陸地棉株型及生育期相關性狀QTL 定位
2021-04-14 06:57:04賈曉昀王士杰趙紅霞朱繼杰李妙王國印
棉花學報 2021年2期
賈曉昀,王士杰,趙紅霞,朱繼杰,李妙,王國印
(河北省農林科學院糧油作物研究所/ 河北省作物遺傳育種實驗室,石家莊050035)
株型是棉花的重要性狀,不同株型材料在產量、品質、抗逆性等方面存在顯著差異,理想株型是高產優質的保證[1-5]。 近年來,全程機械化成為棉花生產的重要發展方向,機采棉成為育種的主攻目標, 而機械采摘對株型的要求較為嚴格,因此挖掘株型相關分子遺傳機制成為基礎研究的重點[3,6]。 此外,株高越矮、株型越緊湊的材料早熟性越好,改良棉花早熟性,適當無膜晚播,有利于減少地膜使用量,保護耕地質量和環境[1,7-8]。
棉花株型性狀主要包括株高 (Plant height,PH)、第一果枝節位(Nodeofthefirstfruitingbranch,NFFB)及其高度(Height of NFFB,HNFFB)、果枝長度 (Fruiting branch length,FBL)、 果節長度(Branch node length,BNL) 及 果 枝 角 度(Fruit branch angle,FBA)等性狀;生育期主要包括出苗期(Budding time,BT)、現 蕾 期(Squaring time,ST)、開花期(Flowering time,FT)、花鈴期(Flowering to boll-opening period,FBP) 和全生育期(Whole growth period,WGP)[7,9-11]。 目前,關于棉花株型及生育期性狀的分子遺傳機制研究主要通過QTL(Quantitative trait loci,數量性狀位點)定位和GWAS (Genome-wide association analysis,全基因組關聯分析)挖掘相關染色體位點。 范術麗等[12]首先開展棉花株型及生育期性狀QTL定位研究,得到12 個相關QTL。Li 等[13]構建2 個F2群體定位到54 個生育期相關的QTL。 Li 等[14]針對10 個株型性狀開展QTL 定位, 在F2和F2:3群體定位到73 個位點。 Jia 等[11]以重組自交系為材料, 定位到247 個株型及生育期相關QTL 位點,并在D3 染色體上發現一段關鍵區域。Su 等[7]以355 份陸地棉為試驗材料,開展株型相關性狀GWAS,關聯到22 個SNP。 Ma 等[15]構建海陸種間回交群體,定位到10 個株高相關QTL,并驗證了1 個GhPIN3 基因。然而,棉花株型及生育期性狀均為復雜的數量性狀,目前得到的基因信息較少,難以為品種選育提供有效參考。
本研究立足生產需求, 以冀豐914 和冀豐817 為親本構建F2群體,分析棉花株型及生育期性狀的相關性。 采用GBS(Genotyping-by-sequencing, 基于測序的基因分型) 技術開發群體SNP(Single nucleotide polymorphism,單核苷酸多態性) 標記, 構建了1 張包含11 488 個SNP 標記、總圖距為4 202.12 cM(centiMorgan,厘摩)的高密度遺傳圖譜(數據未發表),定位與株型、生育期相關的QTL,為深入分析棉花株型及生育期性狀的分子遺傳機制奠定研究基礎,為機采棉分子標記輔助育種提供理論指導。
