海南島鳥(niǎo)巢蕨自然種群遺傳多樣性SRAP 分析
2021-04-15 01:06:14侯祥文徐詩(shī)濤王德立梁靖雯司更花
熱帶生物學(xué)報(bào) 2021年1期
關(guān)鍵詞:物種
侯祥文,徐詩(shī)濤,王德立,梁靖雯,司更花
(1.海南大學(xué) 林學(xué)院,海口 570228; 2.海南大學(xué) 園藝學(xué)院,海口 570228;3.中國(guó)醫(yī)學(xué)科學(xué)院 藥用植物研究所 海南分所,海口 570311)
鳥(niǎo)巢蕨(Asplenium nidus)是鐵角蕨科Aspleniaceae 巢蕨屬的一個(gè)種,又名山蘇花,是熱帶雨林大型附生植物[1]。原生于亞洲東南部、澳大利亞?wèn)|部、印度尼西亞、印度和非洲東部等地[2?3]。《中國(guó)植物志》記載我國(guó)有鳥(niǎo)巢蕨分布于臺(tái)灣、福建、香港、廣東、廣西、海南、湖南、四川、貴州、云南和西藏等省份和地區(qū)。鳥(niǎo)巢蕨植物不僅可以豐富森林生物多樣性,還可調(diào)節(jié)森林生態(tài)系統(tǒng)的水分與養(yǎng)分循環(huán),且具有藥用、食用與園林綠化價(jià)值。近年來(lái),由于人為活動(dòng)干擾,野生鳥(niǎo)巢蕨被大量采集,生境縮小且破碎化嚴(yán)重,導(dǎo)致其遺傳多樣性降低。加上種間雜交嚴(yán)重,導(dǎo)致遺傳背景錯(cuò)綜復(fù)雜,已影響到其品種改良、新品種培育、保護(hù)和開(kāi)發(fā)利用。因此,加強(qiáng)對(duì)鳥(niǎo)巢蕨遺傳多樣性與進(jìn)化研究對(duì)鳥(niǎo)巢蕨種質(zhì)資源保護(hù)和利用以及育種和遺傳改良具有重要意義。SRAP(sequence-related amplified polymorphism)是相關(guān)序列擴(kuò)增多態(tài)性的簡(jiǎn)稱(chēng),利用雙引物設(shè)計(jì)對(duì)基因的ORFS(Open Reading Frames,開(kāi)放閱讀框)進(jìn)行區(qū)域擴(kuò)增是該技術(shù)特點(diǎn),因不同物種內(nèi)含子、啟動(dòng)子與間隔區(qū)的長(zhǎng)度不同而產(chǎn)生多態(tài)性[4?6]。SRAP 技術(shù)具有操作方便、多態(tài)性豐富、重復(fù)性好、易于測(cè)序、速度快和成本低等特性,被廣泛運(yùn)用于動(dòng)物、植物、微生物的品種鑒定、遺傳圖譜構(gòu)建、遺傳多樣性檢測(cè)、基因定位和比較基因組學(xué)等研究[7?8]。SRAP 標(biāo)記通過(guò)直接分析遺傳物質(zhì)多態(tài)性作為不同種群劃分依據(jù)更為可行和客觀,能夠較好地開(kāi)展遺傳多樣性分析,可避免因季節(jié)性生理生化差異和不同發(fā)育階段而影響指標(biāo)的準(zhǔn)確性[9?10]。