基于轉錄組學的不明原因復發性流產關鍵基因及機制分析
2021-04-19 03:35:34肖潤穎肖建華王比男李寒梅郭海春莫鴻英黎小弟任懂平柳紅艷
中南醫學科學雜志 2021年2期
關鍵詞:分析
肖潤穎,肖建華,王比男,李寒梅,郭海春,莫鴻英,黎小弟,任懂平,付 梟,柳紅艷,黎 麗,
(1.南華大學衡陽醫學院病原生物學研究所,湖南省衡陽市 421001;2.長沙市婦幼保健院,湖南省長沙市 410007;3.優生貝(北京)生物技術有限公司,北京市 102206)
復發性流產(recurrent spontaneous abortion,RSA)是指和同一性伴侶存在連續3次及以上、主要發生在孕28周之前的自然流產(spontaneous abortion,SA)[1]。RSA病因復雜,其潛在病因包括生殖系統解剖結構異常、內分泌代謝疾病、母體凝血功能異常、自身抗體的產生及感染等[2-3]。但40%~60%的RSA患者不屬于以上任何原因,確切原因尚不清楚,故稱不明原因復發性流產(unexplained recurrent spontaneous abortion,URSA)[4]。有研究表明,成功的妊娠需要母體和胚胎之間保持動態的免疫平衡,一旦平衡狀態被打破,則可能導致母體對胎兒組織的排斥反應,引起流產或者胚胎停止發育[5]。
探究URSA相關的基因表達譜和關鍵信號通路有助于臨床尋找新的治療靶點或標記物,為此研究者們進行了許多嘗試。Strug等[6]通過對子宮內膜修復部位組織的轉錄組測序(RNA-sequencing,RNA-seq),發現URSA的發生與子宮內膜γ干擾素(interferon γ,IFN-γ)等炎癥信號通路上調影響內膜的修復有關,且轉錄因子RBPJ可能是其關鍵調節因子。S?ber等[7]對早孕胎盤絨毛組織進行轉錄組測序,發現差異表達基因(differentially expressed genes,DEGs)主要參與DNA的復制、RNA的轉錄及加工、線粒體功能等,最關鍵的是大部分DEGs都受E2F/DP1轉錄因子的調節,且與不良妊娠結局發展或胎盤功能相關。Gu等[8]和Zhang等[9]發現了白細胞介素-10(interleukin-10,IL-10)和白細胞介素-1β(interleukin-1β,IL-1β)基因啟動子區相關位點存在多態性變異與URSA的發生相關。然而,目前對這些機制的認識仍有許多疑問。
本研究采用RNA-seq對URSA患者與正常妊娠蛻膜組織轉錄組基因進行分析,運用高密度單核苷酸多態性(single nucleotide polymorphisms,SNP)芯片檢測關鍵基因的位點多態性變異,從而探索母胎界面微環境的基因表達譜改變及與URSA發生相關的關鍵基因及遺傳因素,為URSA的分子免疫機制研究和臨床預防、診斷及治療提供一定參考依據。
1 資料和方法
1.1 一般資料
本研究納入長沙市婦幼保健院2018年4月—2020年10月的15例患者;其中6例不明原因復發性流產孕婦納入URSA組,9例既往無流產史且無其他異常的人工流產孕婦納入對照組。URSA組入組標準:年齡21~40歲,體質量指數(body mass index,BMI)19~24 kg/m2,孕周≤12周,連續自然流產次數≥3次。排除標準:生殖道解剖結構異常,內分泌、血栓史、感染、抗心磷脂抗體、染色體異常,孕期已接受黃體酮等藥物治療。對照組入組標準:年齡21~40歲,BMI 19~24 kg/m2,孕周≤12周,妊娠狀況良好,既往無胚胎停育史,有一次以上正常生產史且子女健康者[3]。本研究經長沙婦幼保健院醫學倫理學委員會批準,并獲得患者知情同意。
1.2 樣本采集和預處理
選取URSA組4例及對照組3例孕婦,獲取其蛻膜組織,放入RNA組織保存液中充分滲透后(康為世紀,貨號:CWY042)迅速冷凍,并儲存在液氮中,以備后續RNA-seq檢測。URSA組及對照組所有患者各取2 mL外周血于一根EDTANa2一次性負壓采血管中,上下輕柔混勻,防止血液凝固,然后轉移到樣本凍存管中,-20 ℃凍存,待DNA提取和高密度SNP芯片檢測用。
1.3 RNA測序
提取1.2中蛻膜組織總RNA。對所提取的RNA完成質量檢測(其中對照組1例RNA樣本不合格,棄去),質量檢測合格后進行下一步的mRNA富集、片段化及cDNA合成。然后通過末端修復、加接頭及擴增等步驟,完成文庫的構建。文庫質量檢測合格后上機進行測序,測序及生物信息分析由優生貝(北京)生物技術有限公司完成。
1.4 測序數據分析
原始下機數據經過過濾及質量控制后得到Clean Reads,然后通過軟件HISAT2(網址http://ccb.jhu.edu/software/hisat2/index.shtml)比對至人類參考基因組(版本:GRCh38),且需滿足對比到人類參考基因組的唯一比對率均超過90%的標準。根據|log2(Fold Change)|>1,P<0.05的原則篩選出DEGs進行GO和KEGG分析。并將DEGs比對到STRING蛋白質相互作用數據庫,構建蛋白相互作用網絡(protein-protein interaction network,PPI network),找出關鍵節點基因并分析其可能的生物學功能。利用GSEA 3.0軟件(網址http://www.gsea-msigdb.org/gsea/index.jsp)進行基因集富集分析(gene set enrichment analysis,GSEA),富集標準為:|NES|>1,NOM p-val<0.05,FDR q-val<0.25;NES的正負分別指示基因集的升高和降低(NES:標準化富集分數;FDR q-val:假陽性率;NOM p-val:標準化P值)。
1.5 高通量SNP芯片測序
采集URSA組和對照組外周血共15例,提取外周血DNA,使用高密度SNP芯片(ASA-CHIA,illumine,USA)對DNA樣本進行單核苷酸多態性基因型檢測,使用iScan平臺對SNP芯片進行掃描分析,采用Bioconductor 軟件(版本號:v3.12,網址http://www.bioconductor.org/)對獲取的下機數據進行分析。操作簡述為:將質檢合格的DNA樣品移入MSA3平板上,經片段化處理、恒溫擴增、沉淀及重懸后與芯片孵育雜交,清洗后進行單堿基延伸及染色操作后上機分析。使用iScan(Illumina,美國)對芯片進行掃描及分析。所有實驗操作流程均嚴格按照廠家標準操作說明書進行。芯片數據分析軟件為 GenomeStudio(Illumina,美國,網址https://support.illumina.com/array/array_software/genomestudio/downloads.html)。
1.6 統計學方法
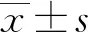
2 結 果
2.1 兩組患者臨床特征比較
兩組患者年齡、身高、體質量、BMI、孕周、入院HCG及入院孕酮水平比較,差異均無統計學意義。

