養殖模式對草魚池塘底泥微生物群落結構影響的分析
2021-04-07 00:49:14章海鑫余智杰張燕萍張愛芳付輝云
淡水漁業 2021年2期
關鍵詞:結構
章海鑫,余智杰,張燕萍,張愛芳,王 生,李 涵,付輝云
(江西省水產科學研究所,南昌 330039)
草魚(Ctenopharyngodonidellus)是我國淡水養殖面積最大、產量最高的品種,產量占淡水養殖總產量的18.4%[1]。水產養殖(特別是草魚養殖)是為國民提供優質蛋白的主要方式之一,但養殖過程中存在著因養殖密度大而導致水體富營養化、病害增多、品質低下等問題。因此,發展既能保證產量又能提高產品質量,減少環境污染的養殖技術是草魚養殖面臨的迫切問題,而生態養殖是解決這一問題的重要發展方向。
水產養殖生態系統是由養殖動物、水體、底泥等因素構成的相互影響、相互作用的綜合生態系統。其中,底泥中的微生物對系統中有機質、無機鹽等物質循環代謝的強度直接影響水體富營養化水平[2]和水體理化環境[3];水體中水生植物對微生物分解形成的無機鹽的吸收和利用又反過來影響著微生物的群落結構和養殖動物的生長;養殖動物所產生的營養物質對于微生物的結構組成又有一定選擇作用。所以,底泥微生物在整個養殖系統中扮演著重要的角色,它不僅參與養殖系統中有機質、無機鹽等物質的循環代謝及能量代謝[4],本身又受到水體環境因子、水體植物和養殖動物的影響。
許多研究者認為通過提高養殖系統微生物的分解能力,平衡好水生植物和浮游植物對無機鹽的吸收和其為養殖動物提供餌料的關系是生態養殖發展的方向[5],也是解決養殖過程中高產量與污染相矛盾的重要手段[6]。在不同的養殖方式下,養殖池塘底泥、水體和養殖動物腸道中的微生物均有所不同,從而表現出多樣性,微生物群落結構的多樣性同樣會影響著不同養殖條件下水體理化環境因子和養殖效果。因此,從底泥微生物群落結構角度出發,探討分析不同養殖方式底泥微生物群落結構的特點及其與水體的理化環境的關系可以為開發生態養殖技術提供理論支持,也能夠為探討不同養殖模式之間的優劣提供一個依據,具有重要生態和經濟意義。