毛烏素沙地油蒿光系統(tǒng)II多時間尺度的環(huán)境響應特征
2021-04-02 08:16:10李鑫豪徐銘澤高圣杰魏寧寧查天山
農業(yè)工程學報 2021年2期
靳 川,蔣 燕,李鑫豪,徐銘澤,高圣杰,魏寧寧,賈 昕,田 赟,查天山
(1. 北京林業(yè)大學水土保持學院寧夏鹽池毛烏素沙地生態(tài)系統(tǒng)國家定位觀測研究站,北京 100083;2. 北京林業(yè)大學水土保持國家林業(yè)局重點實驗室,北京 100083)
0 引言
作為維持生態(tài)系統(tǒng)物質代謝和能量循環(huán)的基礎,植物光合作用非常容易受到環(huán)境波動的影響,不同強度和持續(xù)時間的環(huán)境脅迫會協(xié)同抑制光合過程[1-2]。光系統(tǒng)II(Photosystem II,PSII)能量分配被認為是光合過程中最為敏感的部分,PSII天線色素分子吸收光能有3個去向:1)進行光化學反應,通過碳同化合成有機物[3];2)通過葉黃素循環(huán)以熱能形式將過剩光能耗散[4-5];3)重新釋放一個光子,即產生葉綠素熒光[6-7]。3種途徑相互競爭權衡,且受光合有效輻射(Photosynthetically Active Radiation,PAR)、空氣溫度(Ta)、飽和水汽壓差(Vapor Pressure Difference,VPD)和土壤含水率(Soil Water Content,SWC)等環(huán)境因素影響[5,8-9]。上述環(huán)境變量通過各自波動周期、頻率和振幅調控植物PSII能量分配[10-11]。在分-秒時間尺度上,PAR增加激活葉黃素循環(huán),植物通過改變調節(jié)性熱耗散(ΦNPQ)和非調節(jié)性熱耗散(ΦNO)的比例耗散過剩光能[12];在小時時間尺度上,VPD升高可導致蒸騰加速,植物調節(jié)氣孔減少水分損失[13],同時也不可避免地抑制了實際光化學效率(ΦPSII);在日尺度上,由PAR、Ta和VPD的天循環(huán)驅動PSII能量分配[14];在幾天至數月時間周期上,高輻射、寒潮、熱浪和降水等天氣事件以及植物的物候期都會導致PSII能量分配變化[10,15]。盡管有上述認識,但目前關于PSII能量分配的相關研究大多集中在室內短期控制試驗,難以模擬自然中真實的環(huán)境動態(tài)[16-17]。具有高太陽輻射、極端溫度和水文梯度跨度大等特點的荒漠生態(tài)系統(tǒng)則更加復雜[18]。荒漠夏季光照遠超植物所需,導致光合機構產生氧化損傷[19];極端溫度抑制了相關酶的活性,造成光合效率降低[20-21];空氣干濕循環(huán)引起植物氣孔調節(jié)[13,22],導致熒光淬滅[23]。另外,荒漠植物也常因長期缺水造成PSII失活,產生可逆的光損傷[20-21,24]。因此,荒漠植物PSII能量分配與各環(huán)境因子在多時間尺度上很可能具有明顯關聯特征。
頻譜處理可檢驗數據序列的時間動態(tài)格局[25],如小波分析。小波分析方法已經廣泛應用于多個生態(tài)相關領域,適用于探究多組時間序列數據之間關系[25-27]。然而,目前幾乎沒有研究將小波分析應用于葉綠素熒光的長期監(jiān)測數據上,尤其缺乏植物對環(huán)境波動在多時間尺度下響應過程和調節(jié)機制的理解認識。因此,本研究對寧夏鹽池毛烏素沙地優(yōu)勢荒漠物種油蒿(Artemisia ordosica)進行葉綠素熒光長期原位連續(xù)監(jiān)測,采用小波分析方法探究油蒿PSII能量分配在多時間尺度下的環(huán)境響應特征,旨在為荒漠植物對不同環(huán)境脅迫的敏感程度、適應能力和生理可塑性提供數據支撐,同時為荒漠地區(qū)植被恢復物種選擇提供理論依據。
