聚對苯二甲酸乙二醇酯(PET)降解酶的研究進展
2021-03-31 09:49:14趙彧瑾陳仔君趙晶晶蘇婷婷王戰勇
微生物學雜志 2021年6期
關鍵詞:研究
趙彧瑾,陳仔君,趙晶晶,蘇婷婷*,王戰勇
(1.遼寧石油化工大學 石油化工學院,遼寧 撫順 113001;2.沈陽農業大學 生物科學與技術學院,遼寧 沈陽 110866)
現代社會的日益商品化以及塑料與各行業的緊密關聯,使得全球對塑料的依賴程度逐年提高。據研究統計,每年全球大約生產1億 t塑料制品,并呈逐年上升趨勢[1]。尤其在2020年于抗擊新冠肺炎疫情中,主要由塑料材料制成的手套、面罩、防護服等對病毒傳播的控制更是發揮了重要作用[2]。聚對苯二甲酸乙二醇酯(Polyethylene terephthalate, PET)由于具有優越的力學性能和良好的實用性,目前已成為廣泛使用的塑料材料之一[3],在2020年全球PET產能就突破了1億t[4]。目前PET已應用于工農業各個領域,如飲料或礦泉水瓶、薄膜以及滌綸服裝等[5],由此也產生了大量的PET廢棄物。雖然很多國家已經開始回收利用廢棄的PET,但數量很少,大部分廢棄PET未能得到有效回收,這些物質經過大氣紫外輻射、自由基氧化以及海水水解等作用發生脆化,產生肉眼不可見的微塑料,這些微塑料可被水生生物吸收,人類作為食物鏈的頂端,攝入這些水生生物后,可能會富集大量的微塑料于體內,對人體健康造成難以預計的危害[6]。眾所周知,微生物在污染物的清除和生態系統的物質循環中發揮著重要作用。自20世紀90年代以來,微生物酶在降解高分子材料方面的應用已開始引起人們的關注。酶的催化降解為解決塑料造成的環境問題提供了一個獨特的方案。自從2005年Müller等[7]從嗜高溫放線菌的褐色嗜熱裂孢菌(Thermobifidafusca)中發現了第一個PET降解酶,到現在已有多個研究團隊對多種微生物來源的PET降解酶進行了一系列的研究。在相關研究領域尤其是針對PET降解酶的研究取得了一定進展。本文就PET降解酶及其降解機制進行了綜述,并進一步對其在PET廢棄物處理的應用及發展前景方面進行了探討。
1 脂肪酶
脂肪酶又稱甘油酯水解酶,因其催化水解長鏈(>C10)的甘油三酯而廣為人知,屬于羧基酯水解酶類,其特點是界面活化現象。脂肪酶可通過增強PET織物的潤濕性、染色性和吸收性,在一定程度上對其進行降解[7]。2005年Müller等[7]首次報道,源于褐色嗜熱裂孢菌芳香族聚酯酶能夠降解聚己二酸-對苯二甲酸丁二酯,在后來的研究中發現其也可以降解PET。德國研究者從褐色嗜熱裂孢菌的培養上清液中純化出一種可水解聚丁烯對苯二甲酸-共己二酸酯的降解酶(BTA-1)[8],在55 ℃條件下該降解酶能在3周內將商業PET飲料瓶和PET顆粒水解40%~50%。此后Lykidis等[9]對褐色嗜熱裂孢菌_ YX的全基因組進行了測序,確定了兩個三酰基甘油脂肪酶基因的存在,其中Tfu0883與BTA-1完全匹配。