因此,可通過(guò)分子標(biāo)記技術(shù)分析鳥(niǎo)巢蕨親緣關(guān)系,得到更加科學(xué)和穩(wěn)定的分析結(jié)果。海南島是鳥(niǎo)巢蕨分布中心之一[11?12],野生種群集中分布于霸王嶺、俄賢嶺、黎母山和五指山等熱帶雨林區(qū)域。不同種群間的鳥(niǎo)巢蕨在葉形、株高、孢子量、巢基大小方面存在一定差異[13?14]。為揭示鳥(niǎo)巢蕨自然種群狀況,促進(jìn)野生鳥(niǎo)巢蕨資源有效保護(hù)和利用,本研究采用SRAP 標(biāo)記技術(shù),對(duì)海南島霸王嶺、俄賢嶺、黎母山和五指山6 個(gè)鳥(niǎo)巢蕨自然種群進(jìn)行遺傳多樣性分析,旨在探明海南島鳥(niǎo)巢蕨種群之間遺傳關(guān)系、親緣關(guān)系,進(jìn)而揭示海南島不同地理環(huán)境對(duì)鳥(niǎo)巢蕨遺傳多樣性及種群分化的影響,為鳥(niǎo)巢蕨保育和遺傳資源利用提供理論依據(jù)。
1 材料與方法
1.1 材料于2011 年5 月?8 月及2019 年3 月?4 月,在海南島霸王嶺、俄賢嶺、黎母山及五指山6 個(gè)自然種群進(jìn)行采樣,每個(gè)種群范圍均>2 km2,6 個(gè)自然種群相互間地理距離>10 km。每個(gè)采集種群內(nèi)采樣個(gè)體間間隔距離>1 m。共采集96 份植株葉片樣本,硅膠干燥后,帶回實(shí)驗(yàn)室,提取DNA。各種群地理位置、生境及采樣個(gè)體數(shù)詳見(jiàn)表1。
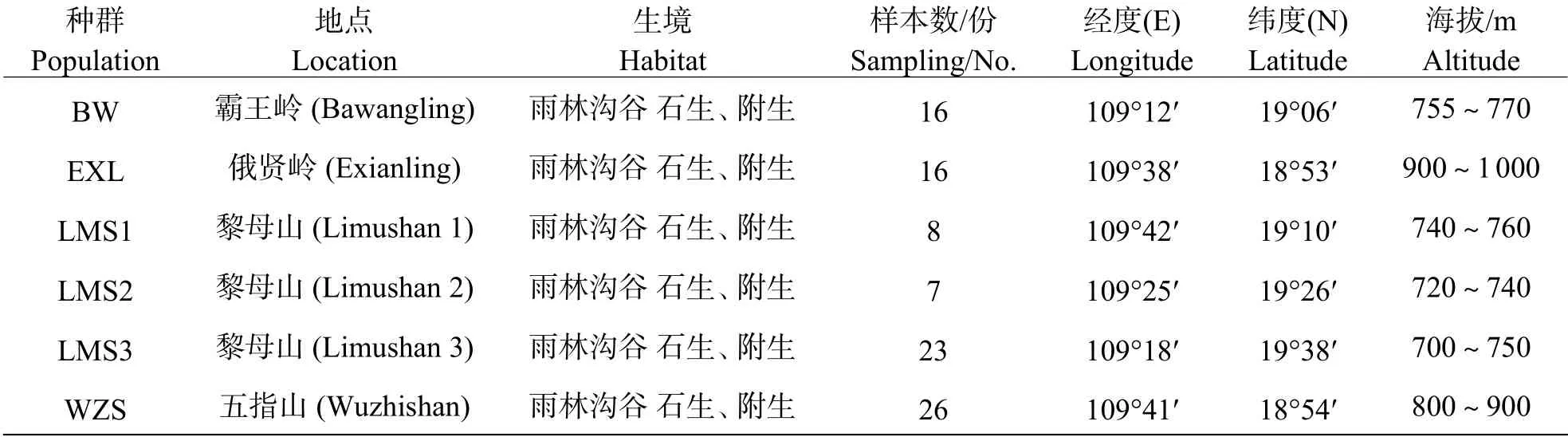
表1 海南島鳥(niǎo)巢蕨6 個(gè)研究種群的地理位置與生境Tab.1 Location, habitat, and sample size of the 6 populations of Asplenium nidus in Hainan Island
1.2 DNA 提取采用北京天根生物有限公司生產(chǎn)的新型植物基因組DNA 提取試劑盒(DP305)提取鳥(niǎo)巢蕨葉片DNA,用w=1.0%瓊脂糖凝膠電泳檢測(cè),圖1 為鳥(niǎo)巢蕨種群基因組DNA 電泳圖。提取樣品基因組DNA 用核酸微量測(cè)定儀稀釋至一致的質(zhì)量濃度(50 mg·L?1),置于?20 ℃下保存?zhèn)溆谩?/p>
1.3 SRAP 擴(kuò)增與檢測(cè)分析根據(jù)LI 等[15]的方法,設(shè)計(jì)正向引物17 個(gè),反向引物20 個(gè),由北京三博生物技術(shù)有限公司合成。PCR 擴(kuò)增反應(yīng)使用TC-512PCR 儀(Techne,Ltd.)。PCR 擴(kuò)增體系:1U 的TaqDNA 酶,1.5 mmol·L?1的Mg2+,0.20 mmol·L?1的dNTP(Fermentas Biotec.)和25 ng 的DNA,0.5 μmol·L?1的引物。