1 材料與方法
1.1 研究區(qū)概況
研究區(qū)位于寧夏吳忠市鹽池荒漠生態(tài)系統(tǒng)定位研究站(37°53"08""N,107°25"46"" E,平均海拔1530 m),是半干旱區(qū)和干旱區(qū)之間的交錯帶,由于過度放牧等人為因素該區(qū)植被已嚴重退化,近20年的封育使該區(qū)植被得到了有效的恢復,這也被認為是該區(qū)荒漠化逆轉的重要標志。該區(qū)屬溫帶大陸性季風氣候,平均氣溫為8.3 ℃,晝夜溫差大,全年晝夜平均溫差可達25 ℃以上。年平均潛在蒸散量為2024 mm,年平均降水量為287 mm,降雨稀少且不均勻,主要集中在6—9月,4—5月容易發(fā)生春季干旱。土壤以風沙土和黑壚土為主,密度為1.54 g/cm3。盛行西北風,年平均風速2.6 m/s。研究地植被群落以旱生植物為主,優(yōu)勢灌木物種有油蒿(Artemisia ordosica)、沙柳(Salix psammophila)和楊柴(Hedysarum mongolicum),觀測期內相對覆蓋度分別為53 %、17 %和12 %。
1.2 試驗材料
油蒿廣泛分布于中國北部及西北部荒漠地區(qū),因其根系發(fā)達耐沙埋、耐旱,在固定流沙和促進群落演替中發(fā)揮著重要作用[17]。油蒿通常在4月萌發(fā),8月積累生物量達到最大,9月末開始凋落。樣地內油蒿平均密度為1.43株/m2,平均高度為34.7 cm,平均冠幅為0.64 m。選取3株長勢相同且具有代表性(高度為50 cm,冠幅為0.8 m,約50分枝左右)的油蒿作為樣本植株。避免監(jiān)測期內人為因素的干擾,保證樣本植株處于完全自然生長的狀態(tài),并進行物候的同步觀測。
1.3 葉綠素熒光監(jiān)測與計算
于2016年4月28日至10月3日,采用多通道連續(xù)監(jiān)測熒光儀(MONITORING-PAM2000,WALZ公司,德國)對油蒿樣本植株進行24 h原位連續(xù)監(jiān)測,測量間隔為30 min。用熒光探頭的特制葉夾固定葉片,調整探頭方位使葉片充分受光,每隔7 d定期檢查葉片并及時更換。測定葉綠素熒光參數包括飽和脈沖下熒光(Fm")和實時熒光(Fs)。將夜間的Fm"、Fs分別作為暗處理最大熒光(Fm)和暗處理最小熒光(Fo)。根據Kramer公式計算PSII能量分配[28],實際光化學效率(ΦPSII,%)、調節(jié)性熱耗散(ΦNPQ,%)和非調節(jié)性熱耗散(ΦNO,%)的計算方法如下:
式中Fm"為飽和脈沖下熒光,Fs為實時熒光,Fm為暗處理最大熒光。
采用式(4)計算最大光化學效率(Fv/Fm)
式中Fo為暗處理最小熒光。
1.4 環(huán)境氣象因子監(jiān)測與計算
光合有效輻射由距熒光儀100 m處通量塔(高度為6 m)配有的輻射傳感器(PAR-LITE,Kipp & Zonen公司,荷蘭)測得。空氣溫度和相對濕度由塔上溫濕度傳感器(HMP155A,Vantaa公司,芬蘭)測得,采用式(5)和式(6)[16]計算飽和水汽壓差(VPD,kPa)
