祁連山南麓高寒灌叢GPP 變化特征及對生長季積溫的響應
2021-03-26 06:53:20祝景彬賀慧丹李紅琴張法偉李英年楊永勝張光茹王春雨羅方林
草業科學 2021年2期
祝景彬 ,賀慧丹,李紅琴,張法偉, ,李英年, ,楊永勝, ,張光茹, ,王春雨, ,羅方林,
(1. 棗莊學院旅游與資源環境學院,山東 棗莊 277160;2. 中國科學院西北高原生物研究所,青海 西寧 810001;3. 洛陽師范學院生命科學學院,河南 洛陽 471934;4. 中國科學院高原生物適應與進化重點實驗室,青海 西寧 810001;5. 中國科學院大學,北京 100049)
由于人類對化石燃料的過度使用,致使空氣中溫室氣體含量逐漸增加,導致全球氣溫升高[1],“溫室效應”將會對陸地生態系統的碳循環過程產生深刻的影響[2-4]。青藏高原是世界上最年輕的高原,并且是我國草地分布最廣的區域[5-6]。分布在青藏高原的大面積天然高寒草地,對于整個青藏高原生態系統的碳平衡具有至關重要的作用,對全球陸地生態系統碳循環也具有重要的影響[7]。由于青藏高原低溫、高海拔的特殊環境,使青藏高原草地生態系統極其脆弱,對氣候變化和環境因素改變的響應極其敏感[8]。青藏高原高寒灌叢生態系統分布面積為0.106 × 106km2,是青藏高原高寒草地的重要組成部分[9]。因此,研究高寒灌叢生態系統的碳平衡動態,對于準確認識青藏高原高寒草地碳循環過程具有重要意義[10-11]。
生態系統總初級生產力(gross primary productivity,GPP)是指植被通過光合作用制造的全部有機質總量,是生態系統碳循環的基礎[9]。大氣中CO2濃度會對GPP 產生重要影響,而GPP 的改變也會對大氣中CO2濃度造成反饋影響,二者相互作用影響著氣候變暖的過程[7-8]。GPP 受植物本身的光合作用潛力、葉面積指數、群落結構,以及光合有效輻射、溫度、有效土壤水分、養分等諸多因子的共同影響[12]。植被生長發育需要一定的溫度(熱量)條件,植物進行光合作用過程中相關酶的生理活性以及酶促反應速度受到溫度調控[13],溫度是影響植物光合作用的關鍵因子[14-16]。但是,溫度與總初級生產力的關系較為復雜,一定程度上溫度的升高有利于光合速率的提高,但是超過其最適反應溫度又開始下降[17]。此外,有研究表明,群落生物量對生態系統熱量條件變化的反應具有一定的“時滯”性,在植被生長發育所需要的其他條件均得到滿足時,溫度越高越有利于植被的生長發育,并且當溫度累積到一定程度時,植被才能完成其整個生長發育過程,在生長季的這個溫度的累積數稱為生長季積溫(growing season degree days, GDD)。因此,分析生態系統氣溫的累加值與植被光合生產能力的關系在一定程度上更能客觀地反映其真實的響應模式[18]。
國內外對陸地生態系統碳循環方面的研究主要集中在低海拔地區,而對于高海拔地區青藏高原高寒生態系統碳循環方面的研究相對較少[19-20];此外,目前對青藏高原高寒生態系統碳平衡的研究局限于短期內的變化動態和影響機制的研究[21],而對于較長時間尺度的年際變化動態和響應機制研究相對較少[4,11]。在高寒地區的研究表明,氣候變暖提前了融雪期和植被返青期,延長了生長季長度,并促進高寒生態系統的CO2同化和植被生產[22]。近幾十年來,青藏高原經歷了顯著的變暖,但有關高寒灌叢生態系統碳平衡的研究相對較少[23]。同時,全球氣候變化的背景下,永凍層和冰川融化可能導致青藏高原不同類型草地生態系統分配格局的改變[24]。此外,研究表明在不同的時間尺度上,氣候環境因子可能對碳循環有不同的影響[11]。在不同時間尺度上分析氣候環境因子對碳通量的影響,更能準確地反映其客觀關系。因此,本研究在不同時間尺度上分析了2003?2016 年生長季利用渦度相關系統在青藏高原東北部高寒灌叢獲得的14 年連續通量數據,旨在:1)量化高寒灌叢生態系統生長季GPP 與熱量條件[GDD、Ta(氣溫)]在不同時間尺度上的變化特征;2)明確不同時間尺度上高寒灌叢生態系統生長季的GPP 對熱量條件(GDD、Ta)的響應。
