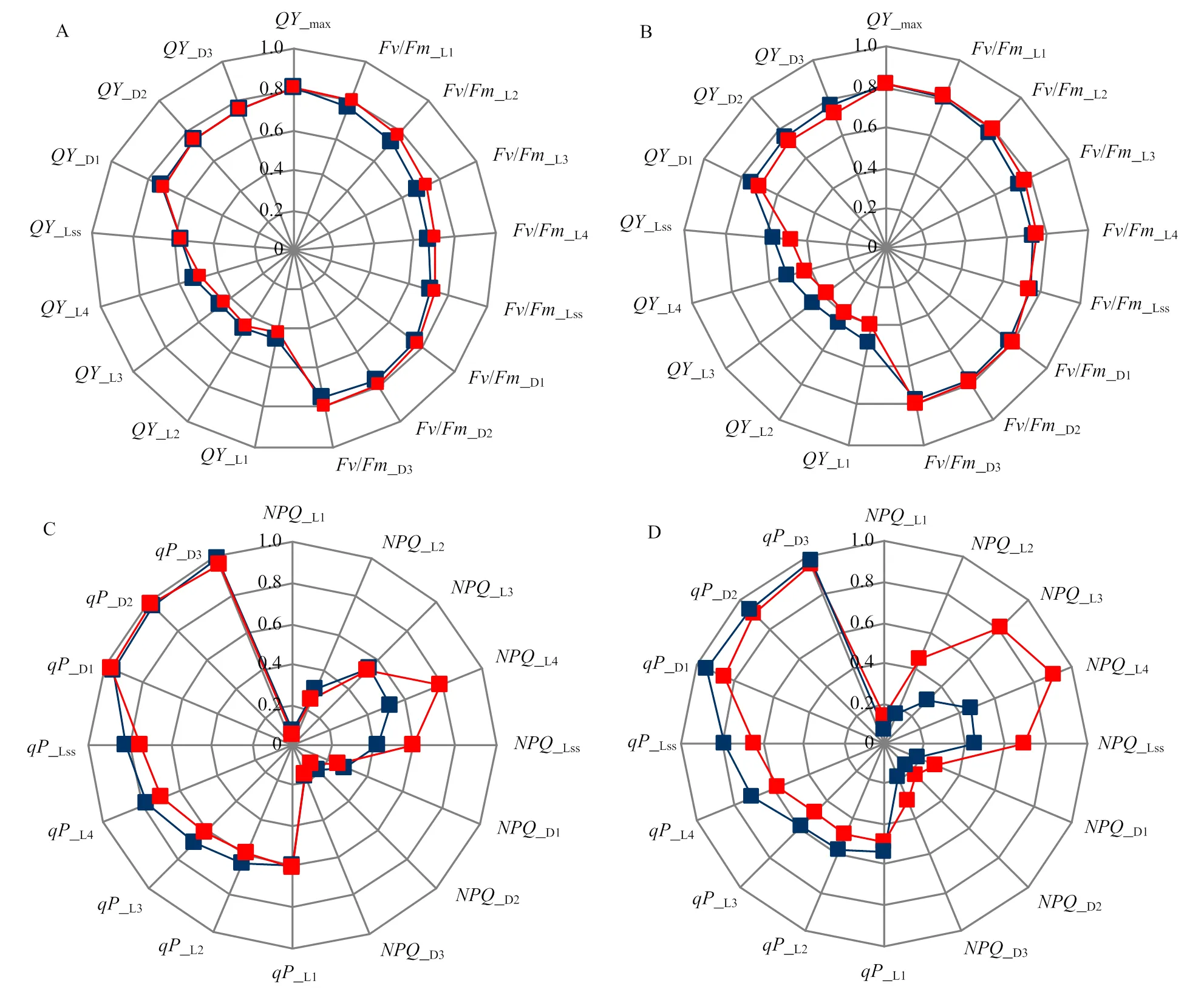
藍、紅色線分別表示對照和弱光處理。A、C為‘佳西娜’,B、D為‘京丹1號’;A、B為光量子產量參數,C、D為熒光淬滅參數
弱光處理后,番茄葉片PSⅡ最大的光量子效率v/m、實際光化學效率(即PSⅡ)與葉綠素熒光成像見圖6。v/m成像圖顯示,弱光下2個品種葉片邊緣有較少藍色或綠色,但整體與對照基本一致,證明葉片PSⅡ光系統未受到嚴重破壞;圖看出,弱光下‘京丹1號’葉片顏色變化較明顯,范圍較大,而‘佳西娜’受傷害面積較少且主要集中于葉片邊緣。圖中,弱光下番茄葉片表面出現少量橘黃色和淺綠色部分,‘京丹1號’面積大于‘佳西娜’,且集中于葉片表面和邊緣,說明弱光下‘京丹1號’葉片受影響程度較高。
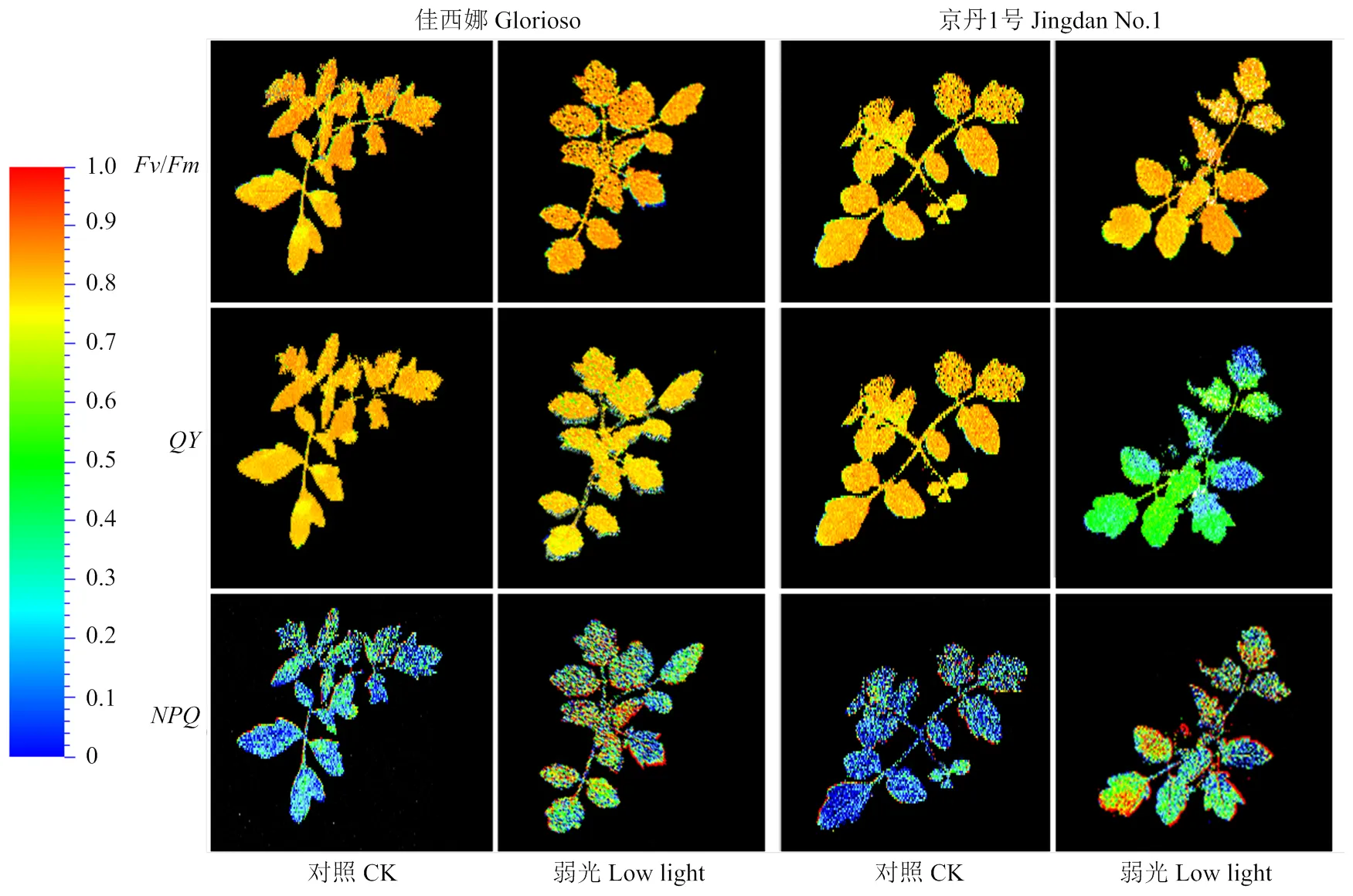
圖6 葉片葉綠素熒光成像圖
2.6 番茄幼苗生長對弱光的響應
弱光下,2個品種的株高、葉面積與最大節間距顯著提高,莖粗、壯苗指數和單株干重均降低(表4)。其中,相比各自對照處理,‘佳西娜’和‘京丹1號’的株高分別顯著提高94.82%、105.58%(<0.05),壯苗指數分別顯著降低56.25%、72.73%(<0.05),單株干重分別降低8.33%(>0.05)、45.00%(<0.05)。圖7可見,弱光下‘京丹1號’變化幅度較大,其受弱光的影響更大。
3 討論
3.1 弱光影響番茄葉片葉綠素含量與葉綠體超微結構
Ren等[27]發現弱光使玉米葉片葉綠體結構遭到破壞,葉綠素合成減少。而葉綠素b可以利用弱光下占比較高的藍紫光,提高葉片捕光能力,以維持光系統穩定[28];其含量減少會導致激發能在光系統與電子傳遞間的分配不平衡,導致PSⅠ、PSⅡ間不穩定,一般直接表現為對逆境的耐受度降低[29]。有研究表明,弱光下,不結球白菜通過增加Chl.b含量以提高光合作用[2];Meng等[30]對弱光下番茄的研究表明葉片通過降低Chl.a/b,在一定程度上減輕對PSⅠ、PSⅡ的抑制。本試驗中,弱光下2個番茄品種葉片總葉綠素含量均降低,但‘佳西娜’降低幅度小于‘京丹1號’,且‘佳西娜’的Chl.b含量較其對照提高6.48%。‘佳西娜’Chl.a/b比值降低,與ALA促進Chl.a向Chl.b轉化有關[31],這一現象是‘佳西娜’適應逆境的表現[32]。說明弱光下,荷蘭品種‘佳西娜’捕光能力強于‘京丹1號’,這是2個番茄品種弱光耐受性差異的原因之一。