目前,草魚池塘養殖中常見的模式有魚菜共生模式[6-7]、山塘模式[8]和傳統精養模式,本實驗比較了具有代表性的兩種草魚生態養殖模式——魚菜共生模式和山塘模式與傳統精養模式條件下池塘底泥的微生物群落結構的四季變化,并分析微生物群落結構與水體環境因子的關系,旨在為評估和判斷草魚養殖環境的變化情況,發展草魚生態養殖提供理論支持。
1 材料與方法
1.1 養殖模式構建
在江西省南昌縣蔣巷鎮三洞村選取一個面積為30 000 m2的草魚養殖塘,改造成3個面積均為10 000 m2,深度為2.5 m的標準養殖塘。選取其中一個塘采用山塘模式養殖(投喂草,投喂率10%),另一個塘采用魚菜共生模式養殖(使用PVC管構建浮床,種植水空心菜,種植面積占水面10%,投喂配合飼料,投喂率3%~4%),剩余一個塘采用傳統精養模式(投喂配合飼料,投喂率3%~4%)。其中山塘放養草魚3 740尾/ 10 000 m2,規格為16~24尾/kg,精養池塘和魚菜共生池塘均放養草魚12 000尾/1 0000 m2,規格為16~24尾/kg。放養時間均在2018年12月,水深常年保持在2~2.2 m。
1.2 樣品采集與預處理
1.1.1 底泥樣品采集與預處理
三種養殖模式條件下樣品的編號情況見表1。每次采樣用彼得森采泥器在池塘四個角離岸邊25 m處和池塘中點采集表層底泥樣品各50 g,然后混勻成一個樣并編號,每個池塘按季度采四次。采集的底泥暫時保存于干冰采樣箱中,24 h內帶回實驗室,凍存于-80 ℃冰箱中。
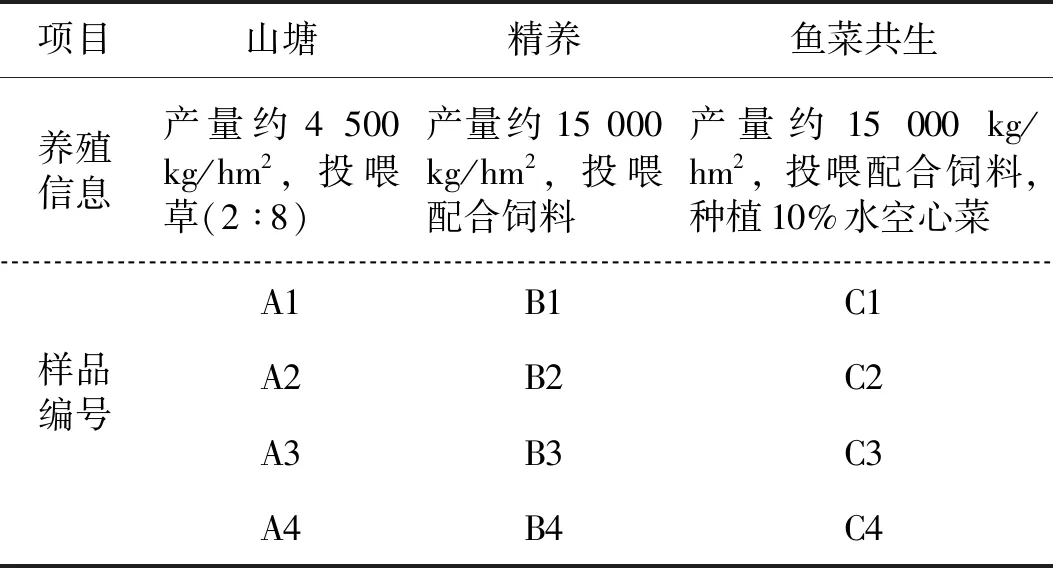
表1 養殖信息及樣品編號Tab.1 Information of culture models and sample number
1.1.2 水樣采集與處理

1.2 底泥樣品DNA 提取、擴增和測序