1 材料與方法
1.1 試驗材料
以自育國審品種冀豐914(國審棉2015003)為母本, 以優質自交系冀豐817 (原系名: 優系817)為父本。 冀豐914 株型高大、豐產性好,冀豐817 株高較低、纖維品質優良。 于2018 年在河北石家莊組配雜交組合,收獲F1種子,同年冬天在海南加代自交,收獲F2種子。于2019 年在河北石家莊播種F2群體413 個單株, 行長7 m, 行距0.76 m,株距0.2 m,共15 行,管理同當地大田。
現蕾后調查NFFB,9 月初調查PH、HNFFB、FNLFFB (First node length of the fourth fruiting branch, 第四果枝第一果節長度)、FNLSFB(First node length of the seventh fruiting branch, 第七果枝第一果節長度);全生育期內及時調查開花期和吐絮期,統計FT、FBP 和WGP。
1.2 分析方法
通過Excel 2016 對數據的基本統計量進行分析,通過SPSS21.0 分析性狀之間的簡單相關性。
通過QTL IciMapping 4.0 分析加性及上位性QTL[16]。 加性QTL 的分析參數為:Step=1 cM,PIN=0.001。 上位性QTL 的分析參數為:Step=5 cM,PIN=0.000 1。LOD 值通過1 000 次迭代計算確定。 本研究中,母本基因型為BB,父本基因型為AA;加性效應為負,表示對應QTL 的增效基因來源于母本冀豐914;加性效應為正,表示對應QTL 的增效基因來源于父本冀豐817。
1.3 基因注釋及表達量分析
根據Wang 等[17]公布的參考基因組信息對重要QTL 位點進行基因注釋,并通過KOBAS 3.0軟件[18]進行KEGG 通路和GO 富集分析,根據Hu 等[19]公布的轉錄組信息進行注釋基因表達量分析。
2 結果與分析
2.1 群體表型的基本統計
表1 為群體性狀基本統計量。 除花鈴期外,冀豐914 的其他性狀值顯著大于冀豐817,F1的性狀表型值居于親本之間,F2群體性狀呈現雙向超親分布;峰度和偏度的絕對值小于1,近似正態分布,適合進行QTL 分析。 由標準差和變異系數可 以 看 出, 試 驗 群 體 的PH、NFFB、HNFFB、FNLFFB 和FNLSFB 等5 個株型性狀的變異幅度顯著大于FT、FBP 和WGP 等3 個生育期性狀,說明試驗群體的株型性狀變異豐富,含有豐富的株型相關QTL/基因。

表1 親本及群體性狀的基本統計量Table 1 Statistics of the parents and population traits
2.2 性狀之間的簡單相關性分析