表4 弱光對番茄生長指標的影響
研究表明弱光會導致葉綠體結構發生不同程度的變化[10,29]。秦玉芝等[14]認為隨著馬鈴薯對弱光環境的適應,其光合產物運輸逐漸協調,葉綠體內淀粉粒積累減少。也有研究表明,相對于中度弱光脅迫,嚴重弱光會導致花生葉片葉綠體結構發生較大變化,淀粉粒數增多[33]。本試驗中,弱光下的‘佳西娜’葉綠體基粒片層增多,垛疊整齊有序,結構清晰,少數葉綠體內含淀粉粒;而‘京丹1號’的基粒片層部分解體、模糊不清,大量淀粉粒積累。推測‘京丹1號’因n下降較多影響其無機磷供應,蔗糖在細胞質內外運輸受阻,無機磷釋放也隨之減少,磷酸丙糖將主要用于葉綠體內儲存淀粉。大量淀粉粒積累會對類囊體造成一定的機械損傷,并吸附部分碳同化相關酶,降低其酶活性[34]。意味著弱光下荷蘭品種‘佳西娜’的光能利用率與環境適應性高于‘京丹1號’。
3.2 弱光影響番茄葉片氣孔空間分布與光合氣體交換參數
氣孔是植株與外界環境進行H2O和CO2交換的重要介質,當植株遭受非生物脅迫后,葉片會通過調整氣孔密度、開度或形狀來緩解傷害[17,35]。本試驗發現,弱光下2個番茄品種葉片的氣孔密度及寬度均顯著減小,‘佳西娜’比‘京丹1號’下降幅度少,可見‘佳西娜’自我調節能力較強。另外,弱光使2個番茄品種葉片氣孔規則分布空間的尺度減小,‘佳西娜’的單個氣孔間最小鄰域距離L(d)略有降低,但‘京丹1號’的L(d)較其對照提高,說明弱光下‘佳西娜’的氣孔空間分布比‘京丹1號’更規則;且n、s降低的同時i升高,CO2氣體更易通過氣孔擴散進入組織內部,影響光合作用的最大反應速率[36-37]。這是荷蘭品種‘佳西娜’比‘京丹1號’耐弱光能力強的一個主要原因。
3.3 弱光影響番茄葉片葉綠素熒光參數與熒光成像