表1 兩組患者臨床特征比較
2.2 相關性分析與差異表達基因的篩選
兩組數據的主成分分析(principal component analysis,PCA)結果如圖1所示,URSA組(n=4)和對照組(n=2)在轉錄水平能得到了較好的線性分離。以|log2(Fold Change)|> 2,P<0.05為篩選標準,共獲得1 057個差異基因。與對照組相比,URSA蛻膜組織中上調表達基因469個,下調表達基因588個。由此構建URSA患者基因差異表達熱圖(圖2)。

圖1 蛻膜組織RNA測序數據主成分分析U1~U4為URSA組4例樣本;C1~C2為對照組2例樣本。

圖2 DEGs等級聚類圖紅色表示基因表達量高,藍色表示基因表達量低。U1~U4為USRA組4例樣本;C1和C2為對照組2例樣本。
2.3 差異表達基因富集分析
GO富集分析結果如圖3所示,DEGs在生物過程中主要富集于T細胞活化與分化、白細胞趨化性、免疫應答過程的負調控及Toll樣受體信號通路等(P<0.05);在分子功能中主要富集于受體配體活性、抗原肽與主要組織相容性復合體Ⅱ類分子(major histocompatibility complex class Ⅱ,MHC-Ⅱ)結合活性等(P<0.05);在細胞成分中主要富集于細胞外基質、MHC蛋白復合物、MHC-Ⅱ類蛋白復合物、分泌性顆粒膜等(P<0.05)。KEGG富集分析結果如圖4所示,DEGs主要富集于吞噬體、金黃色葡萄球菌感染、抗原加工和遞呈、自身免疫病、同種異體移植排斥反應、造血細胞譜系、補體和凝血級聯反應及Toll樣受體信號通路等(P<0.05)。這些DEGs主要與炎癥反應、異常免疫應答、母體高凝狀態有關,不利于母胎免疫耐受的維持。