式中e為飽和空氣壓力,kPa;RH為相對濕度,%;Ta為空氣溫度,℃。
土壤含水率由樣地內土壤溫濕度傳感器(ECH2O-5TE,Decagon Devices公司,美國)測得,深度為30 cm。降雨量由距樣地200 m處空地內雨量筒(TE525W,Campbell公司,美國)測得。所有環(huán)境氣象因子測量間隔均為30 min。
1.5 數據處理與分析
本研究使用連續(xù)小波變換(Constant Wavelet Transform,CWT)和小波相干性(Wavelet Coherence,WTC)分析熒光參數的時間序列。CWT是將時間序列數據在固定間隔的變量進行平移或伸縮等變換,用母小波來表示。CWT可以反映時間序列數據在不同時間尺度的波動[29],數量為N的離散變量xn的積分譜 ()xnWs定義為
式中δt為時間間隔,為復共軛尺度,s為小波變換尺度,n為數組1至N,n"為正整數取值。本研究選取Morlet小波作為母小波,采用式(8)計算xn的小波功率譜(Sn)[26,30],得到其貢獻最大的時間周期
定義2個時間序列數據之間的交叉小波功率譜(Cn),相位角譜(An)和WTC功率譜(Rn2)[26,30]
式中x為環(huán)境因子,y為植物熒光參數,為變量x與y的交叉小波變換,S為時間尺度的平滑處理,和分別為的實部和虛部[31](在時頻域圖中,An可以用箭頭來繪制,若x與y正相關,向右的箭頭表示x與y的相位是一致的,向左則相反。而向下的箭頭表示y滯后于x90°或270°,向上的箭頭表示x滯后于y90°或270°。每360°為1個時間周期)。
2 結果與分析
2.1 環(huán)境因子與油蒿PSII能量分配參數動態(tài)
PAR求日最大值和平均值,Ta求日最大值、日最小值和日平均值,VPD和SWC求日平均值,降雨量求日總值。如圖1所示,觀測期內PAR和Ta趨勢相近,在5 —7月呈上升趨勢,8—9月呈下降趨勢。PAR最高達1963μmol/(m2·s)(6月8日),7月和8月出現持續(xù)多天低值(PAR<400μmol/(m2·s))。Ta最高值達32.1 ℃(7月29日),最低值為2.9 ℃(5月6日),出現1次大幅降溫事件(8月22日)。VPD在5—7月呈周期性波動,8—9月較低,日平均值最高達2.5 kPa(7月25日)。SWC整體呈持續(xù)下降趨勢,在8月20日降雨量達到52.6 mm的降水后有較大增加。如圖2所示,ΦPSII在7月和8月有2處明顯的下降,日平均值最低為0.45%(7月16日)。ΦNPQ和ΦNO趨勢大致相同,7月和8月有2處明顯增高。ΦNPQ日平均值最高達23 %(7月16日),ΦNO日平均值最高達36%(7月3日)。Fv/Fm呈周期性波動,最高0.82(5月31日),最低0.68(5月3日)。觀測期內,各葉綠素熒光參數和環(huán)境因子具有明顯季節(jié)波動。
PAR、Ta和VPD都呈現先升高后降低的日變化趨勢,PAR峰值在13:00左右,Ta和VPD峰值約在16:00。PAR在5—7月較高,Ta在5月和9月較低,VPD在6—7月較高。ΦPSII呈現與ΦNPQ和ΦNO相反的日變化趨勢,峰值出現時間與PAR一致。ΦPSII在6—7月較低,ΦNPQ在5 —7月的峰值較高,ΦNO在7月的峰值較高(圖3)。