_max(即vm)反映了暗適應下PSⅡ反應中心光能轉化效率,可作為判斷植物對逆境耐受程度的指標[38]。(即PSⅡ)為PSⅡ反應中心電荷分離實際量子效率,反映PSⅡ吸收的光能不用于光化學反應,而以熱能耗散掉的部分[39]。本研究發現光照對_max影響不大,與熒光顯示結果一致,說明葉片PSⅡ沒有受到嚴重破壞。弱光下2個番茄品種的_Ln、P_Ls、P_Ln與P_Dn下降,_Lss、_Ln和_Dn升高,表明弱光下植株通過熱耗散消耗光合色素吸收的光能增多[40],光合中心有所損傷[41],可能會對RuBP再生產生抑制,但‘佳西娜’變化幅度顯著低于‘京丹1號’。因此,弱光下相比‘京丹1號’,‘佳西娜’的天線色素吸收的激發能用于光化學傳遞的份額較多,熱耗散比例少,反應中心開放比例和實際光化學速率較高,與其Chl.b含量高于‘京丹1號’的結果一致。通過葉綠素熒光成像圖,發現‘佳西娜’葉片光合系統遭受損害主要位于葉片邊緣,‘京丹1號’則于葉片表面大面積受影響,因此‘京丹1號’光合活性降低更多。這是荷蘭品種‘佳西娜’對弱光抵御能力與適應能力較強的重要原因之一。
3.4 弱光影響番茄幼苗生長狀況與SOD活性、MDA含量
植物處于逆境下,體內ROS增多,植物膜透性增大,膜脂肪酸中的不飽和鍵形成MDA[11];SOD可將ROS轉化成H2O2,通過酶促系統減輕膜傷害,以保護植株[42]。本試驗中,弱光下‘佳西娜’葉片的SOD活性降低較少,MDA含量低,膜脂過氧化程度較輕,而‘京丹1號’葉片過氧化程度嚴重。說明‘佳西娜’以提高SOD活性,來減輕葉綠體膜過氧化程度,提高逆境適應能力,這使荷蘭品種‘佳西娜’對弱光耐受性強于‘京丹1號’。程亞嬌等[43]認為弱光下,植株通過抑制莖橫向生長,增加株高并將更多的光合同化產物轉移至莖桿,以保證其營養供應,導致植株徒長。本試驗中,弱光下2個番茄品種株也出現徒長現象,‘京丹1號’光合產出能力也低于‘佳西娜’。
4 結論
弱光下,‘佳西娜’植株變化幅度顯著小于‘京丹1號’,其適應弱光環境的能力較強。主要表現在‘佳西娜’葉片Chl.b含量、氣孔空間規則分布程度、_Lss、P_Lss與SOD活性高于‘京丹1號’,而_Lss、葉片受傷害面積及程度低于‘京丹1號’。因此,‘佳西娜’葉片的捕光能力較高,用于光化學傳遞的份額多,提高了實際光化學效率,保證了光合產物的輸出平衡;且‘佳西娜’的葉綠體基粒片層垛疊整齊,光能轉化效率較高,植株干重顯著高于‘京丹1號’。