表2 為性狀之間簡單相關性分析。 可以看出,除FT 與FBP 負相關外,其他性狀間均為正相關。PH 與NFFB、HNFFB、FNLFFB、FNLSFB 的相關性達到極顯著水平, 與WGP 的相關性達到顯著水平。 NFFB 與HNFFB、FNLFFB、FT、WGP的相關性達到極顯著水平,與FBP 的相關性達到顯著水平。HNFFB 與FNLFFB、FT 的相關性達到極顯著水平, 與WGP 的相關性達到顯著水平。FNLFFB 與FNLSFB、FT、WGP 的相關性達到極顯著水平;FNLSFB 與WGP 的相關性達到極顯著水平,與FBP 的相關性達到顯著水平。WGP 與FT、FBP 的相關性達到極顯著水平。 可知,PH 與其他4 個株型性狀均為極顯著正相關,NFFB 與生育期性狀均為顯著/極顯著正相關。
2.3 株型性狀的主成分分析
表3 為株型性狀主成分分析結果,前兩個主成分因子累計貢獻率為61.53%。因子1 的特征值為2.03,貢獻率為40.53%,特征向量值較大的是HNFFB(0.74)和PH(0.73),主要是垂直高度相關性狀。因子2 的特征值為1.05,貢獻率為21.00%,特征向量值較大的是FNLSFB(0.77),為水平長度相關性狀。 因此,PH、HNFFB 和FNLSFB 是鑒定棉花株型垂直和水平方向特征的典型性狀。
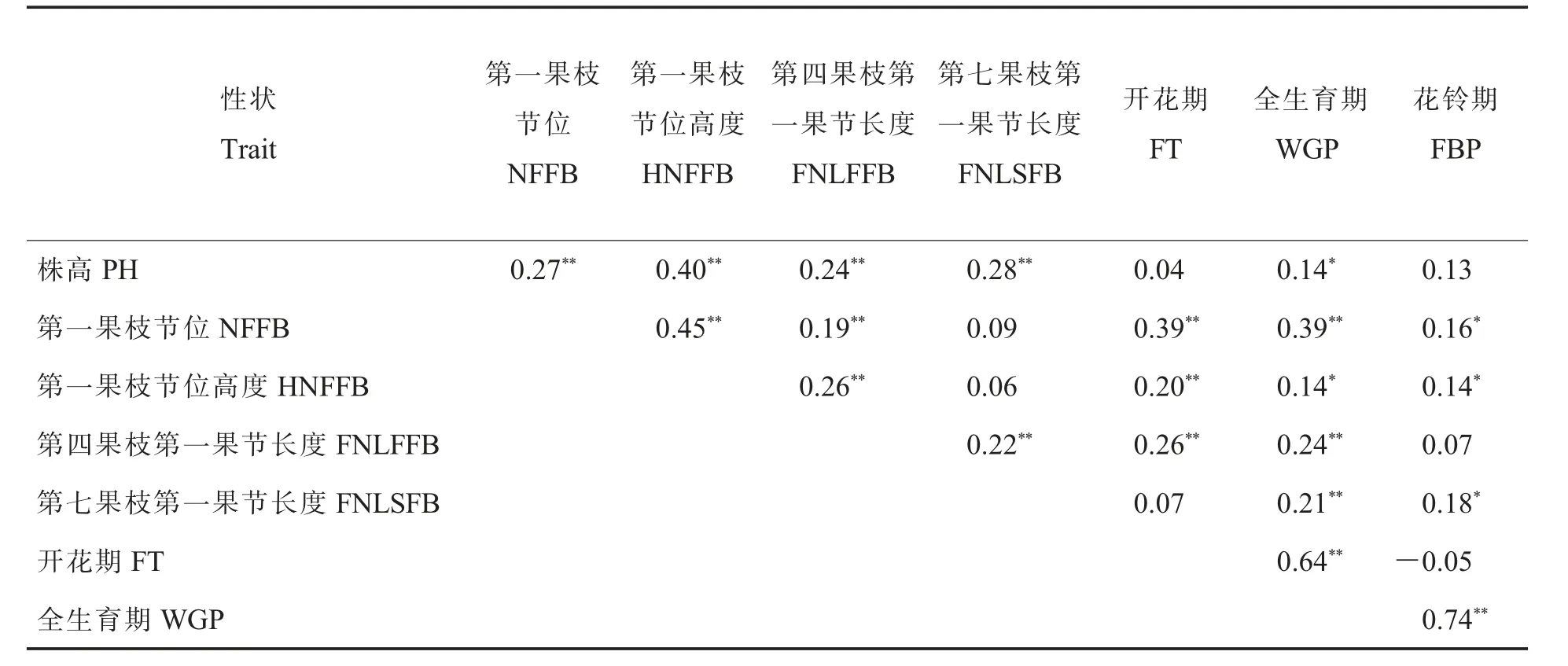
表2 各性狀之間的相關性分析Table 2 Correlation analysis among the seven traits

表3 五個株型性狀主成分分析Table 3 Principal component analysis of the five plant architecture traits
2.4 QTL 定位結果及分析
本研究共得到36 個加性QTL(Additive QTL,aQTL)位點,包括31 個株型相關aQTL 和5 個生育期相關aQTL,其中26 個位點的增效基因來源于母本冀豐914,10 個位點的增效基因來源于父本冀豐817 (表4 和圖1)。 定位到14 個PH 相關aQTL, 分布于11 條 染色體。 其 中,qPH-D12-1 的貢獻率最大, 為9.64%,LOD 值達到11.97,增效基因來源于母本冀豐914;qPHD2-1 和qPH-D5-1 的貢獻率分別為8.53%和8.62%,LOD 值分別為10.85 和10.86, 增效基因均來源于母本冀豐914。 定位到6 個NFFB 相關aQTL, 分布于5 條染色體。 其中,qNFFB-A11-1的貢獻率最大,為8.63%,增效基因來源于母本冀豐914。 定位到3 個HNFFB 相關aQTL,分布于A12、D3 和D8 染色體, 貢獻率分別為6.18%、5.61%和6.75%。 定位到5 個FNLFFB 相關aQTL, 分布于A2、A6、D8 和D11 染色體,A6 染色體分布有2 個位點, 即qFNLFFB-A6-1 和qFNLFFB-A6-2, 這兩個位點的貢獻率分別為10.36%和10.38%, 其中qFNLFFB-A6-1 的增效基因來源于父本冀豐817,qFNLFFB-A6-2 的增效基因來源于母本冀豐914。 