2.2 環(huán)境因子與油蒿PSII能量分配參數在各時間周期內變化
除SWC以外,所有變量全局小波功率譜在1 d時間周期均出現強峰值,并且在4~40 d時間周期內分別出現多次振蕩。葉綠素熒光參數(ΦPSII、ΦNPQ和ΦNO)峰值相對環(huán)境因子(PAR、Ta、VPD和SWC)較小。SWC既無日尺度譜峰,也無季節(jié)尺度譜峰。變量PAR、ΦPSII、ΦNPQ和ΦNO在小于1 d的時間周期內出現多次峰值,但其功率遠低于1 d時間周期處(圖4)。
小波功率譜可以清楚地顯示各變量在觀測期內的時頻域局部特征。在0.5 d時間周期處,PAR出現顯著小波功率區(qū)域。在1 d時間周期處,PAR、Ta和VPD均出現顯著小波功率區(qū)域,區(qū)域非連續(xù)且強烈震蕩。Ta在4~32 d周期內、VPD在8~32 d周期以及SWC在1~32 d周期內均出現小波功率值的升高(圖5)。在1 d時間周期處,ΦPSII、ΦNPQ和ΦNO均出現強烈震蕩的顯著區(qū)域。ΦPSII在8~32 d周期內、ΦNPQ在4~32 d周期以及ΦNO在16~32 d周期內也呈現小波功率值的升高(圖6)。
2.3 油蒿PSII能量分配參數與環(huán)境因子的小波相關性
油蒿PSII能量分配參數與環(huán)境因子在多個時間尺度上存在顯著小波相關區(qū)域,分布不均且波動明顯。在日尺度上,ΦPSII與PAR、Ta、VPD和SWC小波相關區(qū)域相位角分 別 為(12.32±3.56)°、(47.88±21.20)°、(73.25±13.01)°和(183.69±96.81) °,即ΦPSII滯后于PAR、Ta、VPD和SWC分別為(0.82±0.24)、(3.19±1.41)、(4.88±0.87)和(12.25±6.45) h(圖7;表1)。ΦNPQ滯后于PAR、Ta、VPD和SWC分別為(0.62±0.27)、(3.45±1.57)、(4.94±2.64)和(11.87±4.98) h(圖 8;表1)。ΦNO滯后于PAR、Ta、VPD和SWC分別為(0.72±0.32)、(3.27±1.43)、(3.83±1.69)和(8.07±6.97) h(圖 9;表 1)。油蒿PSII能量分配參數(ΦPSII、ΦNPQ和ΦNO)滯后于PAR、Ta、VPD和SWC分別平均為43 min、3.3、4.6和10.72 h。
在季節(jié)尺度上,ΦPSII滯后于PAR、Ta、VPD和SWC分 別 為(13.64±5.54)、(9.87±3.47)、(7.68±1.99)和(23.47±12.46) d(圖7;表2)。ΦNPQ滯后于PAR、Ta、VPD和SWC分別為(16.05±7.65)、(3.52±2.73)、(9.90±5.64)和(17.89±14.80) d(圖8;表2)。ΦNO滯后于PAR、Ta、VPD和SWC分別為(14.44±6.12)、(8.23±4.47)、(8.73±5.05)和(23.73±10.97) d(圖9;表2)。油蒿PSII能量分配參數分別滯后于Ta、VPD、PAR和SWC 7.2、8.8、14.7和21.6 d。
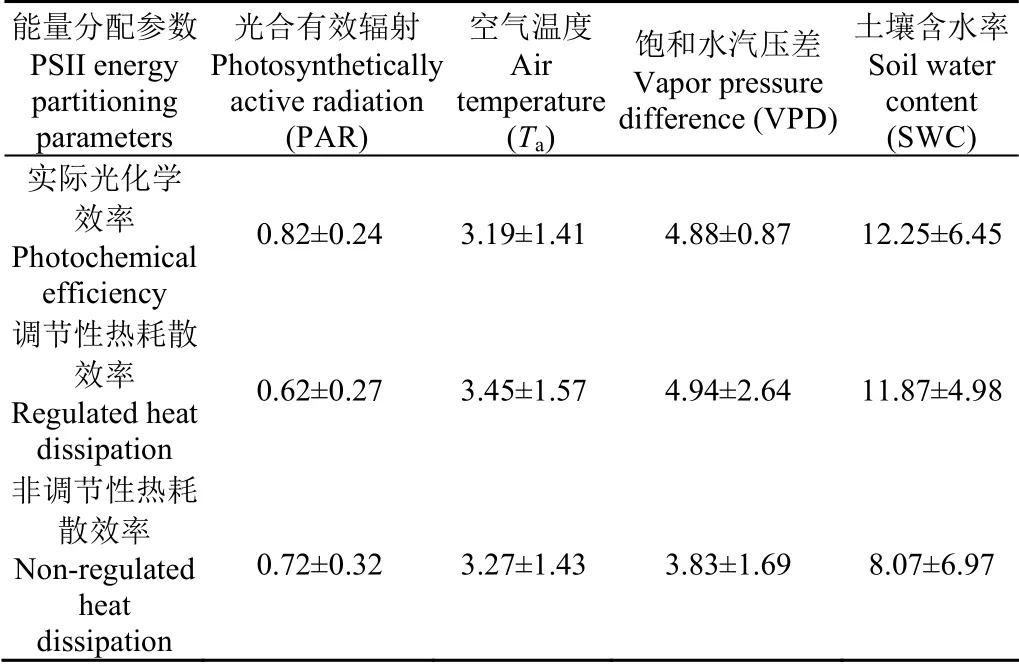
表1 PSII能量分配參數在日尺度上滯后環(huán)境因子的時間 Table 1 Lag time of PSII energy partitioning parameters relative to environmental factors on daily scale h