PCR 擴(kuò)增程序:94 ℃預(yù)變性5 min, 94 ℃變性30 sec,52 ℃退火30 sec,72 ℃延伸1 min,共35 個(gè)循環(huán);最后72 ℃終延伸10 min,4 ℃保存[16]。擴(kuò)增產(chǎn)物在w=6%變性聚丙烯酰胺凝膠上(65 w 恒定功率)電泳2~2.5 h。電泳完畢后,進(jìn)行銀染,通過(guò)成像儀拍照并保存。
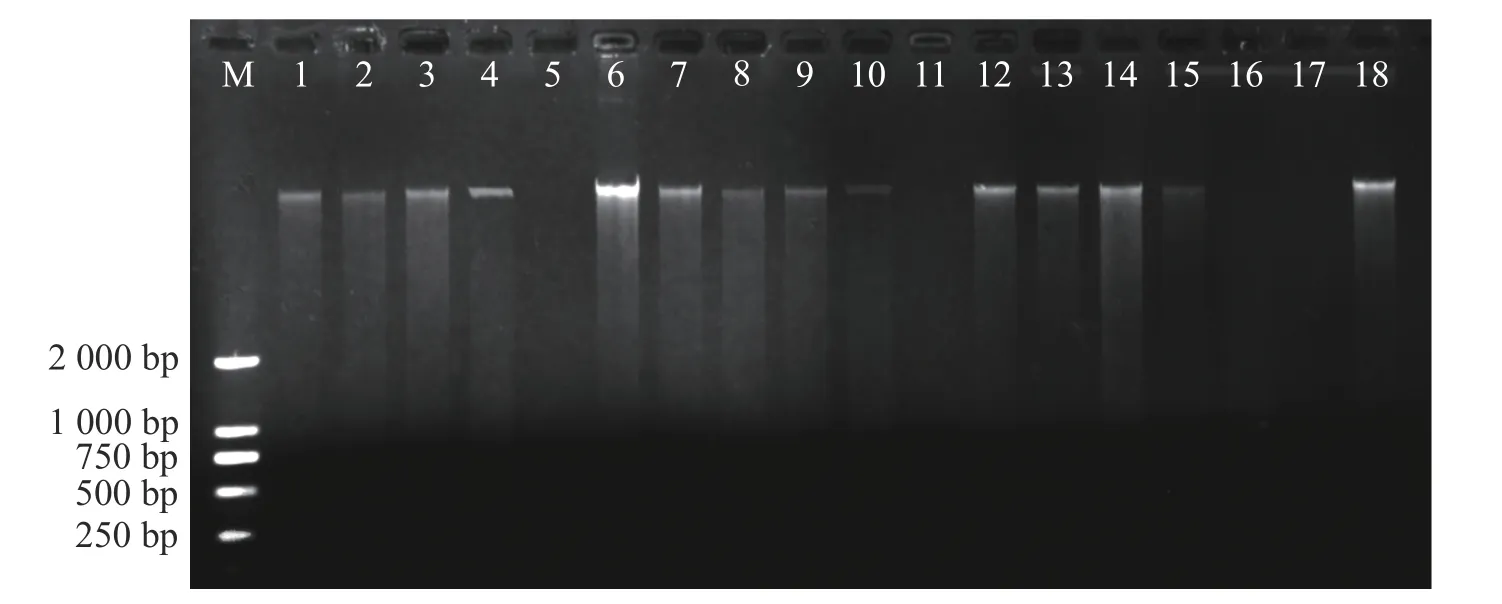
圖1 海南島6 個(gè)鳥(niǎo)巢蕨種群基因組 DNA 電泳圖Fig.1 The electrophoretogram of genomic DNA of 6 populations of Asplenium nidus in Hainan Island
1.4 數(shù)據(jù)統(tǒng)計(jì)分析將SRAP 記錄的電泳圖進(jìn)行人工讀帶,由圖1 可得,以同一位點(diǎn)有帶記作“1”,無(wú)帶記作“0”,僅記錄清晰、穩(wěn)定且長(zhǎng)度在300~2 500 bp 的擴(kuò)增條帶,形成0-1 矩陣二元數(shù)據(jù)輸入電腦。
采用POPGENE32 軟件衡量SRAP 水平上遺傳變異程度,對(duì)全部種群和各個(gè)種群分別進(jìn)行遺傳參數(shù)分析,分別對(duì)有效等位基因數(shù)(Ne, effective number of allele)、等位基因觀察值(Na,number of allele)、多態(tài)位點(diǎn)百分率(PPB,percentage of polymorphic bands)、Nei’s 基因多樣性(He,Nei’s genetic diversity)、Shannon 信息指數(shù)(Ho,Shannon’s information index)[16?17]、種群總基因多樣度(Ht,total gene diversity)、種群內(nèi)基因多樣度(Hs,gene diversity within populations)、種群間的基因流(N,geneflow),種群間遺傳分化系數(shù)(Gst,coefficient of population differentiation)、Nei’s 無(wú)偏差遺傳距離(D,Nei’s genetic distance)和遺傳一致度(I,genetic identity)等參數(shù)進(jìn)行計(jì)算[18]。
應(yīng)用GenAlEx v.6(Genetic Analysis in Excel)軟件[19]對(duì)種群內(nèi)遺傳變異和種群間遺傳變異進(jìn)行分子變異分析(AMOVA),計(jì)算遺傳變異在種群內(nèi)和種群間分布。Shannon 信息指數(shù)HO由如下公式計(jì)算:
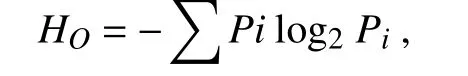