1 材料與方法
1.1 試驗地概況
采集數據的渦度相關系統位于青海省海北藏族自治州門源縣境內的高寒灌叢生態系統,研究地點的具體位置屬于高寒草甸生態系統國家野外科學觀測研究站(海北站)。海北野外觀測站地處青藏高原的東北邊緣,地理位置為37°29′?37°45′ N,101°12′?101°23′ E,海拔為3 200~3 600 m。該地無明顯的四季之分,一年之中只分為冷熱兩季,在11 月至次年4 月明顯干燥寒冷,而在5 月至10 月則明顯溫暖濕潤。全年平均氣溫約為?1.0 ℃,最熱月為7 月,而最冷月為1 月。全年平均降水量為580 mm,但是降水在一年之中分布極不均衡,大約有80%的降水集中在植被生長季[25]。青藏高原高寒灌叢生態系統的植被主要由兩層組成,分別為灌木層和禾本科層。青藏高原高寒灌叢生態系統的建群種為金露梅(Potentilla fruticosa),其他群落組成物種主要包括矮嵩草(Kobreia humilis)、垂穗披堿草(Elymus nutans)、異針茅(Stipa aliena)和麻花艽(Gentiana straminea)等。青藏高原高寒灌叢生態系統的土壤為一種粘性壤土[26-27]。
1.2 研究材料及分析方法
1.2.1 通量和相關監測
利用渦度相關系統對研究區CO2通量進行長期連續的觀測(2003?2016 年)。通量觀測的主要儀器為開路式渦度相關觀測系統和常規氣象觀測系統,其中渦度相關系統包括三維超聲風速儀(CSAT3, Campbell Scientific, Logan, USA)、紅 外 氣體分析儀(LI-7500, LI-COR Lincoln, USA)和數據采集器(CR5000, Campbell Scientific, Logan, USA)。常 規氣象觀測系統觀測項目主要包括溫度、濕度、輻射、風向、風速等。光合有效輻射( photosynthetic photon flux density, PPFD)傳感器(L1190SB, LI-COR, Lincoln,USA)安裝在高1.2 m 的支架上。空氣溫度/濕度傳感器(Model HMP45C, Vaisala, Helsinki, Finland)安裝在 防 輻 射 罩 內(Model 41002, RM Young Company,Michigan, USA)。土壤溫度(soil temperature, Ts)主要是利用熱電偶溫度傳感器(107-L, Campbell Scientific,Logan, USA)分別測定5、10、20、40 和80 cm 處的Ts。5、20 和50 cm 處的土壤含水量(soil water content, SWC)利用時域反射計(Model CS615-L, Campbell Scientific,Logan, USA)測定。
生長季有效積溫(GDD)為大于5 ℃日平均氣溫的和[28]。參考以往研究經驗,將生長季定義為高寒灌叢生態系統的5 月 ? 9 月[11,14]。本研究中日尺度GPP、Ta為當天的GPP、Ta;月尺度GPP、GDD 為當月每天GPP、Ta的和;生長季尺度GPP 、GDD 為整個生長季每天GPP、Ta的和。
1.2.2 數據處理和插補
由于野外各種因素的不確定性,導致儀器運行會出現異常,導致通量塔記錄數據缺失[29]。本文利用研究時段內的輻射有效能(凈輻射與土壤熱通量之差)與湍流熱通量之比的能量閉合率來簡單評價高寒灌叢通量數據的數據質量。高寒灌叢通量數據在生長季的能量閉合率為0.78。有關能量不閉合的研究和相關的結論較多,因此本研究沒有深入討論能量不閉合的原因[4,11]。但從數據分析來看,高寒灌叢通量觀測的數據有效性較高,完全可以滿足后續的數據分析。