圖3 差異基因GO富集分析

圖4 差異基因KEGG富集分析圓形面積代表該信號通路富集的DEGs數量,面積越大DEGs數量越多;P.adjust由藍到紅色的過度色表示P校正值由大到小。
RNA測序數據進行GSEA通路富集分析結果如圖5所示,富集基因集參與的主要通路有IL-2/STAT5通路、PI3K/Akt通路、IL-10抗炎信號通路、絲裂原活化蛋白激酶(mitogen activated protein kinase,MAPK)通路的負反饋調節、轉化生長因子-β1(transforming growth factor β1,TGF-β1)通路、FOXP3信號通路的激活等,且這些信號通路均與Treg細胞活化及免疫抑制功能的發揮密切相關。以上信號通路的下調可能會影響Treg細胞的活化及免疫抑制功能發揮,使得URSA患者母胎界面的免疫平衡被打破,造成不良妊娠結果。

圖5 GSEA基因集富集分析圖
2.4 PPI網絡分析
其中貢獻度排名前24的節點基因有CGB8、CGB5、MUC5AC、GPR183、SLC5A8、FOXS1、CD180、ALPP、HLA-DRB5、CXCL9、MMP27、UST、TUBA3D、SIGLEC14、STX11、BAMBI、HLA-DRB1、SLC2A12、PPP1R3C、SCGB3A1、SHISA3、SLC10A6、CD300C、XCL1等(圖6)。

圖6 PPI網絡分析圖圓代表蛋白質節點,直線代表蛋白質之間相互作用,線段粗細代表相互作用強弱。
2.5 SNP芯片檢測結果
使用高密度SNP芯片對URSA組(n=6)和對照組(n=9)關鍵差異基因HLA-DPA1、IL-2R、CD80、PTPRC、IRF4、CTLA4、XCL1、PDCD1、DUSP1、MAFB、TIGIT、IL-10、TRIB1的位點多態性變異進行檢測。結果顯示僅HLA-DPA1 基因多態性位點(rs1042866、rs2567279)和IL-2RA基因多態性位點(rs2025346)的等位基因頻率在兩組間的分布差異有統計學意義(表2,P<0.05)。且HLA-DPA1 rs1042866位點C→T的突變可能減低URSA的發病風險,而HLA-DPA1 rs2567279位點T→C的突變及IL-2RA rs2025346位點G→A的突變則可能提高URSA的發病風險。其余關鍵基因SNP的差異無統計學意義(P>0.05)。