式中,HO為Shannon 信息指數(shù),Pi為第i條帶的表型頻率。
遺傳分化系數(shù)(coefficient of genetic differentiation,Gst)用來(lái)估算種群間的遺傳分化程度,即種群間遺傳多樣性占總遺傳多樣性的比例。
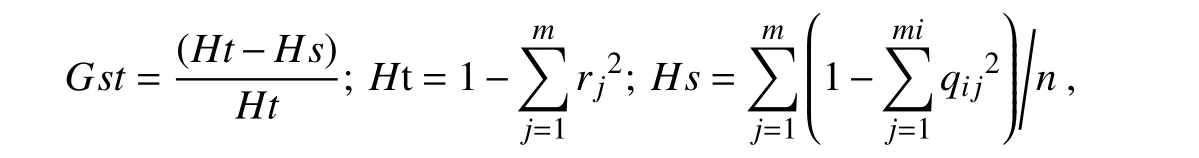
式中,Gst為遺傳分化系數(shù),Ht為種群的遺傳多樣度,Hs為各種群內(nèi)的基因多樣度,m為該位點(diǎn)上的等位基因,n為居群總數(shù),rj為該位點(diǎn)上第j個(gè)等位基因在總種群中的平均頻率,qij為第i個(gè)種群在該位點(diǎn)上第j個(gè)等位基因頻率,mi為第i個(gè)種群在該位點(diǎn)上的等位基因數(shù)。
基因流(gene flow)用來(lái)估算種群間基因交流程度,為每個(gè)世代在種群間遷移的個(gè)體數(shù),是通過(guò)遺傳分化系數(shù)來(lái)估算的:
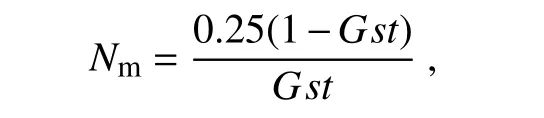
式中,Nm為基因流,N為有效種群大小;m為種群每代遷移率;Gst為遺傳分化系數(shù)。
根據(jù) Nei’s 遺傳距離,使用NTSYS-pc 2.1 軟件對(duì)種群進(jìn)行UPGMA 聚類(lèi)分析,樣本數(shù)為85。
2 結(jié)果與分析
2.1 海南島鳥(niǎo)巢蕨自然種群的遺傳多樣性利用17 個(gè)正向引物與20 個(gè)反向引物共340 對(duì)引物組合進(jìn)行擴(kuò)增,篩選出10 對(duì)條帶清晰、穩(wěn)定性高、特異性好且多態(tài)性豐富的PCR 擴(kuò)增產(chǎn)物用于鳥(niǎo)巢蕨的遺傳多樣性分析[15]。其中,正向和反向引物序列見(jiàn)表2,10 對(duì)引物組合為:ME3-EM3、ME4-EM10、ME5-EM8、ME6-EM16、ME5-EM7、ME10-EM10、ME14-EM6、ME16-EM14、ME16-EM10、ME3-EM4。
引物用于對(duì)海南島鳥(niǎo)巢蕨6 個(gè)野生種群共96 個(gè)個(gè)體進(jìn)行PCR 擴(kuò)增,由擴(kuò)增結(jié)果(圖2)可看出,一共擴(kuò)增出184 個(gè)清晰穩(wěn)定的條帶,篩選6 條清晰條帶進(jìn)行分析,其中174 條具有多態(tài)性,多態(tài)位點(diǎn)百分率為94.57%。從圖1 可看出,每個(gè)引物擴(kuò)增出的條帶為5~9 條,平均條帶數(shù)為6.9。條帶分子量在250~2 000 bp之間,而表3 里的各種群間多態(tài)位點(diǎn)百分率存在較大的差異。基于SRAP 分子標(biāo)記的鳥(niǎo)巢蕨物種水平多態(tài)位點(diǎn)百分比(PPB)為94.57%,SRAP 檢測(cè)到的種群多態(tài)位點(diǎn)百分比例(PPB)在76.09%(LMS2 種群)~94.57%(LMS3 種群)之間,平均為85.15%,鳥(niǎo)巢蕨種群間遺傳變異程度較大(76.09%~94.57%),說(shuō)明各種群均具有較高的遺傳多樣性水平。