[1] SHU S, TANG Y Y, YUAN Y H, SUN J, ZHONG M, GUO S R. The role of 24-epibrassinolide in the regulation of photosynthetic characteristics and nitrogen metabolism of tomato seedlings under a combined low temperature and weak light stress. Plant Physiology and Biochemistry, 2016, 107: 344-353.
[2] ZHU H F, LI X F, ZHAI W, LIU Y, GAO Q Q, LIU J P, REN L, CHEN H Y, Zhu Y Y. Effects of low light on photosynthetic properties, antioxidant enzyme activity, and anthocyanin accumulation in purple pak-choi (ssp.Makino). PLoS ONE, 2017, 12(6), e0179305.
[3] 孫建磊, 王崇啟, 肖守華, 高超, 李利斌, 曹齊衛, 王曉, 董玉梅, 焦自高. 弱光對黃瓜幼苗光合特性及Rubisco酶的影響. 核農學報, 2017, 31(6): 1200-1209.
SUN J L, WANG C Q, XIAO S H, GAO C, LI L B, CAO Q W, WANG X, DONG Y M, JIAO Z G. Effect of low light on photosynthesis and Rubisco of cucumber seedlings. Journal of Nuclear Agricultural Sciences, 2017, 31(6): 1200-1209. (in Chinese)
[4] 閆文凱. 日光溫室人工補光對番茄光合作用及生長的影響[D]. 北京: 中國農業科學院, 2018.
