生物信息學分析miR-221/26a、lncRNA GAS5和RECK mRNA在結直腸癌轉移中的作用*
2021-04-13 07:10:16孟濤劉林王海江
貴州醫科大學學報 2021年3期
孟濤,劉林,王海江
(新疆醫科大學附屬腫瘤醫院 胃腸外科,新疆 烏魯木齊 830011)
結直腸癌(colorectal cancer)是世界范圍內最常見的惡性腫瘤之一,其發病率和死亡率均位居惡性腫瘤的前列[1]。約有40%結直腸癌患者最終死于復發和轉移[1-5],且其發生機制仍不明確。因此,研究結直腸癌轉移的發生機制具有重要的意義。回復引導半胱氨酸豐富蛋白Kazal基元(reversion inducing cysteine rich protein with kazal motifs,RECK)是一個公認的腫瘤轉移抑制因子,研究顯示RECK與結直腸癌轉移有密切的關系[2]。MicroRNA(miRNA)是一類長度在19~24個核苷酸的內源性非編碼小分子單鏈 RNA,能通過與靶基因 miRNA 特異性的堿基互補配對,引起靶基因 miRNA 的降解或者抑制其翻譯,負調控靶基因的表達,參與和影響細胞分化、組織發育、腫瘤診斷及預后[3];而長編碼 RNA(long non-coding RNA,lncRNA)是一類長度超過 200個核苷酸、不編碼蛋白的RNA,在發育、分化及代謝等多方面發揮著重要作用[4]。miRNA和lncRNA都高度保守穩定,可競爭性結合并調控靶基因參與腫瘤的發生發展,因而可作為潛在標志物用于腫瘤的預測、診斷及預后的判斷[5-8]。本研究通過生物信息學方法找出結直腸癌轉移相關的miRNA及lncRNA,并選取結直腸癌變組織及其對應的癌旁觀察其表達,分析它們的相互作用及對結直腸癌轉移的影響。
1 材料與方法
1.1 試劑及儀器
Trizol 購自上海生工公司,miRcute miRNA cDNA 試劑盒(KR201)、miRcute增強型miRNA熒光定量檢測試劑盒(SYBR Green,FP411)及lnRcute lncRNA熒光定量檢測試劑盒(SYBR Green) (FP402) 購自北京天根生化科技有限公司,lncGAS5過表達質粒及含lncGAS5啟動子的CRISPR-Cas9質粒購自吉凱基因公司,轉染試劑Lipofectamine 3000購自Thermo Fisher,DMEM等細胞培養試劑購自Gibco公司;Anti-RECK抗體(ab238162)和山羊抗兔IgG H&L (HRP,ab205718)購自Abcam,所用引物U6、RECK及 miRNA 26a購自廣州復能公司。
1.2 方法
1.2.1生物信息學預測 利用miRanda數據庫(http://www.microrna.org/microrna/home.do)和 Starbase數據庫(http://starbase.sysu.edu.cn/)預測RECKmRNA-miRNA,miRNA-lncRNA的結合位點并做分析。
1.2.2構建過表達和沉默lncGAS5的SW480細胞 在轉染的前一天,傳代準備SW480細胞,待細胞密度生長至60%~80%時即可用于質粒轉染。將2種質粒分別與轉染試劑Lipofectamine 3000(先用不含血清及雙抗的DMEM培養基稀釋)混勻,加入到SW480細胞中;于37 ℃培養箱培養6~8 h后,取出轉染細胞,用2 mL新鮮的不含雙抗的完全培養基換掉原培養基,轉染48 h后、加入嘌呤霉素(3.0 mg/L)進行篩選。10~15 d后收取細胞提取RNA和蛋白。
1.2.3RNA、miRNA、lncRNA提取 取手術切除的結直腸癌患者結直腸癌變組織及癌旁 5 cm 以上的正常黏膜組織,用Trizol提取總RNA,用試劑盒提取miRNA和lncRNA,紫外分光光度儀測定濃度及純度(OD260/OD280>1.8可用)。按照試劑盒說明書對miRNA和lncRNA進行熒光定量檢測。
1.2.4熒光定量PCR(qPCR) 吸取1.2.3中得到的RNA,加入超純水,70 ℃孵育5 min,冷卻后置于冰上,加入反轉錄酶,42 ℃孵育60 min,70 ℃加熱10 min,保存于-20 ℃。基因引物由上海生工生物有限公司合成:lncRNA GAS5,F為5-CTTCTGGGCTCAAGTGATCCT-3,R為5-TTGTGCCATGAG
ACTCCATCAG-3;GAPDH,F為5-ACCAGCCCCAGCAAGAGCACAAG-3,R為5-TTTGCTTGAAGTTTCA
CTGGCATC-3;miR-221,F為5-AGCTAAAAAAGCTACATTGTCTGCTGGGTTTCG-3,R為5-GATCCGA
AACCC AGCAGACAATGTAGCTTTTTT-3)。反應條件為95 ℃ 2 min,95 ℃ 30 s、60 ℃ 30 s、72 ℃ 5 min,32個循環。實驗重復3次,采用2-ΔΔCt法計算基因的相對表達量。
1.2.5ECK蛋白表達水平 采用蛋白質印跡法(Western blot),將少量組織或構建過表達和沉默lncGAS5的SW480細胞置于勻漿器中,加入蛋白質裂解液研磨至無肉眼可見組織置于冰上裂解30 min,將裂解液轉移至離心管中,4 ℃ 12 000 r/min離心5 min,取上清,在紫外分光光度儀上測定蛋白質濃度并置-20度保存。制膠后,加入50 ng變性蛋白溶液進行電泳及蛋白轉移,結束后加入RECK一抗和二抗進行孵育,最后進行凝膠圖像分析。
1.3 統計學方法
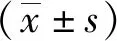
2 結果
2.1 預測RECK/miRNA/lncRNA結合位點及組織標本中含量測定
利用miRanda數據庫預測結果顯示(表1),RECKmRNA與miR-221有4個結合位點,與miR-26a有1個結合位點,推測miR-221/26a能靶向結合到RECKmRNA上對其進行調控。利用Starbase數據庫預測結果顯示lncRNA GAS5與miR-221/26a存在1個結合位點,提示它們之間存在相互作用關系。因此,利用qPCR對結直腸癌組織和癌旁組織中lncRNA及miR-221/26a的含量進行驗證(圖1),結果顯示在結直腸癌組織中,miR-221/26a的表達高于其相應的癌旁組織 (P<0.001、或0.05),lncRNA GAS5 在結直腸癌中表達水平降低,差異具有統計學意義(P<0.001)。