Eberl等[10]通過陽離子染色和X射線光電子能譜等技術確定了從綿毛嗜熱絲孢菌(Thermomyceslanuginosus)中分離得到的脂肪酶不僅能水解PET組成的單體寡聚物——雙(苯甲氧基乙基)對苯二甲酸乙二醇酯(bis-(benzoyloxyethyl)terephthalate, 3PET),還能水解多聚物,如PET織物和薄膜。但是脂肪酶在活性位點上具有“蓋子”結構,這種結構的存在抑制了大分子進入活性位點的可能性,使得聚合物與催化中心結合困難,導致脂肪酶對PET的水解活性偏低。該研究還表明非離子洗滌劑Triton X-100能夠激活脂肪酶的“蓋子”結構,使其對3PET的降解能力明顯增強。
2 酯 酶
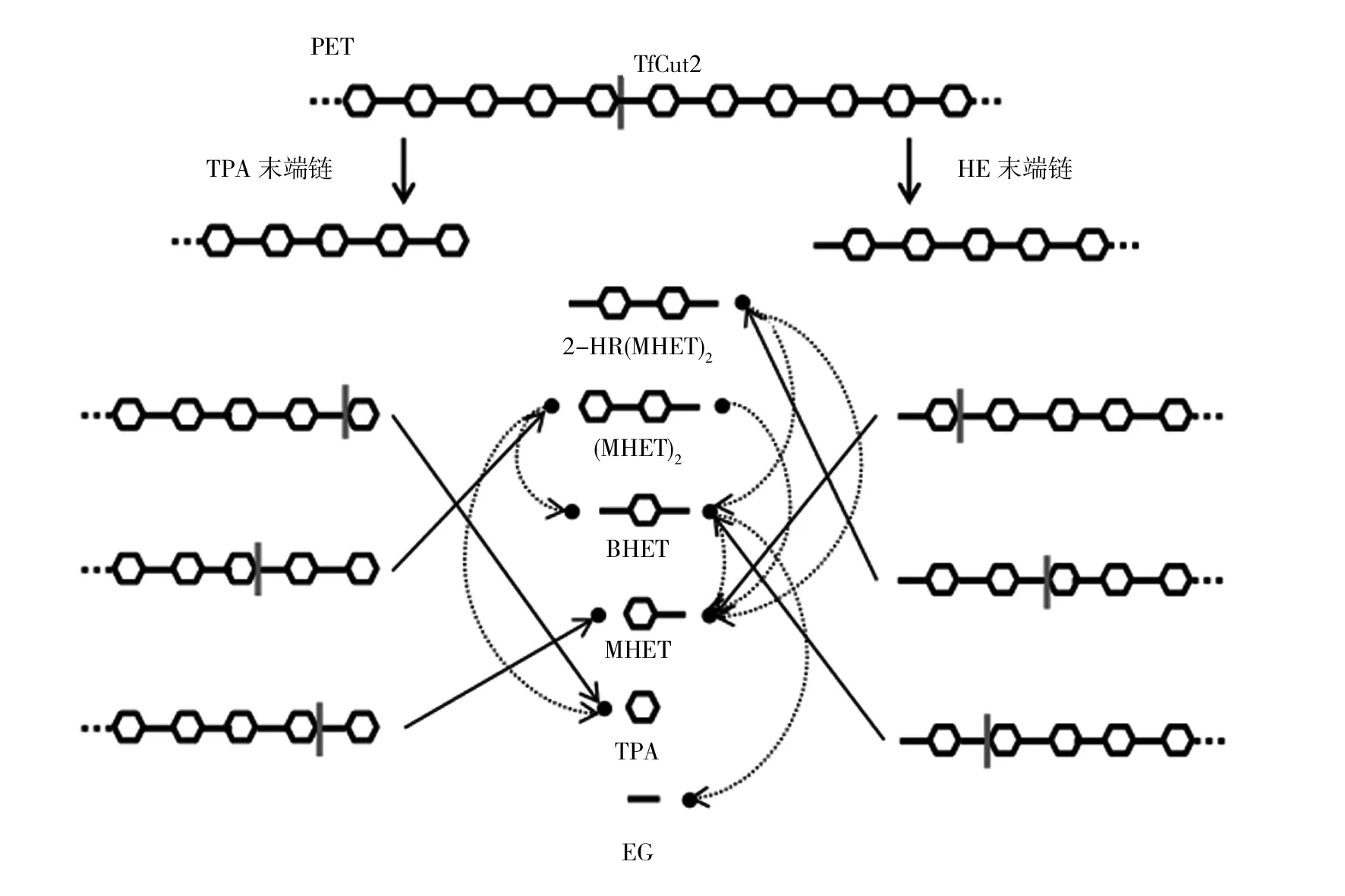
酯酶通常裂解水溶性短鏈甘油酯( 2010年Billig等[11]將嗜熱放線菌褐色嗜熱裂孢菌KW3b在含有PET纖維的培養基中培養,產生了分子量為50 kDa的酯酶(TfCa)。TfCa能夠水解回收的PET薄膜,且TfCa對短鏈和中等鏈長(C2~C8)的對硝基苯基酯均具有水解活性,但對長鏈酰基酯沒有裂解作用(C16)。相比之下,此前報道的褐色嗜熱裂孢菌DSM 43793角質酶(TfH)對C2~C6酰基鏈和C8、C10、C12、C18酰基鏈酯均具有酶促活性,因此,該角質酶同時具有酯酶和脂肪酶的特性[8]。在之后的研究中Ribitsch等[16]發現來自裂孢菌屬(Thermobifida)Thh_Est的酯酶(Thh_Est)對PET表現出有效的表面水解,且其作用與來自裂孢菌屬的角質酶相似。 2016年Yoshida等[17]從PET瓶回收廠收集了包括沉淀物、土壤、廢水和活性污泥等250個樣本,從中篩選出一種可利用低結晶度(1.9%)PET膜作為主要碳源的菌株Ideonellasakaiensis201-F6。將該細菌產生的PETase在30 ℃下與PET薄膜(1.9%)孵育6周后,將PET薄膜降解為低分子量的低聚物或單體,如對苯二甲酸雙(2-羥乙基)酯(bis(2-hydroxyethyl)terephthalate, BHET)和對苯二甲酸單(2-羥乙基)酯(mono(2-hydroxyethyl)terephthalate, MHET)。Yoshida等還比較了PETase和部分角質酶的催化效率,發現PETase的活性優于其他角質酶。研究中還發現了一種MHET水解酶(MHETase),能與PETase配合,將PET的主要中間產物MHET特異性水解成對苯二甲酸(terephthalic acid, TPA)和乙二醇(ethylene glycol, EG),對PET的完全降解起著重要作用,具體水解過程如圖1所示[17]。雖然PETase已經引起了相當大的關注,但相比于其他的PET降解酶[18-19],PETase的降解能力和穩定性較差,37 ℃下其大部分降解活性在12 h內基本消失[19-20]。