YAN W K. Effects of artificial lighting on photosynthesis and growth of tomato in Chinese solar greenhouse [D]. Beijing: Chinese Academy of Agricultural Sciences, 2018. (in Chinese)
[5] 劉銘, 張英杰, 呂英民. 荷蘭設施園藝的發展現狀. 農業工程技術(溫室園藝), 2010(8): 24-33.
LIU M, ZHANG Y J, Lü Y M. The development and present situation of Dutch horticulture.Agriculture Engineering Technology (Greenhouse & Horticulture), 2010(8): 24-33. (in Chinese)
[6] 蔣衛杰, 鄧杰, 余宏軍. 設施園藝發展概況、存在問題與產業發展建議. 中國農業科學, 2015, 48(17): 3515-3523.
JIANG W J, DENG J, YU H J. Development situation, problems and suggestions on industrial development of protected horticulture. Scientia Agricultura Sinica, 2015, 48(17): 3515-3523. (in Chinese)
[7] Aliferis K A, Chrysayi-Tokousbalides M, Fasseas C. Physiological and ultrastructural changes in “green islands” onleaves caused by (8, 16) - (–) -pyrenophorin. Plant Physiology and Biochemistry, 2006, 44(11/12): 851-856.
[8] 艾希珍, 郭延奎, 馬興莊, 邢禹賢. 弱光條件下日光溫室黃瓜需光特性及葉綠體超微結構. 中國農業科學, 2004, 37(2): 268-273.
AI X Z, GUO Y K, MA X Z, XING Y X. Photosynthetic Characteristics and ultrastructure of chloroplast of cucumber under low light intensity in solar greenhouse. Scientia Agricultura Sinica, 2004, 37(2): 268-273. (in Chinese)
[9] Murchie E H, Hubbart S, Peng S, Horton P. Acclimation of photosynthesis to high irradiance in rice: gene expression and interactions with leaf development. Journal of Experimental Botany, 2005, 56(411): 449-460.
[10] 王學文, 王玉玨, 付秋實, 趙冰, 郭仰東. 弱光逆境對番茄幼苗形態、生理特征及葉片超微結構的影響. 華北農學報, 2009, 24(5): 144-149.
WANG X W, WANG Y J, FU Q S, ZHAO B, GUO Y D. Effects of low light stress on morphological trait, physiological characters and leaf ultrastructure of tomato (L.) seedlings. Acta Agriculturae Boreali-Sinica, 2009, 24(5): 144-149. (in Chinese)
[11] Kanazawa S, Sano S, Koshiba T, Ushimaru T. Changes in antioxidative enzymes in cucumber cotyledons during natural senescence: comparison with those during dark-induced senescence. Physiologia Plantarum, 2010, 109(2): 211-216.
[12] 李翔, 桑勤勤, 束勝, 孫錦, 郭世榮. 外源油菜素內酯對弱光下番茄幼苗光合碳同化關鍵酶及其基因的影響. 園藝學報, 2016, 43(10): 2012-2020.
LI X, SANG Q Q, SHU S, SUN J, GUO S R. Effects of epibrassinolide on the activities and gene expression of photosynthetic enzymes in tomato seedlings under low light. Acta Horticulturae Sinica, 2016, 43(10): 2012-2020. (in Chinese)
[13] 楊柳燕, 陳菁菁, 陳年來. 甜瓜葉片光合產物輸出能力對弱光的響應. 中國農業科學, 2018, 51(13): 2561-2569.
YANG L Y, CHEN J J, CHEN N L. Responses of leaf assimilate export to lowlight stress in melon. Scientia Agricultura Sinica, 2018, 51(13): 2561-2569. (in Chinese)
[14] 秦玉芝, 邢錚, 鄒劍鋒, 何長征, 李炎林, 熊興耀. 持續弱光脅迫對馬鈴薯苗期生長和光合特性的影響. 中國農業科學, 2014, 47(3): 537-545.
QIN Y Z, XING Z, ZOU J F, HE C Z, LI Y L, XIONG X Y. Effects of sustained weak light on seedling growth and photosynthetic characteristics of potato seedlings. Scientia Agricultura Sinica, 2014, 47(3): 537-545. (in Chinese)
[15] Apple M E, Olszyk D M, Ormrod D P, Lewis J, Southworth D, Tingey D T. Morphology and stomatal function of douglas fir needles exposed to climate change: Elevated CO2and temperature. International Journal of Plant Sciences, 2000, 161(1): 127-132.