表1 miRanda數據庫預測RECK mRNA與miR-221/26a結合位點Tab.1 Predicted binding sites of RECK mRNA with miR-221/26a using miRanda database

表2 Starbase數據庫預測lncRNA GAS5與miR-221/26a結合位點Tab.2 Predicted binding sites of lncRNA GAS5 with miR-221/26a using Starbase database

圖1 lncRNA GAS5及miR-221/26a在結直腸癌組織及癌旁組織中的表達Fig.1 The relative expression levels of lncRNA,miR-221,and miR-26a in CRC tissues and corresponding adjacent tissues
2.2 過表達lncRNA GAS5的SW480細胞中lncRNA GAS5及miR-221/26a表達
取穩轉lncRNA GAS5后的SW480細胞提取RNA和蛋白,結果如圖2所示。結果顯示,過表達組SW480細胞中GAS5相對含量為0.137±0.045,轉染空載組含量為 0.031±0.015,過表達組GAS5的含量是空載組的4.4倍,2組比較差異有統計學意義(P<0.05)。過表達組SW480細胞中的miR221相對含量為0.004±0.001,空載組為0.010±0.002,過表達組低于空載組,僅為空載組的0.4倍,差異有統計學意義(P<0.01)。過表達組SW480細胞中的miR26a相對含量為0.005 8±0.002 3,空載組為0.012±0.001,過表達組低于空載組,僅為空載組的0.48倍,差異有統計學意義(P<0.01)。Western blot結果顯示,過表達組細胞中的RECK蛋白表達含量低于空載組。

注:與空載組比較,(1)P<0.01,(2)P<0.01。圖2 穩轉lncRNA GAS5后SW480細胞中lncRNA GAS5及miR-221/26a表達Fig.2 The expression levels of lncRNA,miR-221,and miR-26a in SW480 cells overexpressing lncRNA GAS5
2.3 lncGAS5 沉默細胞株情況
將質粒轉染到SW480細胞后,用qPCR檢測lncGAS5、miR221及miR26a含量,結果如圖3所示。結果顯示,GAS5在沉默組SW480細胞中相對含量約為0.000 5±0.000 13,而在空載組則為0.032±0.005 6,沉默組低于空載組,差異有統計學意義(P<0.01),提示成功建立lncGAS5 沉默細胞株。miR221在沉默組相對表達含量為0.078±0.021,空載組中為0.03±0.02,沉默組高于空載組,差異有統計學意義(P<0.01)。miR26a在沉默組相對表達含量為0.059±0.007 3,空載組中為0.039±0.011,沉默組高于空載組,差異有統計學意義(P<0.01)。提示GAS5的降低在一定程度上會上調miR221和miR26a的含量。Western blot 結果顯示,沉默GAS5時,RECK蛋白表達增加。
3 討論
我國結直腸癌的發病率和死亡率均呈上升趨勢,其發病率、死亡率在所有惡性腫瘤中均居第5位[6]。因此,尋找和發現結直腸癌相關的癌基因并揭示和預測其在結直腸癌的病程進展和轉移過程中的功能,對于結直腸癌的診治具有十分重要的理論意義和實際應用價值。
RECK是一個公認的腫瘤轉移抑制因子,其蛋白主要定位于細胞外。它富含半胱氨酸,是一類膜錨定糖蛋白,在常見細胞中通過轉錄后能調控多種基質金屬蛋白酶(MMPs)的表達,阻遏腫瘤血管生成,從而抑制腫瘤的侵襲與轉移[7]。有研究發現,在結直腸癌病人的腫瘤樣本中,RECK的表達水平和病人復發風險有關,高濃度的RECK往往預示著病人的復發風險較低。此外通過動物及細胞實驗發現,提升RECK的表達能抑制腫瘤細胞的侵襲、轉移及血管新生[8-9]。以上研究提示RECK與結直腸癌轉移有密切的關系。課題組在前期研究中探討了RECK蛋白和mRNA在原發腫瘤組、結直腸轉移灶組以及癌旁組織的表達情況,證實了RECK蛋白及mRNA水平在腫瘤組中是呈低表達的,且在轉移灶組中RECK蛋白及mRNA表達量更低。