本研究嚴格采用ChinaFLUX 推薦的通量數據處理方法來進行數據分析[30-31]。對于缺失的通量數據,一般通過非線性經驗方程進行插補。本研究夜間通量數據(Reco,n)采用其與5 cm 深處土壤溫度(Ts)的Van′t Hoff 函數[公式(1)]進行插補,生長季白天的碳通量數據(NEE)則利用與PPFD 的Michaelis-Menten 函數[公式(2)]進行插補[32-33]。

式中:Reco,ref為生態系統在10 ℃時的參考呼吸速率,Q10為生態系統呼吸熵(呼吸隨溫度升高10 ℃所增加的倍數),反映了生態系統呼吸對溫度的敏感性。

式中:Reco,d為生態系統白天的呼吸速率,a 和Pmax分別為生態系統表觀光量子效率和潛在最大CO2同化速率(即PPFD 趨向極大時生態系統凈光合速率)。以上參數均為方程擬合值。
渦度相關系統無法直接測定生態系統總初級生產力(GPP)和生態系統呼吸(Re),需利用公式外推得到。渦度相關系統觀測的CO2通量為NEE,將夜間呼吸函數外延至白天(Reco,d),即可得生態系統的總呼吸量(Re),進而可得生態系統總初級生產力[GPP,公式(3)],日GPP 計算如公式(3)所示,日Re為白天呼吸(Reco,d)和夜間呼吸(Reco,n)之和[27]。本研究中GPP 和GDD 的關系采用線性回歸分析,利用Excel 和SPSS 20.0 軟件對數據進行統計分析。

2 結果與分析
2.1 高寒灌叢生態系統的日GPP 變化特征及其對氣溫的響應
高寒灌叢生態系統的日GPP 在2003?2016 年生長季(5 月 ? 9 月)的變化趨勢基本一致,表現為先增加后減小的單峰變化趨勢(圖1)。14 年的數據表明,日GPP 在每年7 月達到峰值,日GPP 的最大值為2009 年7 月17 日的8.02 g·(m2·d)?1。高寒灌叢在2003?2016 年生長季(5 月 ? 9 月)日平均氣溫(Ta)變化趨勢基本一致,表現為先升高后降低的單峰變化趨勢(圖1)。最高氣溫出現在2010 年7 月29 日,為17.26 ℃;最低氣溫出現在2009 年5 月1 日,為?1.93 ℃。線性回歸分析表明(圖2),日GPP 與Ta呈極顯著正相關關系(GPP = 0.41Ta+ 0.42,R2= 0.50,P < 0.001),由此表明在日尺度上氣溫越高,高寒灌叢植被的光合生產能力越強。

圖1 高寒灌叢日總初級生產力(GPP)與氣溫(Ta)的變化特征Figure 1 Variation characteristics of daily gross primary productivity (GPP) and temperature (Ta) of alpine shrubland
2.2 高寒灌叢生態系統的月GPP 變化特征及其對GDD 的響應
高 寒 灌叢 在2003?2016 年生 長 季(5 月 ? 9 月)GPP 月變化趨勢基本一致,表現為先增大后減小的單峰變化趨勢,2003?2016 年生長季的月GPP 基本都在7 月達到峰值,只有2003 年在8 月達到峰值,為130.54 g·m?2;月GPP 的最大值為2015 年7 月的值,為191.49 g·m?2,7 月GPP 的均值為165.49 g·m?2(圖3)。2003?2016 年每年5 月的GPP 為當年生長季的月GPP 最小值,5 月GPP 的均值為23.02 g·m?2。6、8、9 月GPP 的均值分別為93.47、146.68 和75.66 g·m?2。在2003?2016 年生長季(5 月 ? 9 月)的月GDD 變化趨勢基本一致(圖3),在7、8 月達到峰值,7、8 月GDD的均值分別為302.80 和302.75 ℃。5 月GDD 最小,其均值為70.70 ℃;6、9 月GDD 均值分別為206.43和107.54 ℃。