[16] Zheng Y P, Xu M, Hou R X, Shen R C, Qiu S, Ouyang Z. Effects of experimental warming on stomatal traits in leaves of maize (L.).Ecology and Evolution, 2013, 3(9): 3095-3111.
[17] 郭麗麗, 郝立華, 賈慧慧, 李菲, 張茜茜, 曹旭, 徐明, 鄭云普. NaCl脅迫對兩種番茄氣孔特征、氣體交換參數和生物量的影響. 應用生態學報, 2018, 29(12): 3949-3958.
GUO L L, HAO L H, JIA H H, LI F, ZHANG Q Q, CAO X, XU M, ZHENG Y P. Effects of NaCl stress on stomatal traits, leaf gas exchange parameters, and biomass of two tomato cultivars. Chinese Journal of Applied Ecology, 2018, 29(12): 3949-3958. (in Chinese)
[18] Wang Y W, Zhang B, Kai C. Design and development of intelligent LED plant light supplement system based on solar - powered for facility agriculture. Applied Mechanics and Materials, 2014(672/674): 26-29.
[19] Davis P A, Burns C. Photobiology in protected horticulture. Food and Energy Security, 2016, 5(4): 223-238.
[20] LIU X Y, GUO S R, XU Z G, JIAO X L, Takafumi T. Regulation of chloroplast ultrastructure, cross-section anatomy of leaves and morphology of stomata of cherry tomato by different light irradiations of LEDs. Hortscience, 2011, 46(2): 217-221.
[21] 劉增鑫. 特種蔬菜無土栽培. 北京: 中國農業出版社. 2000.
Liu Z X. Soilless Cultivation of Special Vegetables. Beijing: China Agriculture Press, 2000. (in Chinese)
[22] 郭世榮. 無土栽培學. 北京: 中國農業出版社. 2011.
Guo S R. Soilless Culture. Beijing: China Agriculture Press. 2011. (in Chinese)
[23] 李合生. 植物生理生化實驗原理和技術. 北京: 高等教育出版社, 1998.
LI H S. Principle and Technology of Plant Physiology and Biochemistry Experiment. Beijing: Higher Education Press, 1998. (in Chinese)
[24] Cakmak I, Marschner H. Magnesium deficiency and high light intensity enhance activities of superoxide dismutase, ascorbate peroxidase, and glutathione reductase in bean leaves. Plant Physiology, 1992, 98: 1222-1227.
[25] Arnon D L. Copper enzymes in isolated chloroplasts, polyphenol oxidase in.Plant Physiology, 1949, 24(1): 1-15.
[26] 黃磊, 孫耀清, 郝立華, 黨承華, 朱玉, 王賀新, 程東娟, 張運鑫, 鄭云普. 高溫對北高叢越橘葉片結構和生理代謝的影響. 園藝學報, 2016, 43(6): 1044-1056.
HUANG L, SUN Y Q, HAO L H, DANG C H, ZHU Y, WANG H X, CHENG D J, ZHANG Y X, ZHENG Y P. Effects of high temperatures on leaf structures and physiological metabolism of north highbush blueberry. Acta Horticulturae Sinica, 2016, 43(6): 1044-1056. (in Chinese)
[27] Ren B Z, Zhang J W, Dong S T, Liu P, Zhao B. Effects of waterlogging on leaf mesophyll cell ultrastructure and photosynthetic characteristics of summer maize. PLoS ONE, 2016, 11(9): e0161424.
[28] Cornah J E, TERRY M J, SMITH A G. Green or red: What stops the traffic in the tetrapyrrole pathway? Trends in Plant Science, 2003, 8(5): 224-230.
[29] Chu H A, Nguyen A P, Debus R J. Site-directed photosystem II mutants with perturbed oxygen-evolving properties. 1. instability or inefficient assembly of the manganese cluster. Biochemistry, 1994, 33(20): 6150-6157.
[30] Meng Z J, Lu T, Zhang G X, Qi M F, Tang W, Li L L, Liu Y F, Li T L. Photosystem inhibition and protection in tomato leaves under low light. Scientia Horticulturae, 2017, 217: 145-155.