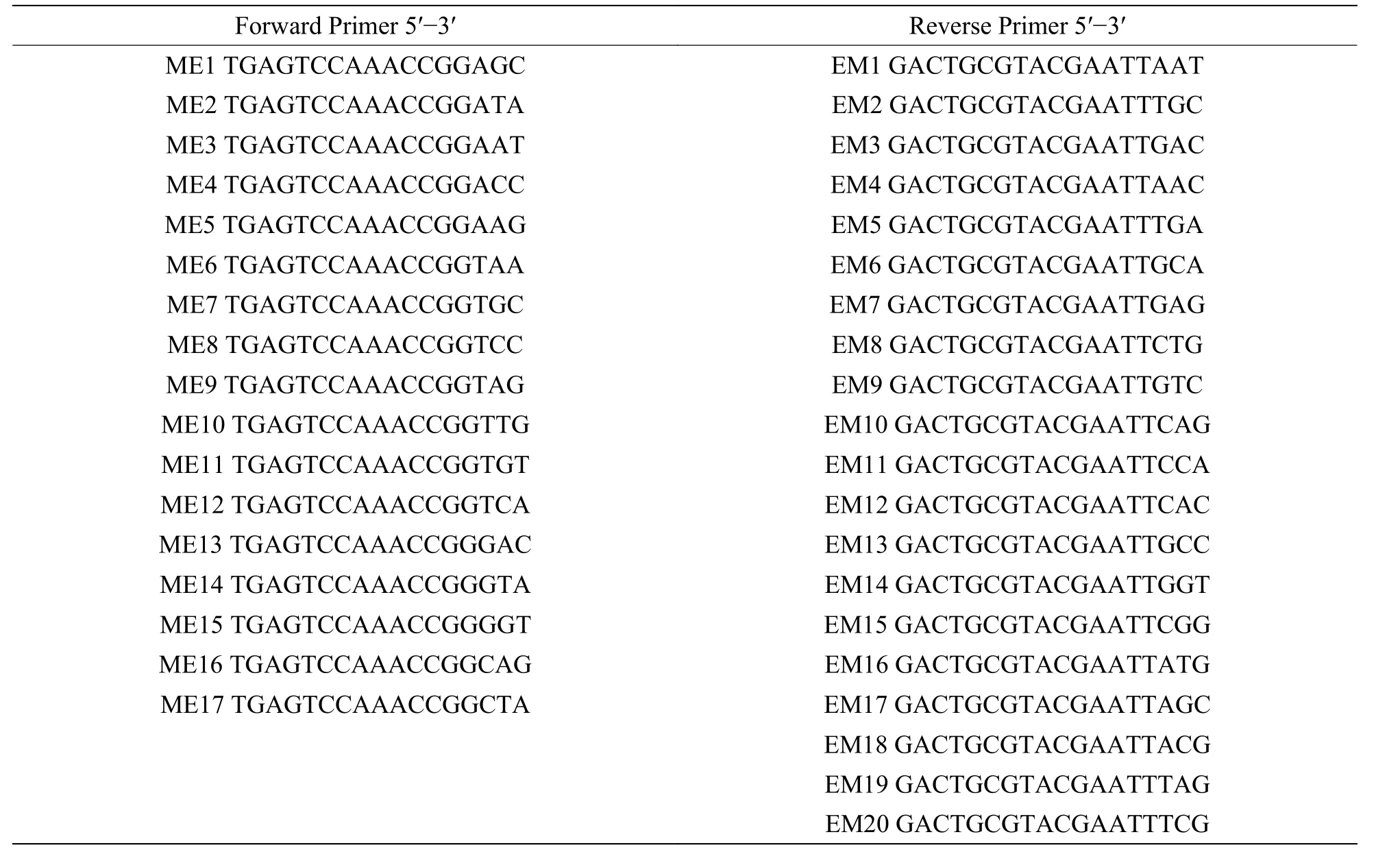
表2 SRAP 分析中使用的正向和反向引物序列Tab.2 Forward and Reverse Primer sequences used in SRAP analysis
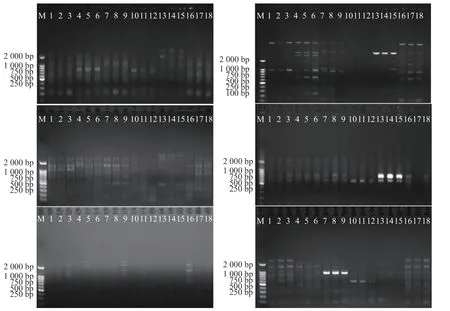
圖2 6 個(gè)鳥(niǎo)巢蕨鳥(niǎo)巢蕨種群的PCR 擴(kuò)增Fig.2 PCR amplification of six populations of Asplenium nidus
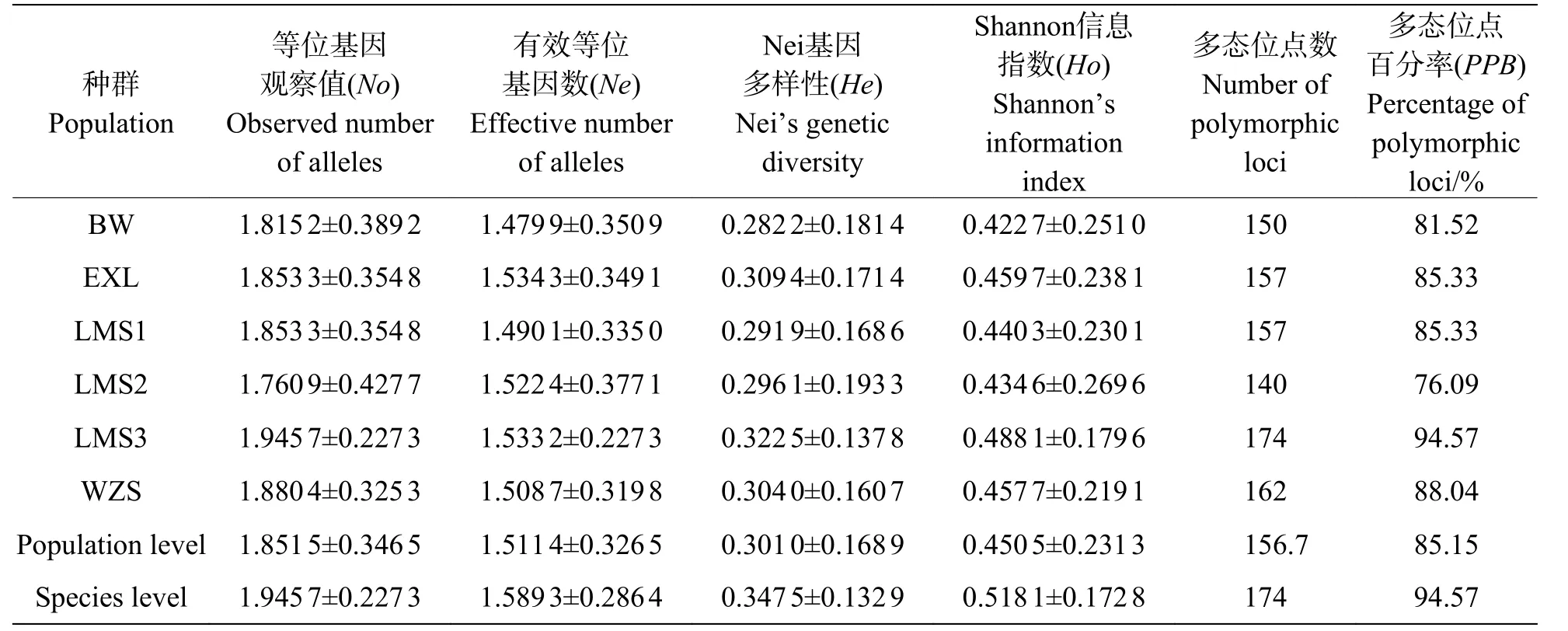
表3 鳥(niǎo)巢蕨6 個(gè)自然種群SRAP 標(biāo)記遺傳多樣性(平均值±標(biāo)準(zhǔn)差)Tab.3 Genetic diversity (Mean±SE) of 6 natural populations of Asplenium nidus based on SRAP markers
2.2 鳥(niǎo)巢蕨的遺傳分化POPGENE 軟件分析結(jié)果表明,種群間和種群內(nèi)均存在一定遺傳變異,但主要發(fā)生在種群內(nèi)部。種群總基因多樣度(Ht)為0.346 2±0.018 3,其中,種群內(nèi)基因多樣度(Hs)為0.301 0±0.014 2,即86.94%的遺傳分化發(fā)生在種群內(nèi)部;種群間遺傳分化系數(shù)(Gst)為0.130 6,即種群間遺傳分化為13.06%,說(shuō)明鳥(niǎo)巢蕨種群間遺傳分化水平較高,且遺傳變異主要發(fā)生在種群內(nèi)部。種群間基因流Nm=1.664 2,表明鳥(niǎo)巢蕨之間基因流動(dòng)暢通。
AMOVA 分析表明,遺傳變異以居群內(nèi)變異為主,此分析結(jié)果與POPGENE 的分析結(jié)果基本一致,由表4 可看出,顯著的遺傳分化(P= 0.010)于種群內(nèi)部和種群間均存在,遺傳變異在種群間占總遺傳變異的10.62%,在種群內(nèi)占89.53%。
2.3 鳥(niǎo)巢蕨種群間的遺傳距離與相似系數(shù)從表5可知,鳥(niǎo)巢蕨6 個(gè)種群的平均遺傳相似度為0.922 4,平均遺傳距離為0.080 9。其中,BW 和EXL 的遺傳距離最近,為0.050 8,LMS2 和LMS3 遺傳距離最遠(yuǎn),為 0.116 6。
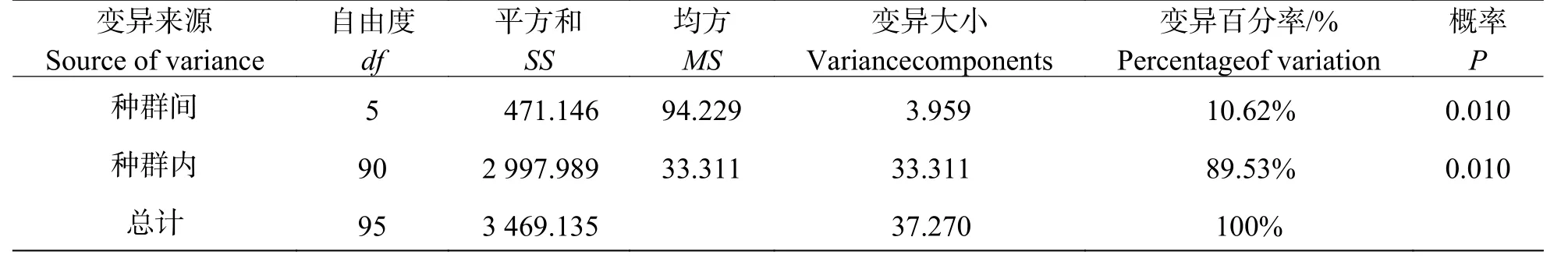