將每季度混合后的底泥樣品充分混合后稱取2 g用土壤 DNA 提取試劑盒(OMEGA Soil DNA Kit,D5625)提取。16S rRNA基因V3-V4區擴增引物為:341F(5′-CCTACGGRRBGCASCAGKVRVGAAT-3′) 和 805R(5′-GGACTACNVGGGTWTCTAATCC-3′)。PCR產物使用2%濃度的瓊脂糖凝膠進行檢測后,利用凝膠回收試劑盒(QIAGEN)對目的條帶進行回收。然后使用TruSeq?DNA PCR-Free Sample Preparation Kit 建庫試劑盒(Illumina)進行文庫構建,構建好的文庫經過 Qubit 和 Q-PCR 定量,使用 HiSeq2500(諾禾致源公司,北京)進行測序。
1.3 數據處理與分析
1.3.1 序列處理
參照陳瓊等[9]的方法采用Miseq測序平臺得到原始數據(raw data),然后進行拼接和質控,得到有效數據(clean data),再進行嵌合體過濾,得到可用于后續分析的有效數據(effective tags)。 然后基于有效數據進行 OTUs(operational taxonomic units)聚類。
1.3.2 統計分析
水質數據采用Excel 2010處理數據,用SPSS19.0進行方差分析和相關性分析。
序列數據利用 Uparse 軟件對所有樣品的全部effective tags進行聚類,以97% 的一致性將序列聚類成OTUs。根據OTUs聚類結果對每個OTU的代表序列做物種注釋,得到對應的物種信息,根據物種注釋結果,選取每個樣品在門和屬2個分類級別上最大豐度排名前10的生成相對豐度柱形累加圖。對所有樣品的OTUs進行豐度分析、Alpha多樣性分析等。使用Qiime 軟件(Version 1.7.0)計算Alpha多樣性指數,包括豐富度指數(Richness)、ACE 指數(ACE Index)和Chao1 指數(Chaos1 Index)。不同模式間進行UPGMA(unweighted pair-group method with arithmetic means)聚類分析和Anosim(analysis of similarities)分析。