[31] 安玉艷, 張麗穎, 馮新新, 田凡, 李潔, 汪良駒. 5-氨基乙酰丙酸對蘋果葉片耐弱光能力的影響. 西北植物學報, 2016, 36(5): 987-995.
AN Y Y, ZHANG L Y, FENG X X, TIAN F, LI J, WANG L J. Effects of 5-aminolevulinic acid on low light tolerance of apple leaves. Acta Botanica Boreali-Occidentalia Sinica, 2016, 36(5): 987-995. (in Chinese)
[32] Rüdiger W. Biosynthesis of chlorophyll, and the chlorophyll cycle. Photosynthesis Research, 2002, 74(2): 187-193.
[33] 吳正鋒, 孫學武, 王才斌, 鄭亞萍, 萬書波, 劉俊華, 鄭永美, 吳菊香, 馮昊, 于天一. 弱光脅迫對花生功能葉片RuBP羧化酶活性及葉綠體超微結構的影響. 植物生態學報, 2014, 38(7): 740-748.
WU Z F, SUN X W, WANG C B, ZHENG Y P, WAN S B, LIU J H, ZHENG Y M, WU J X, FENG H, YU T Y. Effects of low light stress on rubisco activity and the ultrastructure of chloroplast in functional leaves of peanut. Acta Phytoecologica Sinica, 2014, 38(7): 740-748. (in Chinese)
[34] 姚允聰, 王紹輝, 孔云. 弱光條件下桃葉片結構及光合特性與葉綠體超微結構變化. 中國農業科學, 2007, 40(4): 855-863.
YAO Y C, WANG S H, KONG Y. Characteristics of photosynthesis machinism in different peach species under low light intensity. Scientia Agricultura Sinica, 2007, 40(4): 855-863. (in Chinese)
[35] Casson S, Gray J E. Influence of environmental factors on stomatal development. New Phytologist, 2008, 178(1): 9-23.
[36] 朱玉, 黃磊, 黨承華, 王賀新, 姜國斌, 李根柱, 張自川, 婁鑫, 鄭云普. 高溫對藍莓葉片氣孔特征和氣體交換參數的影響. 農業工程學報, 2016, 32(1): 218-225.
ZHU Y, HUANG L, DANG C H, WANG H X, JIANG G B, LI G Z, ZHANG Z C, LOU X, ZHENG Y P. Effects of high temperature on leaf stomatal traits and gas exchange parameters of blueberry. Transactions of the Chinese Society of Agricultural Engineering, 2016, 32(1): 218-225. (in Chinese)
[37] Farquhar G D, Sharkey T D. Stomatal conductance and photosynthesis. Annual Review of Plant Physiology, 1982, 33: 317-345.
[38] 張守仁. 葉綠素熒光動力學參數的意義及討論. 植物學通報, 1999, 16(4): 444-448.
ZHANG S R. A discussion on chlorophyll fluorescence kinetics parameters and their significance. Chinese Bulletin of Botany, 1999, 16(4): 444-448. (in Chinese)
[39] Genty B, Briantais J M, Baker N R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence.Biochimica et Biophysica Acta (BBA)-General Subjects, 1989, 990(1): 87-92.
[40] 艾希珍, 王秀峰, 崔志峰, 王振林. 鈣對弱光亞適溫下黃瓜光合作用的影響. 中國農業科學, 2006, 39(9): 1865-1871.
AI X Z, WANG X F, CUI Z F, WANG Z L. Effect of calcium on photosynthesis of cucumber under low light intensity and sub-optimal temperature. Scientia Agricultura Sinica, 2006, 39(9): 1865-1871. (in Chinese)
[41] 孫德智, 韓曉日, 彭靖, 范富, 張慶國. 外源NO對Ca(NO3)2脅迫下番茄幼苗PSII功能及光能分配利用的影響. 核農學報, 2016, 30(12): 2451-2459.
SUN D Z, HAN X R, PENG J, FAN F, ZHANG Q G. Effect of exogenous nitric oxide on PSII function and distribution and utilization of luminous energy in tomato seedlings under stress of Ca(NO3)2. Journal of Nuclear Agricultural Sciences, 2016, 30(12): 2451-2459. (in Chinese)
[42] Kornyeyev D, Logan B A, Payton P, Allen R D, Holaday A S. Enhanced photochemical light utilization and decreased chilling-induced photoinhibition of photosystem II in cotton overexpressing genes encoding chloroplast-targeted antioxidant enzymes. Physiologia Plantarum, 2001, 113(3): 323-331.