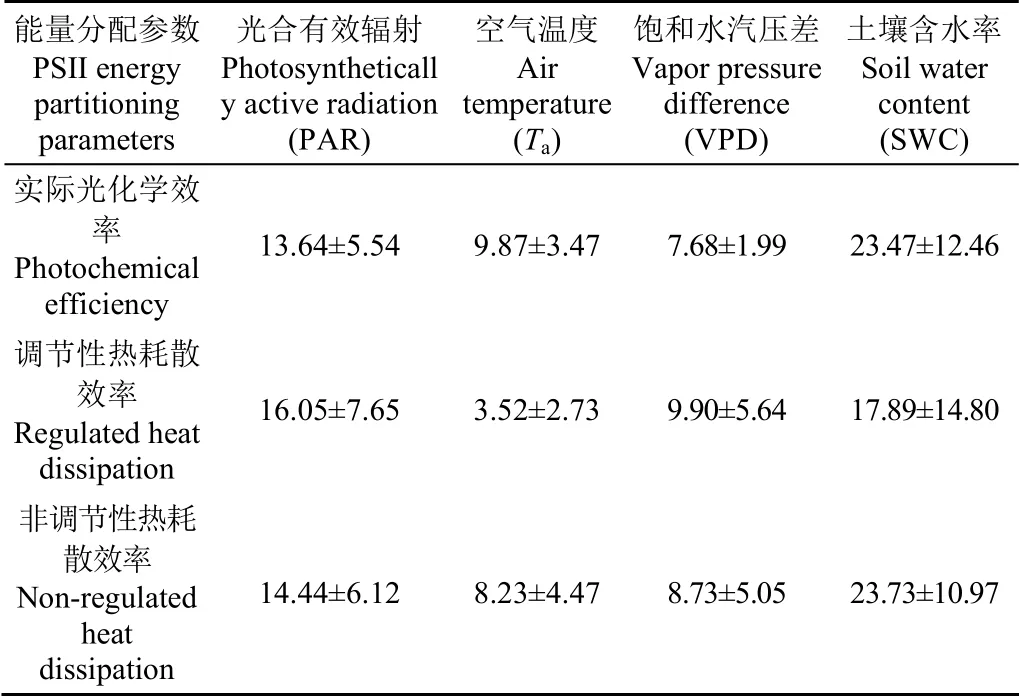
表2 PSII能量分配參數在季節(jié)尺度上滯后環(huán)境因子的時間 Table 2 Lag time of PSII energy partitioning parameters relative to environmental factors on seasonal scale d
3 討論
植物通過熱耗散機制消耗光合過程中的過剩光能,減少活性氧以避免產生光損傷[23]。將熱耗散分為ΦNPQ和ΦNO可獲得更精準的定量信息,ΦNPQ表征熱耗散等光保護調節(jié)機制的強弱[28],ΦNO則指示光抑制或光損傷的程度[10,28]。日尺度上,油蒿PSII能量分配參數與各環(huán)境因子均有很強相關性(圖7~圖9),對PAR變化最敏感且受其主導。PAR增加導致更多過剩光能需要耗散,因此ΦNPQ和ΦNO與PAR正相關,同時不可避免抑制了ΦPSII,這一系列過程是瞬間發(fā)生的[5,16]。油蒿ΦPSII在12:00左右發(fā)生明顯降低,在夜晚恢復(圖3)。表明在高輻射下,雖然油蒿光化學反應過程受到了限制,但其PSII反應中心實際并沒有產生損害[32-33]。這與同地區(qū)不同物種相關研究結果不一致,研究發(fā)現沙柳和楊柴因高輻射表現出嚴重光抑制,并造成永久性光損傷。這可能與植物形態(tài)學有關,沙柳葉呈葉線形,楊柴為互生扁平小葉,而具有針形葉的油蒿則有利于避免過量太陽輻射。油蒿PSII能量分配分別滯后Ta和VPD 3.3和4.6 h(表1)。研究表明Ta降低抑制葉綠體捕光蛋白酶活性,從而影響植物光合生理狀態(tài)[20-21]。清晨ΦPSII下降(圖3),此時Ta處于全天最低,油蒿PSII光反應相關酶活性受到抑制,導致ΦPSII早于PAR開始降低[21,34]。VPD增加導致氣孔調節(jié),蒸騰速率降低[13,22],抑制了水的裂解[24],也提高了跨類囊體質子梯度[35],最終造成ΦPSII降低,ΦNPQ和ΦNO增加。油蒿PSII能量分配在1 d時間周期內滯后SWC約10.72 h,遠高于其他環(huán)境因子(表1)。這說明短時間內油蒿對于土壤水的敏感程度較低[10,34]。
荒漠植物需要應對干旱、高輻射和極端溫度等多種環(huán)境脅迫[5,18,36],PSII長期適應機制的形成需要幾天甚至幾周時間[10,17],主要通過調整光合色素濃度、葉綠素a/b比例或PSII特定蛋白質含量等途徑實現[10,37]。Fv/Fm是表征植物PSII受脅迫程度的關鍵指標,通常情況下在0.75~0.85之間,發(fā)生下降則表示PSII 受損失活[34]。油蒿Fv/Fm和PSII能量分配季節(jié)波動明顯,但始終維持在一個整體水平上(Fv/Fm約為0.78)(圖2)。研究認為油蒿可以通過自身修復調節(jié),使其PSII恢復到正常狀態(tài)[16,28]。研究發(fā)現,油蒿PSII能量分配受物候影響,5月初處于萌芽期,葉綠素含量及活性低,對外界抵抗力較差,易產生光抑制[10,23]。7—8月受高溫高輻射影響,產生光抑制造成PSII損傷,此時油蒿處于長勢最旺盛的花果期,其生物量和葉片功能性狀達到最佳,PSII處于不斷受損和修復的過程[32-34]。9月末油蒿葉片老化進入枯萎期,葉綠素含量、各種酶活性伴隨光合單元機構衰退降低,低溫脅迫導致Fv/Fm低值出現[10,38]。