Cui等[19]發現PETase的降解能力僅發生在微摩爾水平,并將PETase突變為更嗜熱的類型DuraPETase,雖然結果表明該突變酶在60 ℃高溫下孵育3 d仍能保持活性,且在溫和溫度下對半結晶PET薄膜的降解率達到23%,但綜合來看其熱穩定性和降解率仍然偏低。 圖1 源于Ideonella sakaiensis 的PETase降解PET的過程[17] 角質酶是一種α/β-水解酶,屬于胞外絲氨酸酯酶,存在于某些細菌和真菌中。最初被發現于某些植物病原菌和昆蟲病原菌中,其中以源于腐皮鐮刀菌(Fusariumsolani)的角質酶被研究的最為廣泛[21-22]。Carvalho等[23]對角質酶在水解和合成反應中的生物催化潛力進行了總結,并通過對聚酯降解的研究,發現角質酶在聚酯的降解中起著決定性作用[24-25]。2009年,Ronkvist等[26]發現一種嗜熱真菌特異腐質霉(Humilicainsolens)的角質酶(HiC)能夠在70 ℃下完全降解非晶態PET,而且HiC還具有很好的熱穩定性。同時Ronkvist等也證明了門多薩假單胞菌(Pseudomonasmendocina)的角質酶(PmC)和腐皮鐮刀菌的角質酶(FsC)均能夠將PET降解成TPA和EG。2012年,Sulaiman等[27]利用宏基因組方法從植物堆肥中克隆了一種能夠降解PET的角質酶(LCC),LCC被認為可能是來自堆肥中的放線菌。Danso等[28]開發了一種搜索算法,從各種數據庫中識別出504個可能的PET降解酶候選基因。其中包含了從IMG數據庫中獲得的108個海洋宏基因組和25個陸地宏基因組中超過16 GB的序列信息,檢測到了349個可能的PET降解酶,并表征了兩種具有較高應用潛力的新型耐高溫酶。基于氨基酸的相似性對這504種候選酶的聚類表明,這些PET降解酶主要來自放線菌門、變形菌門(β、δ和γ)和擬桿菌門。值得注意的是,在海洋環境中,隸屬于擬桿菌門的細菌似乎是PET降解酶基因的主要宿主。在陸地環境中,PET降解酶主要來自放線菌。此外,基于數據庫和宏基因組的分析還發現,PET降解酶是一類相對罕見的酶,雖然數據庫和宏基因組涵蓋了陸地和海洋環境,但是這些環境的某些特殊地區仍有待充分分析,特別是極端環境,如火山、兩極地區、深海和湖泊、酸性和堿性環境以及高鹽度地區。此外,還有研究證明了無脊椎動物體內的微生物群也有可能是塑料降解酶的重要來源[29-30]。 總之,降解PET的角質酶一般都具有良好的降解活性和熱穩定性。其中來自嗜熱放線菌和綠色糖單孢菌、植物堆肥(可能是放線菌的混合基因)和嗜熱真菌特異腐質霉的角質酶被認為是嗜熱PET降解酶,到目前為止,從特異腐質霉中提取的商業化角質酶對非晶態PET薄膜的水解效果最為顯著,但其具體的水解途徑和其他因素尚未有報道。此外,與脂肪酶相比,角質酶具有開放的、淺的活性位點,沒有“蓋子”結構,可以容納大量的聚酯鏈,并使其與活性絲氨酸在催化三聯體(Ser-His-Asp)處接觸。