表4 鳥(niǎo)巢蕨自然種群遺傳變異的AMOVA 分析Tab.4 Analysis of molecular variance (AMOVA) of natural populations of Asplenium nidus
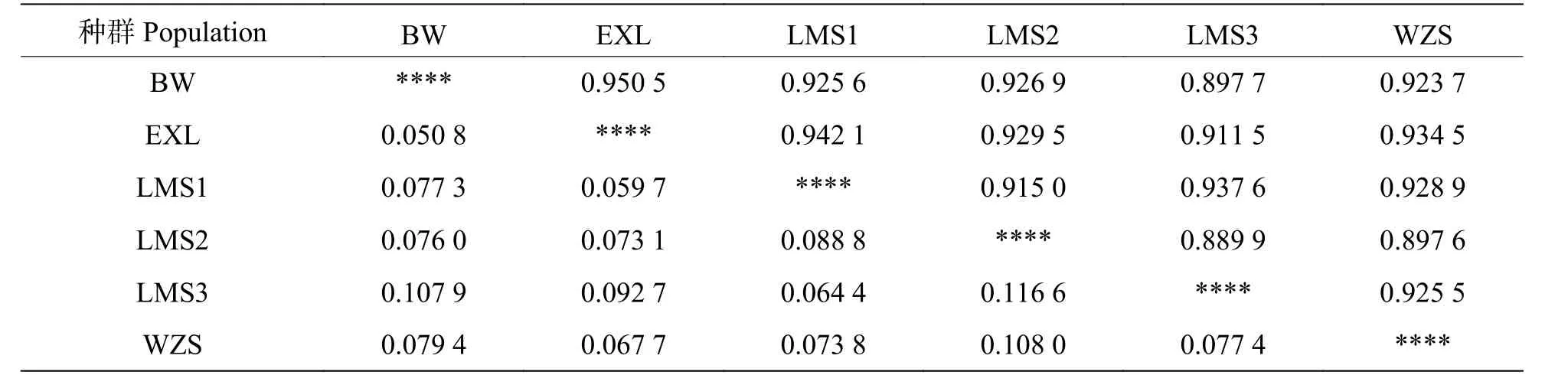
表5 基于SRAP 6 個(gè)鳥(niǎo)巢蕨種群間的遺傳相似度和遺傳距離Tab.5 Genetic similarity and genetic distance among the 6 populations of Asplenium nidus based on SRAP
SRAP 標(biāo)記對(duì)6 個(gè)種群鳥(niǎo)巢蕨野外種群遺傳距離的UPGMA 聚類(lèi)分析結(jié)果表明,海南島6 個(gè)鳥(niǎo)巢蕨種群可分為三大類(lèi):由圖3 可得,地理位置較近的BW 和EXL 為第I 類(lèi);同一地區(qū)的LMS1 和LMS3聚類(lèi)在了一起為第II 類(lèi);而LMS2 與其余種群為第Ⅲ類(lèi),其遺傳距離最大,遺傳分化也最顯著。
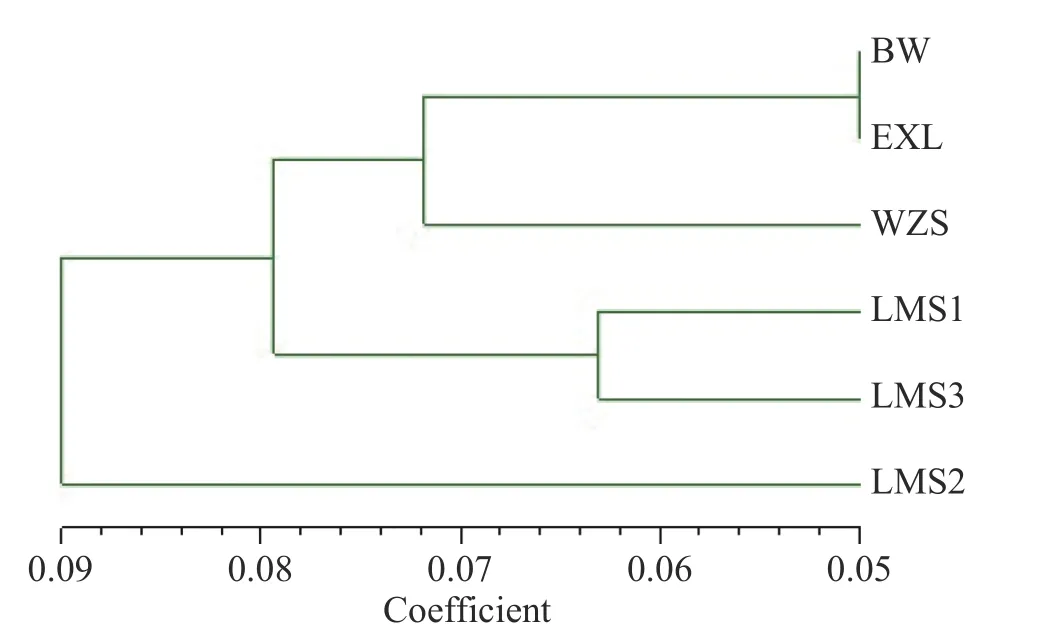
圖3 海南島鳥(niǎo)巢蕨6 個(gè)種群Nei’s 遺傳距離UPGMA 聚類(lèi)圖Fig.3 Dendrogram of the six natural populations of Asplenium nidus in Hainan Island based on Nei ’s makers using UPGMA cluster analysis
3 討 論
3.1 鳥(niǎo)巢蕨自然種群遺傳多樣性遺傳多樣性是生物多樣性重要組成部分,是生態(tài)系統(tǒng)多樣性和物種多樣性的基礎(chǔ)[16?17],也是分析物種進(jìn)化潛力和未來(lái)命運(yùn)重要方向[18]。遺傳多樣性大小是物種長(zhǎng)期進(jìn)化的產(chǎn)物,是其生存適應(yīng)和發(fā)展進(jìn)化的前提[20]。遺傳多樣性越高的物種對(duì)環(huán)境變化適應(yīng)能力就越強(qiáng),越容易在其分布范圍內(nèi)擴(kuò)展和開(kāi)拓新生境[20?21]。
本研究結(jié)果表明,海南島野生鳥(niǎo)巢蕨種群具有較高遺傳多樣性,多態(tài)位點(diǎn)百分比(PPB)在76.09%(LMS2 種群)~94.57%(LMS3 種群)之間,平均為85.15%。該結(jié)果與潘麗芹等[22]的荷葉鐵線蕨自然居群的遺傳多樣性研究和董元火等[23]的水蕨的生境及其遺傳多樣性分析結(jié)果相似,但與黃慶陽(yáng)等[24]對(duì)香鱗毛蕨種質(zhì)資源遺傳多樣性的AFLP 分析研究結(jié)果存在一定差異。原因可能是研究材料地理分布差異較大,或使用的分子標(biāo)記技術(shù)不同所導(dǎo)致。本研究中,野生鳥(niǎo)巢蕨種群間生境差異不大,但隨著海拔增加,氣候、土壤、植被等也隨之發(fā)生了明顯變化,因而造成6 個(gè)種群較高遺傳多樣性。