定位到3 個FNLSFB 相關aQTL,分布于A9、D8 和D10 染色體,其中qFNLSFB-D8-1 的位置與qFNLFFBD8-1 相近,位于135~139 cM 區段,增效基因均來源于父本冀豐817。 分別定位到2、1、2 個FT、WGP、FBP 相關aQTL,分布于5 條染色體,貢獻率為6.29%~8.11%。
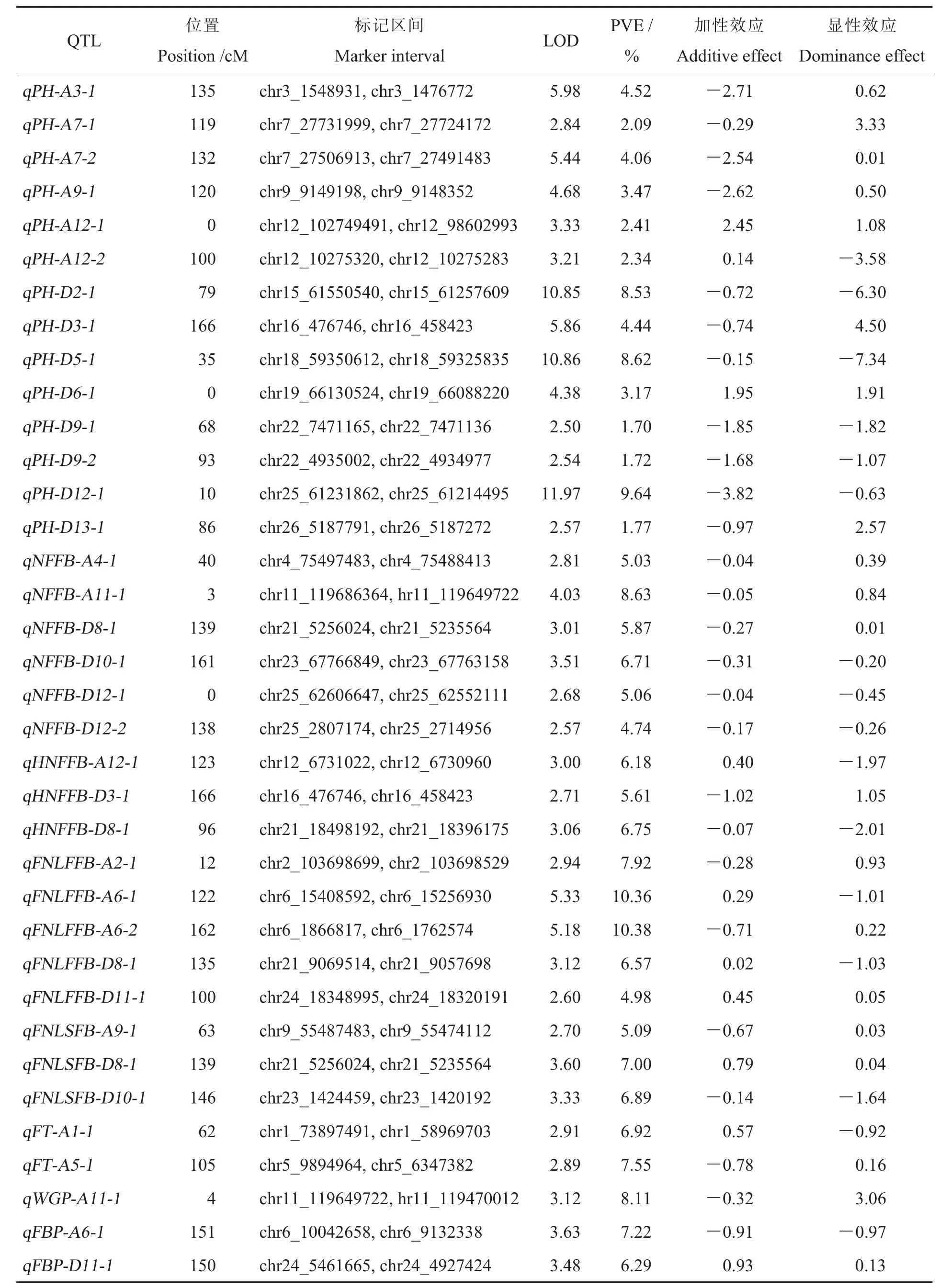
表4 株型及生育期相關性狀加性QTL 信息Table 4 Additive effect QTL information of the plant architecture and growth period related traits

圖1 定位的加性QTL 在染色體上的分布Fig. 1 Additive QTL distribution on chromosomes
發現3 個染色體片段處同時分布有2 個aQTL 位點, 分別為A11 染色體1.5~4.5 cM 處qNFFB-A11-1 和qWGP-A11-1 重疊、D3 染色體157.5~166 cM 處qHNFFB-D3-1 和qPH-D3-1 重疊、D8 染色體138.5~139.5 cM 處qNFFB-D8-1和qFNLSFB-D8-1 重 疊。 其 中,D8 染 色 體 的134.5~135.5 cM 有1 個qFNLFFB-D8-1。 因此,D8 染色體135~139 cM 區段內可能存在與株型相關的關鍵基因。
此外, 定位到263 個上位性QTL(Epistatic QTL,eQTL)位點,涉及全部的26 條染色體(附表1)。 其中31 個與PH 相關、28 個與NFFB 相關、19 個與HNFFB 相關、62 個與FNLFFB 相關、21個 與FNLSFB 相 關、44 個 與FT 相 關、21 個 與WGP 相關和37 個與FBP 相關,單個eQTL 的貢獻率為1.17%~6.19%。 其中,96 個eQTL 分布于41 個染色體位點, 每個位點至少有2 個eQTL;21 個eQTL 與19 個aQTL 位置重疊,但是僅有4個位點影響相同性狀,說明在互作條件下同一個遺傳位點表現出復雜遺傳效應。
2.5 基因注釋及候選基因篩選
基于Wang 等[17]公布的陸地棉參考基因組注釋信息,針對貢獻率大于8.00%的加性QTL 所在區間及D8 染色體的135~139 cM 區段進行基因注釋,共注釋到115 個基因。 通過KEGG 通路比對發現,注釋基因比對到37 個KEGG 通路(附圖1);GO 富集結果發現,注釋基因主要富集到纖維素酶活性等生物活性、纖維素及植物初生細胞壁生物合成等生物過程、細胞內膜結合細胞器細胞組分等40 個GO 條目(附圖2)。 根據Hu 等[19]公布的轉錄組信息,對注釋得到的115 個基因進行表達量分析,篩選到在根、莖、葉、花或萼片5個組織表達量較高的基因共21 個, 其中GH_D02G1779 和GH_D08G0595 在葉片中表達量最高,GH_D08G0525 在莖中表達量最高,GH_D08G0638 在根和花中表達量最高(圖2)。