Joo等[31]將催化三聯體中的Ser、His和Asp突變為Ala,發現角質酶對PET薄膜和BHET的降解失去了催化活性,說明這3個殘基是酶催化反應的關鍵組成。 角質酶最早發現于植物和昆蟲病原真菌。在研究過程中逐漸發現角質酶不僅能夠降解植物角質還具備降解脂肪族聚酯(尤其是PET)的能力。2005年,Silva等[32]利用腐皮鐮刀菌角質酶對幾種合成纖維的表面進行改性,發現該角質酶可將PET水解為TPA和EG,而且反應最佳溫度低于50 ℃。Yang等[33]從太瑞斯梭孢殼霉中分離得到一種低分子角質酶(TtcutA),研究發現TtcutA能夠有效降解各種酯類聚合物,包括PET、聚己內脂和聚丁二酸丁二酯。重要的是太瑞斯梭孢殼霉能夠在50 ℃或以上的條件生長良好。關于PET的酶促表面修飾的研究大約開始于40年前,并在隨后的許多研究中表明真菌角質酶適合用于PET織物/纖維的改性[18,25]。這些真菌角質酶屬于表面修飾酶,并沒有直接降解PET的特性,而是能夠將PET薄膜或纖維表面酯鍵部分進行酶解導致表面性質發生改變,使其親水性能提高。唯一例外的是一種商品化真菌角質酶HiC,其最佳催化溫度為75~80 ℃,可在70 ℃下高效水解非晶態PET[26]。相比之下,其他真菌角質酶的最佳反應溫度一般為50 ℃左右。因此,其對非晶態PET的降解率非常低。 在已報道的PET降解酶中大多數角質酶均來自于放線菌。Thumarat等[34]報道了源自白色高溫雙歧菌(Thermobifidaalba)AHK119的兩種角質酶基因(est1和est119)的遺傳圖譜,兩種角質酶具有95 %的同源性和98%的相似性。目前報道的所有裂孢菌屬微生物均顯示存在角質酶基因且基因序列相似度很高,因此,具有降解PET能力的角質酶可能均來自裂孢菌屬微生物[8]。Kawai等[35]從綠色糖單孢菌AHK190中克隆出一種角質酶(Cut190)基因,并實現了其在大腸埃希菌(Escherichiacoli)中的表達,且Ca2+能夠使該酶在75 ℃仍然具有較高的熱穩定性。除此之外,之前提到的源于褐色嗜熱裂孢菌[7]的角質酶和堆肥宏基因組的LCC[27]也是為數不多的嗜熱角質酶,與其他角質酶相比,其具有較高的水解活性和熱穩定性,這可能是因為它們均來自于高溫環境。 角質酶尤其是放線菌角質酶能夠降解PET薄膜,且可以有效地將低結晶度的薄膜完全降解成對人體無害的CO2和H2O等。PET表面親水性越高,降解PET所需溫度就越低,因此對PET表面進行改性可使PET更容易被降解。而PET降解酶的作用也可以有助于PET的表面親水,因此可以利用這一點對PET表面進行改性,篩選鑒定或設計更有效的PET降解酶也已成為當前的PET酶促降解研究的工作重點。 基于Joo等[31]對PET降解酶結構觀察和生化研究,提出了以下PET降解機制(圖2)。