其中,LMS3 地區(qū)遺傳多樣性最高(Ne、He、Ho分別為1.533 2、0.322 5、0.488 1),可能黎母山(LMS)作為海南三大江河(南渡江、萬(wàn)泉河、昌化江)的發(fā)源地,石灰?guī)r溝谷較多,符合鳥(niǎo)巢蕨對(duì)生境要求。加上黎母山林區(qū)屬熱帶季風(fēng)氣候,季風(fēng)帶來(lái)長(zhǎng)期潮濕更有利于鳥(niǎo)巢蕨孢子傳播和更新,而霸王嶺樣地鳥(niǎo)巢蕨種群遺傳多樣性最小(Ne、He、Ho分別為1.479 9、0.282 2、0.422 7);有兩個(gè)方面的原因:一是所采集個(gè)體的海拔分布較低,受人為干擾較重;另一原因可能是鳥(niǎo)巢蕨分布地受季風(fēng)影響較小,不利于孢子傳播和附著。本研究結(jié)果表明,在海南島6 個(gè)鳥(niǎo)巢蕨種群中LMS3 鳥(niǎo)巢蕨種群內(nèi)部遺傳變異最多,而B(niǎo)WL 鳥(niǎo)巢蕨種群內(nèi)部遺傳變異則相對(duì)較少。
3.2 種群的遺傳變異和遺傳結(jié)構(gòu)遺傳結(jié)構(gòu)指種群中遺傳變異分布的時(shí)空格局,其在基因流、基因突變和自然選擇的共同作用下形成,也與物種進(jìn)化史和生物學(xué)特性有關(guān)[25?26]。
鳥(niǎo)巢蕨雖然在種群和物種水平均具有較高的遺傳多樣性,但相比較而言,種群水平遺傳多樣性要低于物種水平,與種群間具有較高基因流(Nm=1.664 2,Nm>1)有關(guān),使漂變成為劃分種群遺傳結(jié)構(gòu)主導(dǎo)因素,表明鳥(niǎo)巢蕨距離較遠(yuǎn)種群間也存在一定程度基因交流[27?28]。遺傳分化程度Gst為0.130 6,在0.05~0.15 之間,屬于中等程度遺傳分化。
基于SRAP 標(biāo)記得出6 個(gè)鳥(niǎo)巢蕨野生種群多態(tài)位點(diǎn)百分率、Shannon 信息指數(shù)(Ho)、Nei’s 基因多樣性(He)、種群內(nèi)遺傳多樣性指數(shù)和總遺傳多樣性指數(shù),通過(guò)AMOVA 分析結(jié)果顯示總的種群間變異為11%,說(shuō)明鳥(niǎo)巢蕨種群內(nèi)存在豐富的遺傳變異,6 個(gè)鳥(niǎo)巢蕨野生分布種群間均有較高遺傳變異,黎母山種群與其他5 個(gè)種群遺傳變異最明顯。蔣速飛等[29]發(fā)現(xiàn),豐富的遺傳變異可能是物種具有較高遺傳多樣性的原因之一 。由于鳥(niǎo)巢蕨是孢子繁殖,孢子不易于長(zhǎng)距離遷徙,易造成地理隔離,推測(cè)地理隔離可能是造成鳥(niǎo)巢蕨種群間遺傳分化較大原因,而較高遺傳變異可能是本研究中各鳥(niǎo)巢蕨種群間遺傳多樣性較高的原因之一[30]。