圖2 根、莖、葉、花、萼片中表達量較高的注釋基因Fig. 2 Highly expressed genes in root, stem, leaf, flower and sepal
3 討論
株型是植物特征特性的外在表現,是進行植物鑒定、分類的重要依據。 株型的形成是遺傳因素與環境條件共同作用的結果,受到內源激素與外源刺激的協同調控[20]。 株型也是農作物產量和品質形成的基礎, 三者之間存在顯著的相互影響,理想株型是實現高產、優質的重要保障[3,21-23]。近年來,機械化的發展特別是機械采摘對棉花株型提出更嚴格的要求[7,24],合理的株型有利于提高機械采摘質量,減少原棉的雜質污染。 因此,開展株型育種、培育適于機采的棉花新品種對發展植棉產業、提高經濟效益有重要影響。 加強棉花株型性狀的分子基礎研究, 挖掘株型相關遺傳位點,有利于提高分子標記輔助育種效率、加快機采棉育種速度。
本研究以國審品種冀豐914 為母本、自交系冀豐817 為父本構建F2分離群體,分析5 個株型性狀和3 個生育期性狀表型和遺傳基礎。 經簡單相關性分析和主成分分析發現,本文研究的大部分性狀間存在正相關關系,株高與其他株型性狀均為極顯著正相關,第一果枝節位與生育期性狀均為顯著或極顯著正相關,可作為鑒定棉花早熟性的株型性狀,株高、第一果枝節位高度和第七果枝第一果節長度是鑒定株型垂直和水平特征的典型性狀,與他人研究結果相似[11,25-27]。 本研究調查的第四和第七果枝第一果節長度在其他研究中報道較少。據紀從亮等[28]研究,第一果節長度與產量呈正相關,并且對棉株內圍構造有顯著影響。 因此,本研究結合試驗材料特征,分別測量棉株下部和中部一個果枝的第一果節長度,不僅有助于了解棉花株型特征,而且為下一步研究株型與產量之間的關系奠定基礎。結果表明,第七果枝第一果節長度可用于鑒定棉花株型的水平特征。
本實驗室前期基于Wang 等[17]公布的參考基因組,構建了1 張包含11 488 個SNP 標記、總圖距為4 202.12 cM 的高密度遺傳圖譜 (數據未發表)。 基于此圖譜,本研究定位到36 個株型及生育期相關的aQTL 位點, 冀豐914 為其中26 個位點提供增效基因, 冀豐817 為其中10 個位點提供增效基因,說明冀豐914 對株型及生育期的可遺傳控制更強,可作為機采棉品種選育的基礎材料。 比對前人的研究結果[7,11,14],本研究定位的17 個aQTL 與已報道的aQTL 位置重疊或相近,例如本研究得到的qPH-D3-1 與Su 等[7]、Jia 等[11]在D3 染色體上得到的區段相近, 另外19 個QTL 未見報道。 由于株型相關的QTL 定位、GWAS 分析等相關研究較少,因此本研究得到的新位點較多。 此外,冀豐914 與冀豐817 首次作為試驗材料開展株型及生育期的基礎研究,與其他的研究材料存在一定遺傳差異。 