圖2 高寒灌叢日總初級生產力(GPP)與氣溫(Ta)的關系Figure 2 The relationship between daily gross primary productivity (GPP) and temperature (Ta) of alpine shrubland
線性回歸分析表明(圖4),在整個生長季的月GPP 與 月GDD 極 顯 著 正 相 關(GPP = 0.49GDD +5.93,R2= 0.84,P < 0.001),此外,分別分析了2003?2016 年生長季每個月的GPP 與GDD 的關系(表1),研究發現在5、9 月GPP 與GDD 沒有顯著相關性(P >0.05),在6、7、8 月GPP 與GDD 顯著相關(P < 0.05),并且在生長旺盛期(7、8 月)的相關性更顯著(P <0.01),在7 月GPP 與GDD 的相關性最強(R2= 0.54,P = 0.003)。這種不同月份GPP 與GDD 的相互關系存在差異(表1)可能是由不同月份之間氣候環境因子的差異造成的。因此,在生長季旺盛期(7、8 月)這種相關性最強,說明在高寒灌叢植被生長最旺盛時期,熱量條件對植被光合生產能力有更顯著的影響。整體上看,在月尺度上,生長季每個月積溫越大,越有利于生態系統總初級生產力的提高。

圖3 高寒灌叢的月總初級生產力(GPP)與生長季有效積溫(GDD)的變化特征Figure 3 Variation characteristics of monthly gross primary productivity (GPP) and growing season degree days (GDD) over alpine shrubland

圖4 高寒灌叢的月總初級生產力(GPP)與生長季有效積溫(GDD)的關系Figure 4 The relationship between monthly gross primary productivity (GPP) and growing season degree days (GDD) over alpine shrubland
2.3 高寒灌叢生態系統的生長季GPP 變化特征及其對GDD 的響應
如圖5 所示,高寒灌叢在2003?2016 年整個生長季(5 月 ? 9 月) GPP (R2= 0.64,P = 0.001)的值和GDD(R2= 0.44,P = 0.01)的值具有一定的變異性,但整體上表現為逐漸增加的趨勢。生長季GPP 的最大值出現在2014 年,為562.13 g·m?2;生長季GPP 的最小值出現在2003 年,為406.32 g·m?2;2003?2016 年的生長季GPP 均值為507.11 g·m?2。2004?2016 年整個生長季總GDD 的最大值為2016 年的1 119.15 ℃,最小值為2004 年的826.35 ℃,整個生長季GDD 的均值為975.93 ℃。線性回歸分析表明(圖6),生長季GPP 與GDD 顯著正相關(GPP = 0.35GDD + 164.99,R2= 0.39,P = 0.016),由此說明在生長季尺度上,高寒灌叢生態系統生長季有效積溫(GDD)的增大有利于提高整個生長季的總初級生產力(GPP)。
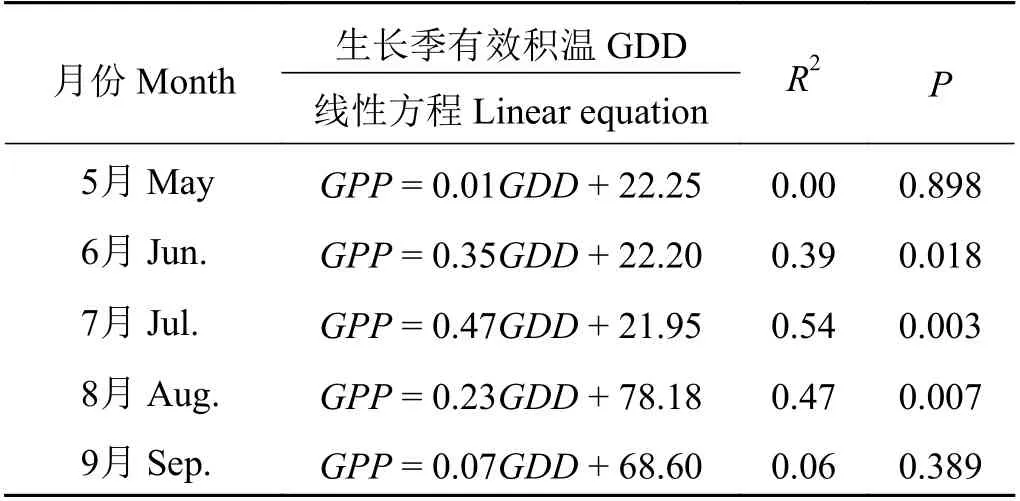