2 結果
2.1 水體環境因子

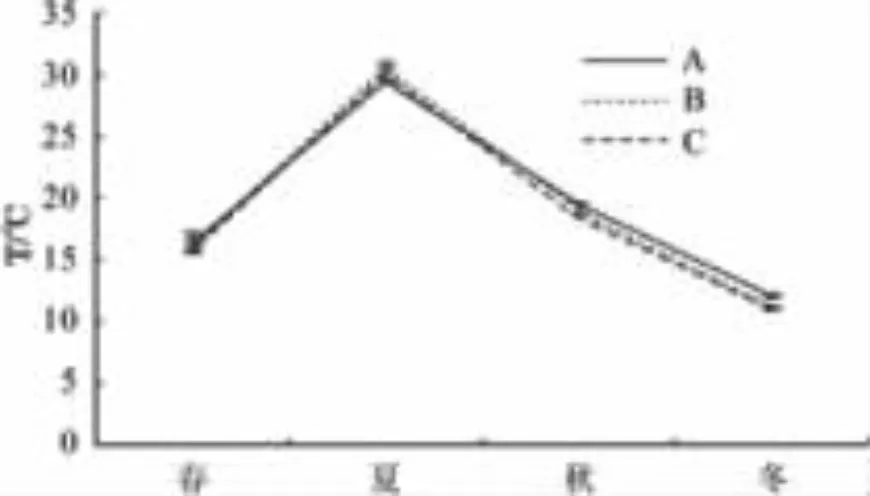
圖1 水溫變化情況Fig.1 Changes of water temperature in four seasons
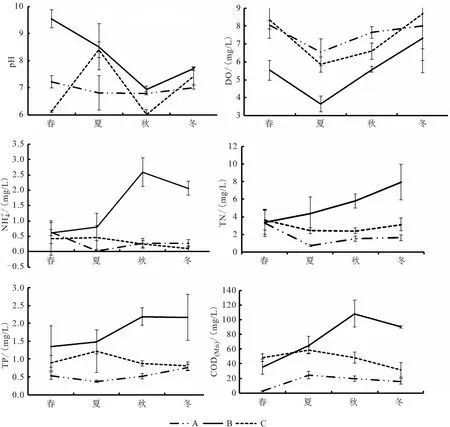
圖2 pH、溶解氧、氨氮、總氮、總磷和CODMn四季變化情況Fig.2 Changes of and CODMn in four seasons
2.2 高通量測序數據
測序得到的序列經過拼接和質控,然后過濾掉嵌合體后得到Effective Tags條數為54 775~68 965。去除無法聚類到OTUs和沒有注釋的序列條數后得到可構建OTUs的序列(taxon tags)條數為48 067~63 071(表2)。以97%的一致性,將每個樣品所得的序列聚類成 OTUs(表2)。
2.3 微生物群落結構
2.3.1 物種相對豐度
樣品中相對豐度最高的10個OTUs在門水平上分布見圖3,可以看到三種養殖模式條件下四季底泥樣品中豐度最高的均是變形菌門(Proteobacteria)。但A組變形菌門占比四季是下降趨勢;其余占比超過10%的門是擬桿菌門(Bacteroidetes)和酸桿菌門(Acidobacteria),其中酸桿菌門四季占比逐漸升高。B組變形菌門占比先降后上升;擬桿菌門和酸桿菌門占比均超過10%,而且擬桿菌門占比逐漸下降和酸桿菌門占比逐漸上升;綠彎菌門(Chloroflexi)占比也超過10%。C組除了變形菌門外,擬桿菌門占比也超過10%;但綠彎菌門、酸桿菌門、硝棘菌門(Nitrospinae)、螺旋體門(Spirochaetes)等占比均比較高;而且每個門占比四季存在一定的波動。
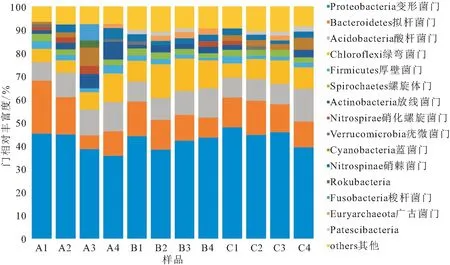