表2 SNP基因芯片篩選出的關鍵基因位點分布
3 討 論
RSA為常見的產科病癥,其患者及伴侶均承受了很大的身體、心理壓力。其中URSA的具體機制尚不明確,臨床的各種治療手段也無法取得讓人滿意的療效[10],因此探究URSA病因,找到行之有效的診療方法已經成為目前婦產及生殖醫學上急需解決的難題之一。目前認為URSA發生主要與免疫因素相關,但其內在的免疫調控機制及關鍵因素尚未明確[11]。
本研究采用RNA-seq對URSA與正常妊娠蛻膜組織的基因表達譜進行檢測,結果顯示:與對照組比較,URSA蛻膜組織中有1 057個差異表達基因(DEGs),其中上調469個,下調588個。對DEGs進行GO、KEGG、PPI及GSEA等生物信息學分析。GO分析顯示,DEGs主要富集于T細胞分化與活化、白細胞趨化性、抗原肽與MHC-Ⅱ類分子結合活性及免疫應答過程的負調控。KEGG分析顯示,DEGs主要與自身免疫病、補體和凝血級聯反應及Toll樣受體信號通路等相關,這提示URSA的發生可能與炎癥信號通路的激活、母體對胚胎免疫耐受狀態的打破以及母體處于高凝狀態相關。為進一步明確URSA患者蛻膜局部功能改變的機制,對DEGs進行GSEA通路富集分析,發現URSA患者蛻膜組織的IL-2/STAT5通路、PI3K/Akt通路、IL-10抗炎信號通路、MAPK通路、TGF-β1通路、FOXP3蛋白激活通路等富集且表達下調。上述信號通路均與Treg細胞密切相關,這些通路的下調可能影響Treg細胞的活化及免疫抑制功能。Treg細胞是一群免疫抑制性T細胞,對妊娠的維持及母體耐受均起重要作用。其主要功能包括消耗IL-2,抑制NK細胞或殺傷性T細胞的殺傷作用,以及表達FOXP3、TGF-β、IL-10等免疫抑制性分子[12-13]。
綜合PPI及GSEA分析挑選出的核心DEGs有HLA-DPA1、IL-2RA、CD80、CTLA4、PDCD1、IL-10、PTPRC、IRF4、XCL1、DUSP1、MAFB、TIGIT及TRIB1,這些基因可能影響Treg細胞的功能,與URSA發生有關。HLA-DPA1屬于HLA-Ⅱ類α鏈旁系同源基因。能夠遞呈外源性抗原肽,在免疫應答中發揮重要作用[14]。Shima等[15]發現,小鼠在孕前幾個小時內就可以在陰道黏液中發現父系抗原和母系MHC-Ⅱ類細胞,可誘導CD4+CD25+FoxP3+Tregs的增殖,促進對父系同種抗原的耐受。IL-2RA基因(即CD25)的突變與白細胞介素-2受體α缺乏有關。CD25為Treg細胞表面的關鍵分子標志,IL-2是啟動和維持Treg細胞增殖和分化的重要細胞因子,而Treg細胞表面的CD25分子能通過高效結合IL-2從而耗盡免疫微環境中的IL-2,維持母胎免疫耐受[12]。CD80(即B7)通過與T細胞表明的CD28或CTLA-4的結合而促進或抑制T細胞活化及細胞因子產生。PDCD1(即PD-1)是活化的T細胞表達的一種免疫抑制受體,能抑制T細胞的活化,還可以促進CD4+T細胞向Treg細胞分化,PD-1基因的下調與自身免疫病有關[16]。IL-10是Treg細胞、Th2細胞分泌的能調節免疫反應的最為重要的妊娠保護性細胞因子之一,該分子下調則不利于母胎界面免疫耐受的維持。PTPRC基因編碼的蛋白是T細胞和B細胞抗原受體信號轉導的調節因子,能夠抑制JAK激酶從而抑制IL-2/STAT5信號通路[17]。IRF4基因編碼蛋白是Ⅰ型干擾素基因表達所必需的轉錄因子之一[18]。XCL1是趨化因子超家族成員,對T細胞具有特異性趨化作用[19]。DUSP1編碼的蛋白能使MAPK1/ERK2激酶去磷酸化,對細胞的增殖起負調控作用[20]。MAFB是一種轉錄因子,在調節特異性造血細胞譜系中起重要作用。TIGIT能輔助調節T細胞對B細胞的應答[21]。TRIB1主要參與脂質代謝,影響M2型巨噬細胞及Treg的分化等[22]。綜上,共篩到的13個關鍵基因均與Treg細胞密切相關,這些基因的表達下調可能影響了Treg細胞分化及免疫抑制功能的發揮。
運用高密度SNP芯片檢測兩組DNA樣本的基因位點多態性變異,發現除HLA-DPA1及IL-2RA外,其他的關鍵DEGs位點多態性在兩組間的差異無統計學意義(P>0.05)。HLA-DPA1 rs1042866、HLA-DPA1 rs2567279及IL-2RA rs2025346等位基因頻率分布間差異有統計學意義(P<0.05)。HLA-DPA1 rs1042866位點C→T的突變可能減低URSA的發病風險,而HLA-DPA1 rs2567279位點T→C的突變及IL-2RA rs2025346位點G→A的突變則可能提高URSA的發病風險。本研究為URSA發病機制的探究提供了新的思路和新靶點基因。
綜上所述,本研究通過比較URSA組及正常妊娠組蛻膜組織的差異表達基因,發現了1057個差異表達基因。在這些差異基因中有下調Treg細胞活化與功能的相關基因及促進炎癥反應和免疫應答的相關基因,它們可能使URSA患者母胎界面的免疫平衡被打破,造成不良妊娠結果。在蛋白質相互作用的關鍵節點基因中,發現了3個新的與URSA相關的關鍵基因DUSP1、MAFB、TIGIT,它們的表達下調可能影響Treg細胞的活化及功能發揮。SNP芯片檢測顯示在URSA患者中存在HLA-DPA1 rs1042866、rs2567279及IL-2RA rs2025346基因位點多態性變異,可能影響URSA的發生。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