3.3 鳥(niǎo)巢蕨自然種群保護(hù)物種遺傳多樣性水平能反映其適應(yīng)能力和進(jìn)化潛力,還有助于物種稀有或?yàn)l危原因及過(guò)程的探討[31]。對(duì)珍稀瀕危物種保護(hù)方針和措施的制定,如采樣策略遷地或就地保護(hù)選樣等都有賴(lài)于人們對(duì)物種遺傳多樣性認(rèn)識(shí)[32?33]。鳥(niǎo)巢蕨靠孢子繁殖,而孢子適宜在潮濕的環(huán)境生長(zhǎng),海南島氣候十分適宜鳥(niǎo)巢蕨繁殖生長(zhǎng),本研究也證實(shí)海南島鳥(niǎo)巢蕨具較高遺傳多樣性,說(shuō)明在本研究所選區(qū)域現(xiàn)階段人為活動(dòng)對(duì)鳥(niǎo)巢蕨的數(shù)量和遺傳多樣性影響較小,各區(qū)域還有一定數(shù)量鳥(niǎo)巢蕨種群分布。
值得注意的是,一旦生境遭到破壞會(huì)直接導(dǎo)致孢子不能傳播和存活,鳥(niǎo)巢蕨原生基因就會(huì)喪失,其遺傳多樣性也會(huì)隨之降低。建議鳥(niǎo)巢蕨保護(hù)應(yīng)以就地保護(hù)為主,可選擇鳥(niǎo)巢蕨種群遺傳多樣性較高的黎母山保護(hù)區(qū)區(qū)域(如本研究的黎母山LMS3 區(qū)域)建立級(jí)別更高的自然保護(hù)區(qū),加大保護(hù)和自然恢復(fù)力度,擴(kuò)大保護(hù)邊界。針對(duì)鳥(niǎo)巢蕨孢子不易于長(zhǎng)距離遷徙導(dǎo)致不同種群之間遺傳分化較大這一現(xiàn)象,可對(duì)不同自然群體中鳥(niǎo)巢蕨個(gè)體遷地保護(hù),結(jié)合采用重引入技術(shù)進(jìn)行自然居群恢復(fù),提高種群競(jìng)爭(zhēng)力和多樣性,擴(kuò)大群體間基因交流,最大限度保障鳥(niǎo)巢蕨遺傳多樣性,促進(jìn)熱帶雨林國(guó)家公園建成。
猜你喜歡
課堂內(nèi)外·初中版(科學(xué)少年)(2025年1期)2025-02-28 00:00:00
課堂內(nèi)外·初中版(科學(xué)少年)(2025年2期)2025-02-28 00:00:00
課堂內(nèi)外·初中版(科學(xué)少年)(2024年12期)2024-12-02 00:00:00
英語(yǔ)世界(2023年10期)2023-11-17 09:18:18
中學(xué)生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(shuō)(2021年8期)2021-11-03 10:55:16
學(xué)苑創(chuàng)造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(shù)(快樂(lè)歷史地理)(2020年9期)2020-03-19 05:10:56
科學(xué)大眾(中學(xué))(2019年3期)2019-05-17 10:04:30
汽車(chē)觀察(2018年10期)2018-11-06 07:05:26