圖3 樣品在門水平上物種相對豐度Fig.3 Relative abundance of samples at phylum level
2.3.2 核心群落相對豐度
樣品在屬水平上相對豐度最高的10個OTUs為核心群落,可以反映群落物種的基本結構情況。從圖3和表3中可以明顯看到三組中沒有鑒定的種類在四季中占比均最高,有未培養(uncultured)、未培養細菌(uncultured-bacterium)、未培養土壤細菌(uncultured-soil-bacterium)和未培養原核生物(uncultured-prokaryote)四種。其中地桿菌屬(Geobacter)和厭氧粘細菌屬(Anaeromyxobacter)在三組中四季均有,其余種屬A組與B組和C組相差較大,而且A組的種屬四季差異較大。B組和C組重要種屬相似,且四季差異較小。具體詳見表3。
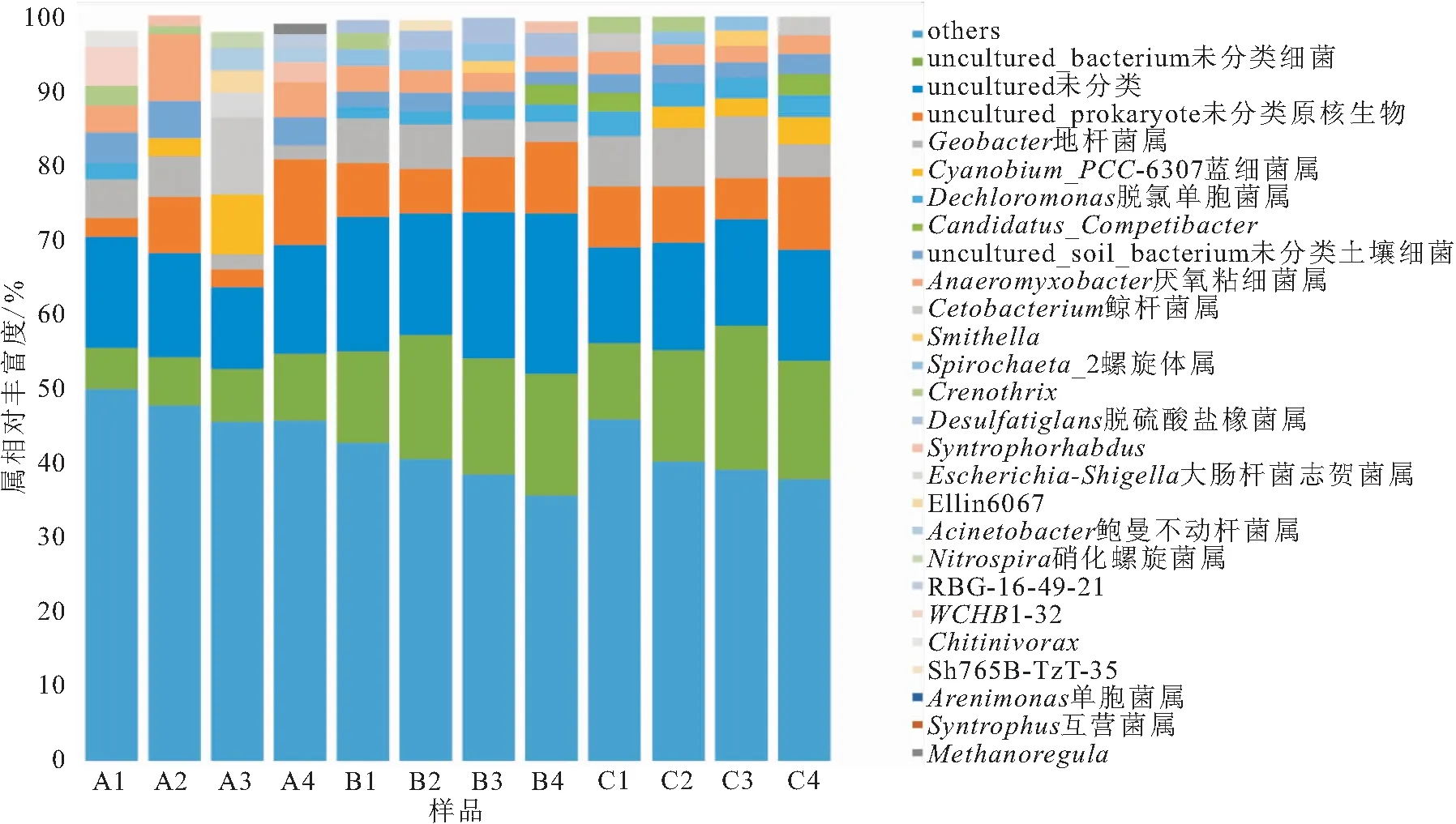
圖4 樣品在屬水平上相對豐度Fig.4 Relative abundance of samples at genus level
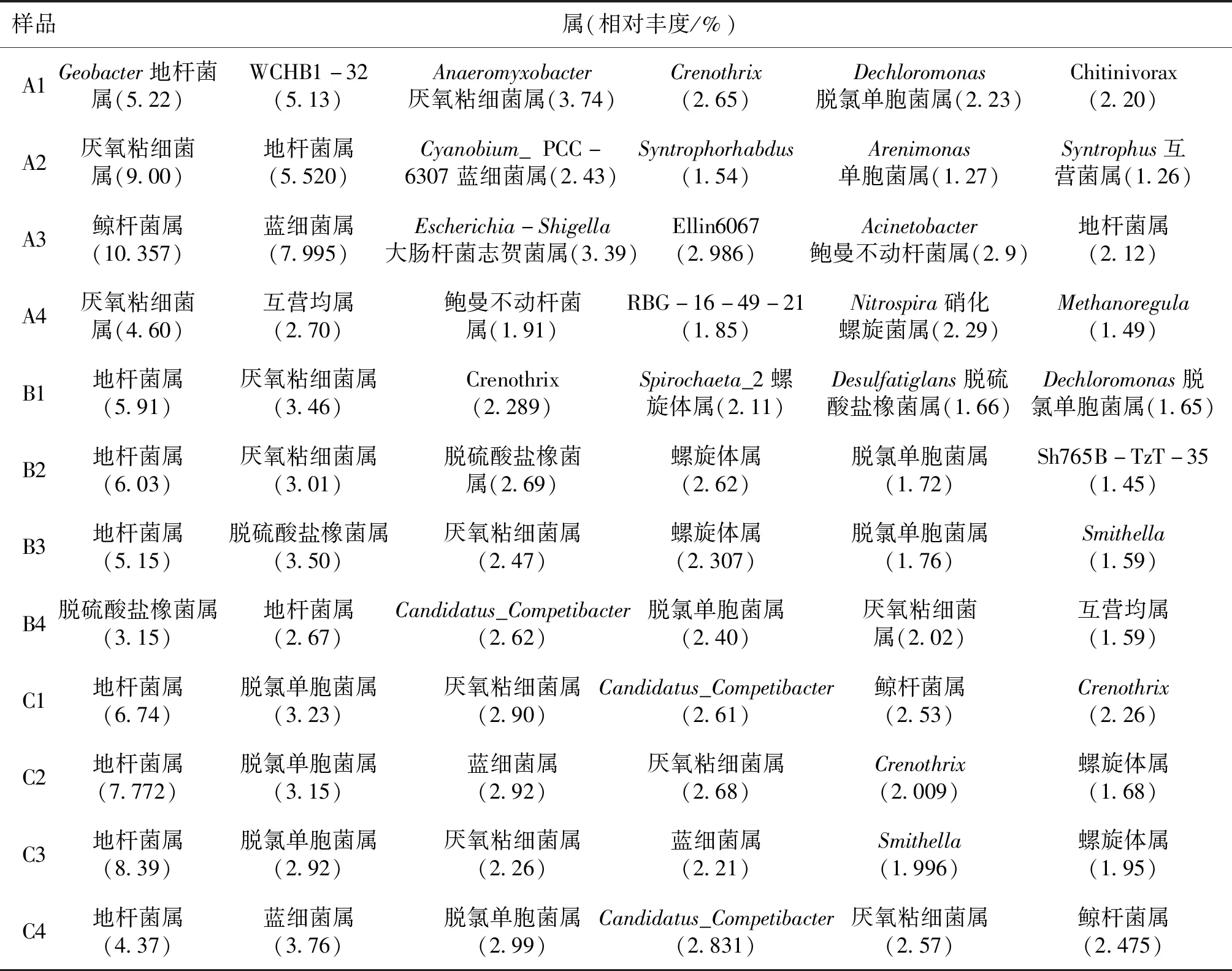
表3 在屬水平豐度最高的10個OTUSTab.3 Top ten OTUs with the highest abundance in genus level
2.3.3 微生物群落結構聚類分析
UPGMA聚類分析結果(圖5)可知,各樣品在時間上的相似性(不同季度)的相似性高于空間上(不同養殖方式)。具體表現為B組和C組相似性接近,而A組與B組和C組相似性低;B組和C組均是夏季和秋季相似度最高,A組是春夏兩季相似度高。
對A、B、C三組樣品均作了Anosim差異性分析,從圖6可以看到A組與C組的組間差異大于A組和B組以及B組和C組,組間差異最小為B組和C組。A、B、C三組的組間差異均顯著高于組內差異。
2.4 多樣性分析