表1 高寒灌叢在生長季每個月總初級生產力(GPP)與生長季有效積溫(GDD)的相關性Table 1 Correlation between gross primary productivity(GPP) and growing season degree days (GDD) of each month in growing season over alpine shrubland

圖5 高寒灌叢的生長季(5 月 ? 9 月)總初級生產力(GPP)與生長季有效積溫(GDD)變化特征Figure 5 Variation characteristics of gross primary productivity (GPP) and growing season degree days(GDD) in growing season (May-Sep.) over alpine shrubland
3 討論

圖6 高寒灌叢的生長季(5 月 ? 9 月)總初級生產力(GPP)與生長季有效積溫(GDD)關系Figure 6 The relationship between seasonal gross primary productivity (GPP) and growing season degree days (GDD) in growing season (May-Sep.) over alpine shrubland
本研究中,GPP 和熱量條件(Ta、GDD)在日尺度、月尺度的變化趨勢都表現為先升高后降低的單峰變化趨勢,都是在7、8 月達到峰值,這和前人有關 青 藏 高 原 高 寒 灌 叢 的 研 究 結 果 基 本 一 致[4,6,10,14]。在生長季尺度上,2003?2016 年高寒灌叢的生長季GPP 和GDD 具有一定波動性,但整體上表現為逐漸增加的趨勢,這與前人對于青藏高原全球氣候變化背景下氣溫升高的研究結論相一致[7,13]。研究表明,隨著溫度的升高,生態系統GPP 有增大趨勢,但增大緩慢且具有較高異質性[13]。降水、溫度、CO2濃度和地表覆蓋變化等都會改變植物蒸騰、光合等生理活動,從而引起陸地生態系統GPP 的變化,進而影響其碳平衡。大量研究表明,熱量條件對于高寒生態系統植被的光合生產能力具有至關重要的作用,但也發現群落生物量對生態系統熱量條件變化的反應具有一定的“時滯”性。因此,分析生態系統氣溫的累加值與植被光合生產能力的關系在一定程度上更能客觀地反映其真實的響應模式[34]。所以,在全球氣候變化的背景下,研究總初級生產力對熱量條件的響應極其重要。
大量研究表明,不同生態系統的碳平衡狀態存在較大差異,這是由不同生態系統生物和非生物因素之間的差異造成的[18-19]。同時,年均溫的年際差異也導致了碳循環各過程的波動[20]。但是,過去的研究大多局限于短期的研究,研究表明在不同的時間尺度上環境因子可能對生態系統生產力有不同的影響[21,26],因此本研究在不同時間尺度上探討了GPP 對熱量條件(Ta、GDD)的響應。研究結果發現,在不同時間尺度上GPP 與熱量條件(Ta、GDD)都呈顯著正相關關系,這與柴曦等[4]、Zhao 等[13]和Li等[11]對青藏高原高寒灌叢的研究結果一致,并且Kato 等[21]、Saito 等[32]對本研究地點附近的高寒矮嵩草草甸的研究也有類似的研究結果。由此說明,高寒灌叢生態系統植被的光合生產能力直接受到熱量條件的控制,這可能是因為溫度的累積效應是打破植被休眠、植被返青和生長代謝的主要控制因素[9,21,35]。因此,高寒植被在生長代謝方面受到溫度的重要影響[36]。此外,溫度能夠影響土壤中微生物的生物量,以及相關酶的活性,由于高寒生態系統土壤中含有大量未分解的土壤有機質,并且微生物分解土壤有機質的過程對溫度極其敏感,較高的溫度能夠刺激土壤有機質的分解,為植被的生長提供更多的養分,因此間接有利于提高植被的光合生產能力[21,23]。此外,青藏高原東部高寒灌叢草氈土的水分含量達到30%以上,遠高于高原腹地的草原土,而且生長季開始就處于比較穩定的高值,受降水量的影響沒有草原土那么大[4];另一方面,灌叢植被根系要深于草甸植被根系,降低了植被根系對表層土壤干旱的敏感性,緩解表層土壤水分的缺失對根系的影響[11,13]。因此,熱量條件是高寒生態系統最為普遍的環境影響因子[21,32],它通過影響生態系統的生態過程,例如冠層發育、蒸散速率和土壤水分等來影響植被光合生產能力及其對氣候變化的響應[6, 11, 21, 32]。