在季節(jié)尺度上,油蒿PSII能量分配滯后Ta和VPD相對PAR和SWC較短。說明在長時間尺度,油蒿PSII對Ta和VPD更為敏感。植物在高VPD下會調節(jié)氣孔關閉[13,22],增強葉黃素循環(huán)等持續(xù)性熱耗散[23],同時對PSII反應中心造成可逆損害[32-34]。極端溫度或高太陽輻射則會打破PSII能量輸入和利用的平衡,造成PSII捕光蛋白損傷[39]。與其他研究比較,油蒿對極端溫度的適應能力要高于荒漠物種沙柳和紅砂(Reaumuria soongorica)[8,36]。荒漠地區(qū)晝夜溫差大,油蒿可通過提高細胞內滲透壓,增加原生質抗凝結性,減緩代謝速率來適應極端溫度[39]。油蒿PSII能量分配滯后PAR約14.7 d(表 2),油蒿對于高輻射具有一定的耐受能力[16,34]。干旱是影響植物生長生存的重要環(huán)境脅迫,水分限制著細胞內一系列的生理生化反應[13,24]。油蒿具有粗長發(fā)達的主根和茂密的側根[17,40],PSII能量分配滯后SWC時間最長,約21.6 d(表2),可見其對干旱有著極強的耐受性,因此也經常用作干旱區(qū)植被恢復的先鋒物種。
4 結論
1)日尺度上,油蒿可能通過葉黃素循環(huán)等熱耗散機制和氣孔導度調節(jié)PSII能量分配,主要受光合有效輻射(Photosynthetically Active Radiation,PAR)控制,同時受空氣溫度(Ta)和飽和水汽壓差(Vapor Pressure Difference,VPD)的調控,對土壤含水率(Soil Water Content ,SWC)敏感性較差。PSII能量分配參數滯后于PAR 43 min,且分別滯后于Ta、VPD和SWC 3.3、4.6和10.72 h。
2)季節(jié)尺度上,油蒿可能通過提高PSII修復能力、增加特定蛋白數量和色素濃度等方式調節(jié)PSII能量分配,對Ta和VPD變化較敏感,并且受物候期的影響。PSII能量分配參數分別滯后于Ta、VPD、PAR和SWC 7.2、8.8、14.7和21.6 d。
3)在荒漠惡劣環(huán)境下,油蒿可以通過自身調節(jié)不斷修復受損PSII(Fv/Fm維持在0.78附近),對干旱和高輻射表現出耐受性,具有一定的適應能力和生理可塑性。
猜你喜歡
中老年保健(2021年12期)2021-08-24 03:30:40
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
鐵道通信信號(2020年9期)2020-02-06 09:15:22
中國生殖健康(2020年6期)2020-02-01 06:28:50
數學大王·趣味邏輯(2019年5期)2019-06-13 20:27:43
小學科學(學生版)(2019年5期)2019-05-21 01:00:18
中國生殖健康(2019年11期)2019-01-07 01:28:02
經濟技術協(xié)作信息(2018年30期)2018-11-22 06:20:24
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