以PET降解酶的代表性角質酶TfCut2[36]為例,PET的初始降解是由TfCut2的殘基Y60和M131的主鏈氮原子與底物PET的羰基氧原子形成氧陰離子空穴,I178和Y60為底物PET提供疏水作用力,使易裂解的酯鍵靠近催化三聯體中的活性絲氨酸[1]被催化斷裂,進而形成兩條末端不同的PET鏈(TPA末端鏈和HE末端鏈)。隨后TPA末端鏈和HE末端鏈被降解成多種PET二聚體或單體,如2-HE(MHET)2、(MHET)2、BHET、MHET、TPA等,最終被消化為MHET、TPA和EG形式的單體。該過程說明PET降解過程中TPA的積累主要來源于TPA末端鏈的末端消化步驟。 圖2 角質酶PET降解機制[31] 目前,許多提高PET降解性能的研究實驗正在接近實際可應用的水平,現有的某些PET降解酶能夠有效地降解非晶態PET。最近,法國科學家Tournier等[37]通過分子改造后的手段將先前報道的LCC進行優化以提高其酶活性和熱穩定性。經10 h以上的作用,PET至少有90%可被改造后的LCC解聚成單體(TPA),這一改造酶的性能優于之前報道的所有PET水解酶,包括Ideonellasakaiensis201-F6中的PETase以及最近引起關注的改造酶[31,38-40]。此外,該酶催化所產生的PET單體經過純化后再次合成PET,最終制成塑料瓶,從而實現了循環經濟。 到目前為止,已發現的PET降解酶分別為脂肪酶、酯酶和角質酶等,其中角質酶是研究最廣泛也是最有效的一類PET降解酶,角質酶不僅可以使PET表面改性,還能夠在適宜溫度下顯著降解PET。同時角質酶有優于其他酶類的晶體結構,因此降解PET的角質酶具有巨大的發展潛力。 目前,對PET降解酶的研究已有一定的進展,根據已報道的降解酶的晶體結構及其降解機理,有望篩選或設計出更有效的PET降解酶。盡管高結晶度和PET水瓶材料的新型聚酯降解酶的開發仍然具有挑戰性,但非晶態PET薄膜(主要是包裝材料)的降解已經能夠很好地實現。因此,從環境中篩選新的PET降解酶仍是十分必要的,事實上目前這一領域的研究還不夠充分。未來PET生物降解研究的工作重點應該集中在兩個主要方面:①改善降解酶的熱穩定性:為了解決這一問題,既需要篩選新的高效降解酶,也需要對已經存在的或新鑒定的降解酶進行分子改造。此外,還必須研究降解產物是否會產生抑制。②PET材料的改性:在不改變材料基本性能的前提下,通過化學或物理方法控制分子量、降低結晶度或者增加表面積,使得PET更易被降解酶作用。這些工作需要生物化學和高分子化學等多個領域的研究人員共同完成,如果能解決上述這些挑戰性的工作,使得無論是非晶態還是高結晶度PET都可以進行酶促降解并實現產物回收利用,將有助于從根本上解決白色污染帶來的環境污染問題,實現循環經濟發展。3 PETase與MHETase

4 角質酶
4.1 角質酶來源
4.2 角質酶PET降解機制
4.3 角質酶的分子改造
5 展 望
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19