[43] 程亞嬌, 諶俊旭, 王仲林, 范元芳, 陳思宇, 李澤林, 劉沁林, 李中川, 楊峰, 楊文鈺. 光強和光質對大豆幼苗形態及光合特性的影響. 中國農業科學, 2018, 51(14): 2655-2663.
CHENG Y J, SHEN J X, WANG Z L, FAN Y F, CHEN S Y, LI Z L, LIU Q L, LI Z C, YANG F, YANG W Y. Effects of light intensity and light quality on morphological and photosynthetic characteristics of soybean seedlings. Scientia Agricultura Sinica, 2018, 51(14): 2655-2663. (in Chinese)
Response of Chloroplast Ultrastructure and Photosynthetic Physiology of Two Tomato Varieties to Low Light Stress
MENG XianMin1, JI YanHai1,2, SUN WangWang3, WU ZhanHui1,2, CHU ZhaoSheng1, LIU MingChi1,2
1Vegetable Research Center, Beijing Academy of Agriculture and Forestry Sciences, Beijing 100097;2Key Laboratory of North China Urban Agriculture, Ministry of Agriculture and Rural Affairs, Beijing 100097;3Beijing Agro-Biotechnology Research Center, Beijing Academy of Agriculture and Forestry Sciences, Beijing 100097
【】 The chloroplast ultrastructure and photosynthetic physiological characteristics of two tomato cultivars under low light were studied to reveal the difference of the response of different tomato cultivars to low light and explore the potential oftolerance to low light. 【】The Dutch low-light tolerance cultivar Glorioso and the Chinese low-light sensitive cultivar Jingdan No. 1 were selected as experimental materials, they exposed to normal light (CK, light intensity 300-350 μmol·m-2·s-1) and 50% normal light (low light, light intensity 70-80 μmol·m-2·s-1) for fifteen days. The plant chlorophyll fluorescence imaging, chlorophyll fluorescence and photosynthetic parameters of leaves were detected. We also observed stomatal morphology and chloroplast ultrastructure of leaves after treatment by the scanning electron microscopy (SEM) and transmission electron microscopy (TEM). 【】 Compared with the control, then,P_Lss, the content of chl. (a+b), SOD activity, stomatal density and spatial scale of regular stomatal distribution of the two cultivars leaves were decreased, which also led to the decrease of dry weight and healthy index, the leaves_Lss, MDA content increased with plant height and maximum internode spacing. The variation range of Jingdan No. 1 was significantly larger than Glorioso, the_Lss,(PSⅡ) and chloroplast structure of Glorioso remained unchanged, itsnin leaves was significantly higher than Jingdan No. 1. In addition, the Glorioso improved the regular distribution of stomata by reducing the value of L(d) between stomata, chl.b content increased and chl.a/b decreased under low light. While the chl.b content of Jingdan No. 1 decreased, chl.a/b increased, the spatial scale and regularity of stomatal regular distribution reduced and the leaves were seriously affected by low light. To sum up, Glorioso has stronger light-harvesting ability than Jingdan No. 1 under low light, and uses more light energy absorbed by photosynthetic pigments for photochemistry transfer, reduced heat dissipation and improves the actual photochemistry rate, light energy conversion rate of PSII, so as to maintain operation of photosynthetic system under low light, which photosynthetic capacity and output are slightly higher than Jingdan No. 1, and has strong low light tolerance.【】 The differences of response to low light between the two tomato cultivars were mainly reflected in chl.b content, stomatal spatial distribution pattern, chloroplast structure, SOD activity and photosynthetic fluorescence characteristics, which made the Dutch cultivar Glorioso maintain high photosynthesis efficiency under low light.
tomato; low light; photosynthetic fluorescence characteristics; stomata; chloroplast

10.3864/j.issn.0578-1752.2021.05.013
2020-05-21;
2020-08-18
國家大宗蔬菜產業技術體系北京綜合試驗站(CARS-23-G-06)、青海省科技計劃(2018-NK-103)、北京市農林科學院科技創新能力建設專項(KJCX20180705)
孟憲敏,E-mail:1475102978@qq.com。通信作者劉明池,E-mail:liumingchi@nercv.org
(責任編輯 趙伶俐)