注:(1)與空載組比較,P<0.01。圖3 沉默lncRNA GAS5的SW480細胞的qPCR及Western blot 結果Fig.3 The results of qPCR and Western blot in SW480 cells silencing lncRNA GAS5
miRNA是一類約含22個核苷酸的非編碼小分子單鏈RNA,通過與靶基因mRNA的3’端非翻譯區上的相應靶點結合,能抑制或降解特定靶基因,從而起到調節蛋白質合成的作用[10-11]。有研究發現miR-25-3p通過調節內皮細胞相關因子表達,可以促進血管通透性和血管生成,同時大腸癌細胞的miR-25-3p外顯子可顯著誘導大腸癌的血管滲漏,促進大腸癌在肝、肺的轉移。此外,miR-25-3p的表達水平在發生轉移的大腸癌患者中明顯高于無轉移者,證明了miR-25-3p是一種促進結直腸癌轉移的miRNA[12]。有學者發現與直腸癌相關的miRNA約有50多個,這些miRNA可通過改變Wnt通路的β-連環蛋白(βcatenin)、表皮生長因子受體(EGFR)、轉化生長因子-β(TGF-β)和腫瘤抑制蛋白(TP53)等信號通路來影響大腸癌的生存、增殖、侵襲和轉移,因此,miRNA與結直腸癌的發生發展密不可分[14-15],而lncRNA與miRNA一樣,屬于非編碼RNA,區別是lncRNA轉錄本長度超過200個核苷酸,在生物發育、分化和代謝等方面起到了重要調控作用,成為近年來的研究熱點[16-18]。不少研究顯示lncRNA和miRNA存在競爭結合參與腫瘤細胞的侵襲和轉移[19-21]。如Wang等[22]人在綜述中就提到40多種lncRNA/miRNA參與結直腸癌發生發展的情況。
本研究利用生物信息學進行預測,發現了多條與RECKmRNA相結合的miRNA,其中 miR-221 與之有4個結合位點,miR-26a 與之有1個結合位點,由此推測RECK表達與這2個miRNA有關系,進而對結直腸癌病人的腫瘤組織和癌旁組織中miR-221/26a進行驗證。結果顯示這2個miRNA在結直腸癌腫瘤組織中高表達。這或許暗示了在結直腸癌中,上調的miR-221/miR-26a 通過結合RECKmRNA,抑制了RECK蛋白的表達,從而促進結直腸癌的轉移。同時數據庫分析結果還顯示lncRNA GAS5能同時結合 miR-221、miR-26a,本研究也對結直腸癌病人的腫瘤組織和癌旁組織中lncRNA GAS5進行驗證,結果顯示lncRNA GAS5在結直腸癌腫瘤組織中呈低表達。
以上結果初步提示在結直腸癌細胞中,lncRNA GAS5表達降低,會使得 miR-221 和miR-26a 的豐度升高,從而抑制了RECKmRNA 表達,最終促進結直腸癌細胞轉移。基于以上推測,本研究在SW480細胞中過表達了lncRNA GAS5,通過測定GAS5含量提示我們過表達細胞模型構建成功且miR-221/26a含量隨著GAS5的增加而下降,RECK蛋白含量降低,提示GAS5能通過負反饋miR-221/26a含量影響RECK蛋白表達。同時,本研究還通過轉染含GAS5啟動子的CRISPR-Cas9質粒,下調其含量,并檢測miR-221/26a變化,結果發現隨著GAS5含量下降,miR-221/26a和RECK表達均上調。通過細胞實驗可以初步證實lncRNA GAS5表達降低,對 miR-221 和 miR-26a 的吸附作用降低,導致 miR-221 和miR-26a 的表達升高,結果提示miR-221 和 miR-26a 能與抑癌基因RECKmRNA結合,并抑制其表達,進而最終促進結直腸癌細胞轉移。
綜上所述,本研究先通過生物信息學預測出相關的lncRNA和miRNA,再通過構建過表達和沉默細胞株驗證GAS5,miR-221/ 26a和RECK之間的關系,結果表明lncRNA GAS5和miR-221 /miR-26a與結直腸癌發展密切相關,檢測其含量變化可提前為癌細胞是否轉移做出預測。