三組多樣性方差分析結果見圖7,從圖7可以看到A組ACE指數顯著低于B組和C組,B組和C組間差異不顯著,但Shannon指數三組差異不顯著。從表4中可以看到三種養殖模式條件下豐富度指數ACE各季度差異也大。其中A組四季豐富度指數變化差異大,其次是B組,C組豐富度指數四季最穩定;除A組外,B組和C組豐富度均是先增后降的趨勢。Chao1指數的變化情況與ACE類似。多樣性指數Shannon A組冬季多樣性高于B組和C組,但夏季和秋季低于B組和C組,C組除了冬季外其他時間多樣性均小于B組;各組內四季也存在一定的變化;A組除了秋季外,其余三個季度均比較穩定; B組和C組類似,均是夏季> 春季>秋季>冬季。
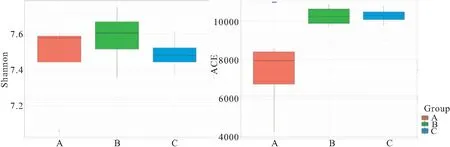
圖7 ACE指數和Shannon指數差異分析圖Fig.7 Differentiation analysis of richness(ACE) and Shannon indices in groups***表示差異極顯著
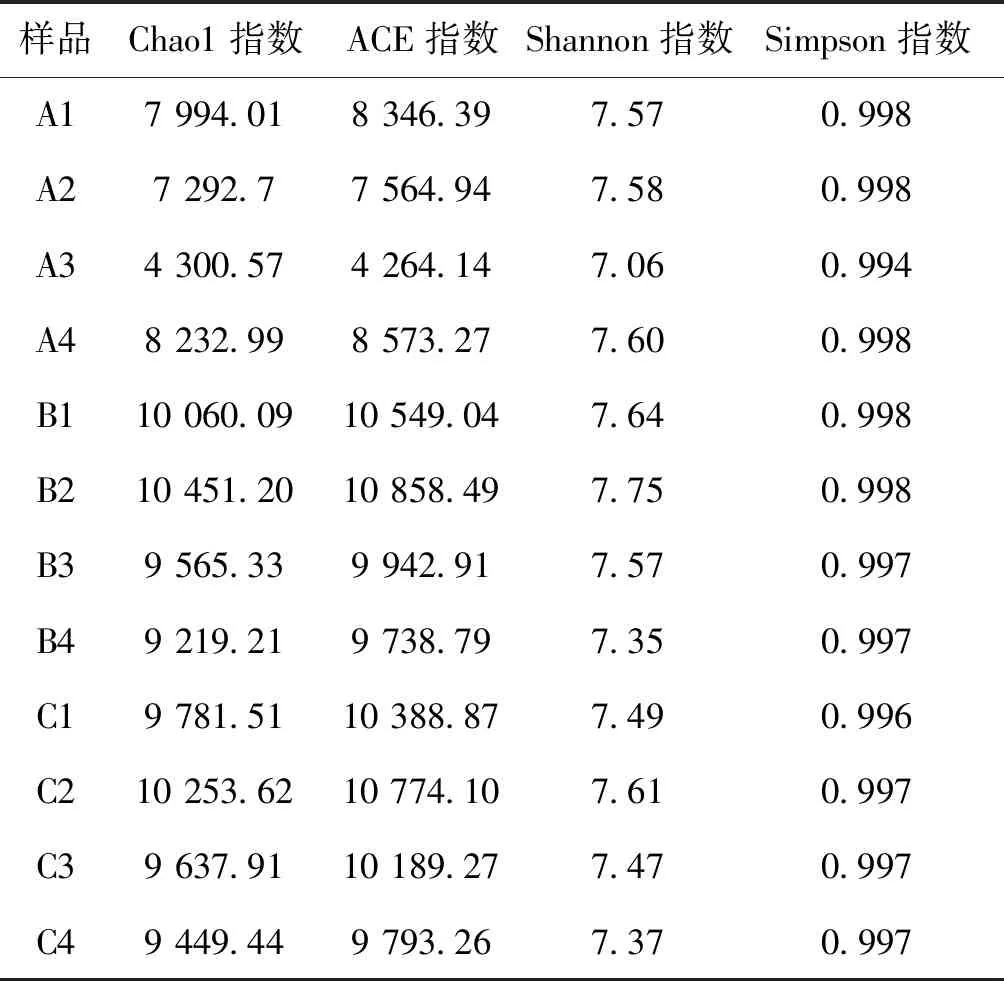
表4 基于OTUs的多樣性指數的 Alpha 多樣性分析Tab.4 Alpha diversity of bacteria samples associated with the number of observed OTUs
2.5 環境因子與微生物群落多樣性相關性分析
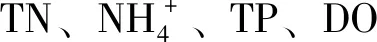
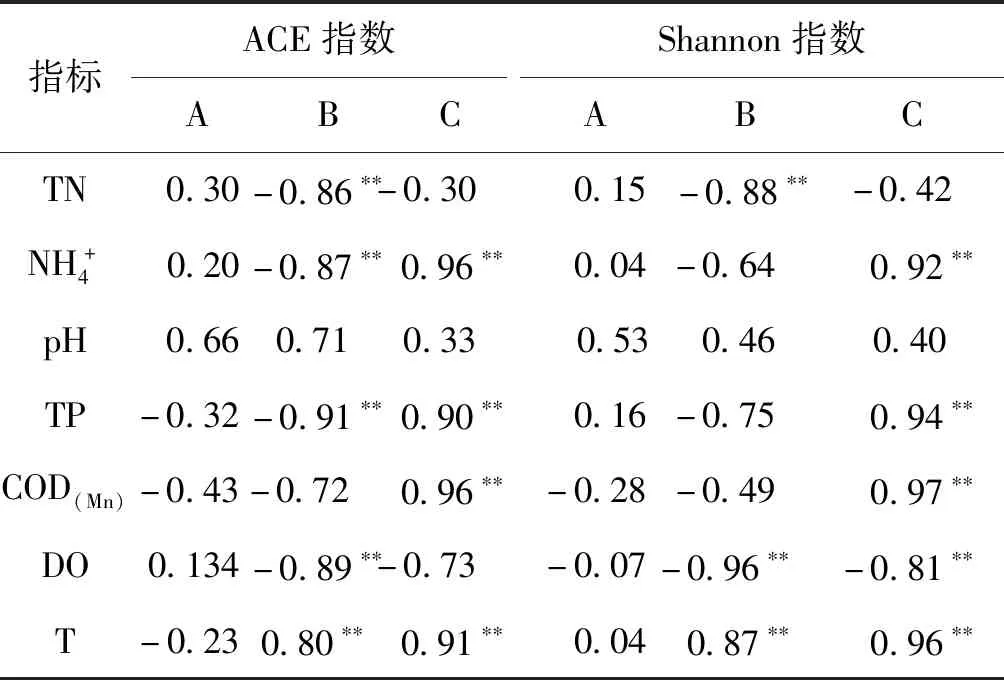
表5 環境因子與多樣性相關性分析Tab.5 Correlation analysis between environmental factors and the microbial community diversity
3 討論
3.1 三種養殖模式條件下底泥微生物群落結構多樣性分析