但值得注意的是,本研究對2003?2016 年生長季中每個月份的GPP 與GDD 作了線性回歸分析,發現在5、9 月GPP 與GDD 沒有顯著相關性,在植被生長旺盛期(7、8 月)相關性較強,在7 月GPP 與GDD 的相關性最顯著。這可能是由在生長季中不同月份的氣候環境因子之間的差異造成的[11-14]。研究結果顯示,在5、9 月GPP 與GDD 沒有顯著相關性,這可能是因為在生長季初期和末期植被生長緩慢[6,14],光合生產能力較弱,而此時有效積溫也較小,所以有效積溫的改變對植被光合生產能力的影響并不顯著。但是,7 月份雨熱條件最好[4,19],植被生長最旺盛,植被在這個時期的光合生產能力最強,因此在這個時期有效積溫的變化,會對生態系統的總初級生產力產生極顯著的影響,說明此時熱量條件對GPP 變化的控制作用更強。總體上看,高寒灌叢生態系統的總初級生產力受到熱量條件的顯著影響,即有效積溫越大,越有利于植被光合生產能力的提高。本研究的結果表明,在全球變暖的背景下,有利于青藏高原高寒灌叢總初級生產能力的提高,但是有研究也表明溫度的升高也會增加生態系統呼吸[11],因此對于高寒灌叢生態系統的碳平衡狀態還需要進一步深入的研究。
本研究結果表明熱量條件是高寒灌叢植物生長的重要限制因子,溫度升高會促進濕潤地區植物光合作用和碳固定。但是,溫度升高會導致土壤水分蒸發的增加,加劇植被受干旱脅迫的程度,可能導致植被生產力降低[13,21],因此應加強青藏高原不同植被類型生態系統植被光合生產能力對氣候變化響應的研究。Wang 等[37]利用CENTURY 模型對松嫩平原的預測表明,一定幅度溫度的增加,有利于提高植被光合生產能力,當溫度升高超過一定閾值,將會造成植被光合生產能力的下降。所以,高寒灌叢GPP 對溫度升高是否存在閾值效應,以及具體閾值如何等問題仍需進一步深入分析。并且,高寒灌叢生態系統的GPP 不僅受到氣候變化的影響,還會受到人類活動的影響,因此應該加強對氣候變化和人類活動對高寒灌叢GPP 變化相對貢獻率的研究。此外,本研究僅探討有效積溫對高寒灌叢生態系統總初級生產力的研究,仍有一定的局限性。在今后的研究中應加強對實測生物量、碳儲量、碳通量等資料的收集和整理,為生態系統模擬模型的驗證和評價工作提供充分的依據。
4 結論
1)高寒灌叢在生長季的日GPP、Ta與月GPP、GDD 都表現為單峰變化趨勢;而生長季總GPP 與GDD 具有較高變異性,但整體上表現為逐漸增加的趨勢(P < 0.05)。
2)在日尺度、月尺度、生長季尺度上,GPP 與熱量條件(Ta、GDD)都呈顯著正相關關系(P < 0.05)。在7 月份的月GPP 與GDD 相關性最顯著(P < 0.01)。整體上看,高寒灌叢GDD 的增加有利于促進GPP的提高,暗示在全球氣候變暖的背景下將會提高青藏高原高寒灌叢生態系統植被的光合生產能力。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
小讀者(2021年2期)2021-03-29 05:03:48
科技傳播(2019年22期)2020-01-14 03:06:54
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