本研究發現了3 個aQTL 重疊區段,其中在D8 染色體上的區段內分布有3 個aQTL 位點,可能存在與株型相關的關鍵候選基因。 At 基因組和Dt 基因組分別分布有17 個和19 個aQTL, 說明2 個亞組中控制株型的遺傳位點數量基本相同。同時,檢測到263個eQTL,顯著多于aQTL 位點數,說明全基因組范圍內的互作效應對株型及生育期有重要影響;共在19 個aQTL 位點檢測到了eQTL, 而僅有4個位點影響相同的性狀,說明在互作條件下,同1個aQTL 位點表現出更多的遺傳效應;At 基因組的eQTL 顯著多于Dt 基因組,可能是由于At 基因組的長度更長,因而檢測到的上位性互作位點更多[29]。 QTL 之間的重疊及互作關系,可能是造成表型性狀之間復雜相互關系的重要遺傳因素。本研究發現的3 個QTL 重疊區段內, 包含了PH、NFFB、HNFFB、FNLSFB、FNLFFB 及WGP 等多個性狀相關的QTL 位點, 由此也可以解釋株型性狀及生育期性狀之間的顯著相關性。 如PH與HNFFB 極顯著正相關,QTL 定位發現qPH-D3-1 與qHNFFB-D3-1 重疊,且增效基因均來自于母本冀豐914。 同時,根據參考基因組信息[17,19],本研究注釋到21 個在陸地棉TM-1 和海島棉海7124 的根、莖、葉、花、萼片等組織中表達量較高的基因,其中GH_D02G1779、GH_D08G0595、GH_D08G0525、GH_D08G0638 在不同組織中的表達量最高。 GH_D02G1779 位于株高相關QTL qPH-D2-1 區間;GH_D08G0595、GH_D08G0525、GH_D08G0638 均位于D8 染色體的135~139 cM區段內,該區段是第一果枝節位、第四果枝第一果節長度和第七果枝第一果節長度相關QTL 的重疊區域。 因此,這4 個基因均位于本研究定位到的重要區段內,可能參與棉花株型的調控。海7124 株高 較 高,TM-1 株 高 一 般,GH_D08G0525 在 海7124 莖中的表達量顯著高于TM-1, 推測該基因可能參與棉花株高的調控。
4 結論
株高、第一果枝節位高度和第七果枝第一果節長度是鑒定棉花株型垂直和水平特征的典型性狀,第一果枝節位可作為鑒定棉花早熟性的株型性狀。 定位到36 個株型及生育期相關QTL 位點, 單個QTL 對表型變異的貢獻率為1.70%~10.38%,發現3 個QTL 重疊的染色體區段,并在qPH-D2-1 和Chr. D8 的135~139 cM 區段 內發現4 個表達量較高的基因,為進一步分析棉花株型的分子遺傳機制及候選基因篩選奠定了基礎。
附圖、附表:
附圖、附表詳細內容參見http://journal.cricaas.com.cn/
附圖1 KEGG 通路分析結果
Fig.S1 Results of the KEGG pathway mapping
附圖2 GO 富集結果
Fig.S2 Results of the GO enrichment analysis
附表1 上位性QTL 檢測結果
Table S1 Results of the epistatic QTL