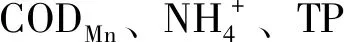
3.2 三種養殖模式條件下底泥微生物核心群落
底泥微生物核心菌群的種類和分布情況可以一定程度上反應底泥微生態情況,比較三種養殖模式各季度微生物群落在屬水平的分布情況可以發現,A組主要屬在豐度上和種類上四季均有很大差別。春夏季以進行還原作用的厭氧或兼性厭氧菌屬的地桿菌屬、WCHB1-32屬、厭氧粘細菌屬、Crenothrix屬和脫氯單胞菌屬等[11、13-18]為主。而秋季則出現了進行亞消化作用的硝化螺旋菌屬[19-21]和能夠在好氧條件下進行固氮,厭氧條件下還原硫的藍細菌屬[22]。同時鯨桿菌屬、大腸桿菌志賀菌屬、鮑曼不動桿菌屬的豐度占比也進入了前10。其中鯨桿菌屬的種類通常寄生于水產動物腸道,參與促進和合成蛋白質及碳水化合物和脂肪的代謝[23、24];大腸桿菌志賀菌屬和鮑曼不動桿菌屬均是環境中的條件致病菌,而鮑曼不動桿菌屬中菌能夠引起水產動物疾病[25-26]。冬季則是以厭氧粘細菌屬、Syntrophorhabdus、地桿菌屬等和Methanoregula等占主要地位。說明A組底泥微生物核心菌群結構穩定性差。與之相比,B組雖然主要種屬的豐富度在四季存在一定的變化,但種類變化較小,主要以地桿菌屬、厭氧粘細菌屬、Crenothrix、螺旋體屬、脫硫酸鹽橡菌屬等為主。其中的地桿菌屬[11、13]、厭氧粘細菌屬[15]、Crenothrix屬[18]、脫硫酸鹽橡菌屬[27]均屬于厭氧或兼性厭氧菌,在厭氧條件下還原有機物。說明B組底部溶氧不足和有機物沉積過量,因為有機物的積累能刺激地桿菌屬和厭氧年細菌屬等菌屬的大量富集[14]。對水體理化檢測的數據也支持這一結論(DO最低、COD(Mn)及三氮均最高)。C組春季豐度較高的是地桿菌屬、脫氯單胞菌屬、厭氧粘細菌屬和Candidatus-Competibacter屬等還原有機物,脫硫及除磷作用的菌屬,到夏季開始出現藍細菌屬和Crenothrix,且豐富度上升,Candidatus-Competibacter屬豐富度也上升。因為藍細菌屬具有能夠在好氧條件下進行固氮,厭氧條件下還原硫的作用[22],Crenothrix[18]具有硝化作用,所以C組池塘夏季底泥中除磷、固氮和硝化菌種類和數量均升高,促使水體COD(Mn)、總磷、總氮和氨氮等理化指標均呈現下降的趨勢。同時鯨桿菌屬也是C組系統中的主要菌屬,鯨桿菌屬具有參與促進和合成蛋白質及碳水化合物和脂肪的代謝的作用[23、24],是對養殖水產動物有益的腸道菌群。所以,從核心菌群功能結構、微生物的多樣性關系,以及參照水體理化指標來分析,均可以表明魚菜共生系統底泥菌群結構更為優化。
3.2 水體理化環境對底泥微生物群落結構的影響

4 結論
山塘模式雖然水體DO相對較高,水體理化環境最好(TN、TP等最低),但菌群結構不穩定、四季波動大,加上養殖密度低、產量也低,也存在能引起魚類病害的條件致病菌的隱患,所以其沒有達到充分提高養殖效益的目的。魚菜共生模式雖然產量與精養模式一樣,但水體有機物低于精養模式,水生植物能夠吸收無機鹽類,提供溶氧,而且底泥微生物群落結構穩定,水環境因子能夠促進底泥微生物的生長和繁殖,能夠穩定持續的凈化水體環境,是一種能夠保證產量且提高養殖效益的養殖方式。
猜你喜歡
小獼猴智力畫刊(2023年4期)2023-04-23 08:49:58
哲學評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
中學生數理化·高一版(2018年1期)2018-02-10 05:20:03
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
七彩語文·寫字與書法(2016年7期)2016-07-28 21:40:22
七彩語文·寫字與書法(2016年6期)2016-07-15 19:36:34
人間(2015年21期)2015-03-11 15:23:21
現代企業(2015年9期)2015-02-28 18:56:50
