宣恩火腿發酵過程中表面微生物群落演替規律
2021-04-27 08:33:44鄧祥宜李繼偉何立超張原源黃國威鮑曉龍邱朝坤
食品與發酵工業 2021年7期
關鍵詞:物種
鄧祥宜,李繼偉,何立超,張原源,黃國威,鮑曉龍,邱朝坤
(武漢設計工程學院 食品與生物科技學院,湖北 武漢,430205)
宣恩火腿是湖北省恩施自治州一帶特產,其在乾隆時期曾作為貢品,以宣恩縣所產火腿最負盛名,甚至被譽為中國“四大名腿”之一[1-2]。宣恩火腿一直沿用傳統工藝生產,微生物自然接種,在長達1~2年的加工過程中,形成了獨特的質地和風味。關于宣恩火腿發酵過程中蛋白質、脂肪等理化指標的變化規律已有報道[2-4],但宣恩火腿發酵過程中微生物群落的變化規律有待揭示。
研究微生物群落的方法主要包括分離培養法[5]、Biolog-ECO微平板技術[6]、變性梯度凝膠電泳法[7]、16S rRNA/18S rRNA/ITS基因克隆文庫測序法[8]和高通量測序法等。高通量測序技術,又稱下一代測序技術,通過測定細菌16S rRNA序列、真菌ITS序列等,與對應的物種分類數據庫進行比對,從而確定微生物種類和豐度[9-11];其雖然不能獲得純培養菌株,但一次可測得單個樣本中數萬條菌株序列信息,故能以更高的靈敏度、準確度和相對低廉的價格快速鑒定樣本中的微生物種類和豐度,包括豐度很低的種類[12-13]。
微生物在火腿發酵過程中扮演著重要角色,與火腿的風味、品質和安全性密切相關,研究火腿發酵過程中微生物群落組成具有很強的實際意義。本文利用高通量測序技術對不同發酵期宣恩火腿表面的細菌、真菌群落多樣性進行研究,以期為宣恩火腿建立工業化生產工藝作鋪墊。
1 材料與方法
1.1 材料及取樣
不同發酵期宣恩火腿表面(肉面)微生物樣本,于2019年11月湖北省思樂牧業集團有限公司采樣。宣恩火腿加工工藝流程[4, 14]:選腿(豬后腿)→修胚→攤涼→腌制→洗腿→整形→烘腿→入庫發酵→洗霉→修割→驗收。自“入庫發酵”開始計算發酵時間,取樣點為發酵前期(發酵40 d,霉菌生長完全覆蓋火腿表面)、發酵中期(發酵90 d,霉菌生長最旺盛階段)、發酵后期(發酵570 d,霉菌生長消退;發酵180 d以上為發酵后期,但發酵1~2年能更好地保證火腿中病毒失活,增加安全性和品質[15])。采樣參照GB 4789.1—2016《食品安全國家標準 食品微生物學檢驗 總則》規定,各階段樣本分別隨機選取20個面積約5 cm×5 cm的取樣區(肉面),用滅菌棉球反復擦拭火腿表面(肉面)微生物,并清洗到100 mL無菌水中,而后轉移至無菌離心管,6 000 r/min離心15 min,取沉淀備用。
1.2 主要儀器與試劑
GeneAmp?9700型PCR儀,美國ABI公司;QuantiFluorTM-ST藍色熒光定量系統,美國Promega公司;Illumina MiSeq測序平臺、TruSeqTM DNA Sample Prep Kit,美國Illumina 公司;FastDNA?SPIN Kit for Soil土壤DNA快速提取試劑盒,美國MPbio公司;TransStart Fastpfu DNA 聚合酶,北京TransGen生物;AxyPrepDNA凝膠回收試劑盒,美國Axygen公司。
1.3 實驗方法
1.3.1 微生物基因組DNA的提取及高通量測序
取沉淀樣本,用土壤DNA快速提取試劑盒提取總DNA,1%瓊脂糖凝膠電泳檢測后,用NanoDrop 2000 測定DNA純度和含量。通過PCR擴增細菌16S rRNA V3~V4區,引物為338F(5′-ACTCCTACGGGAGGCAGCAG-3′)、806R(5′-GGACTACHVGGGTWTCTAAT-3′),擴增真菌ITS1-ITS2序列,引物為ITS1F(5′-CTTGGTCATTTAGAGGAAGTAA-3′)、ITS2R(5′-GCTGCGTTC-TTCATCGATGC-3′);PCR產物經電泳、回收、定量分析后進行Illumina MiSeq測序[16],測序公司為上海美吉生物醫藥科技有限公司。
1.3.2 高通量測序結果分析
MiSeq高通量測序結果借助美吉生物云平臺進行分析,主要分析流程為:將測序原始序列進行拼接和質控,按樣本進行操作分類單元(operational taxonomic unit,OTU)聚類分析和物種分類學分析,對數據進行抽平處理;基于抽平后的OTU進行Alpha多樣性分析;基于分類學信息,在不同分類學水平上進行群落結構統計分析;并對細菌、真菌優勢種進行物種相關性網絡分析、系統發育分析(即進化分析);并對群落功能進行預測。
OTU分析使用Usearch平臺(version 7.0, http://drive5.com/uparse/),相似性在97%以上的序列聚為一類,細菌比對Silva數據庫(Release132, http://www.arb-silva.de),真菌比對Unite數據庫(Release 7.2, http://unite.ut.ee/index.php);物種相關性網絡分析使用Networkx軟件;系統發育分析使用FastTree軟件(version 2.1.3, http://www.microbesonline.org/fasttree/);功能預測分析使用PICRUSt軟件包(version 1.1.0, http://picrust.github.io/picrust/),細菌使用EggNOG(evolutionary genealogy of genes:Non-supervised Orthologous Groups,http://eggnog.embl.de/)數據庫,真菌使用MetaCyc(https://metacyc.org/)數據庫;樣本與物種關系Circos圖用Circos-0.67-7(http://circos.ca/)完成,其他圖使用Excel軟件、R語言工具統計和作圖。
1.3.3 高豐度未知物種的進一步注釋
高通量測序結果分析中,一些物種的分類無法確定(no_rank/unclassified),挑選其中高豐度OTU代表序列,在線比對NCBI中的16S rRNA或ITS數據庫(如果沒有高度相似序列,則進一步比對nr數據庫),并基于查詢的16S rRNA或ITS序列,用MEGA7軟件通過最大似然法構建系統發生進化樹,以對物種進行更精確的注釋(Bootstrap值設置為1 000次重復)[17]。
2 結果與分析
2.1 樣本序列統計
宣恩火腿樣本測序信息見表1。經質控、拼接,細菌(含古菌)16S rRNA序列V3~V4區共測得134 721條序列,平均長度426 bp;真菌ITS1-ITS2序列共測得111 395條序列,平均長度231 bp。按最小序列數抽平處理[18],獲得抽平后的序列數為細菌38 829條/樣本、真菌36 268條/樣本。
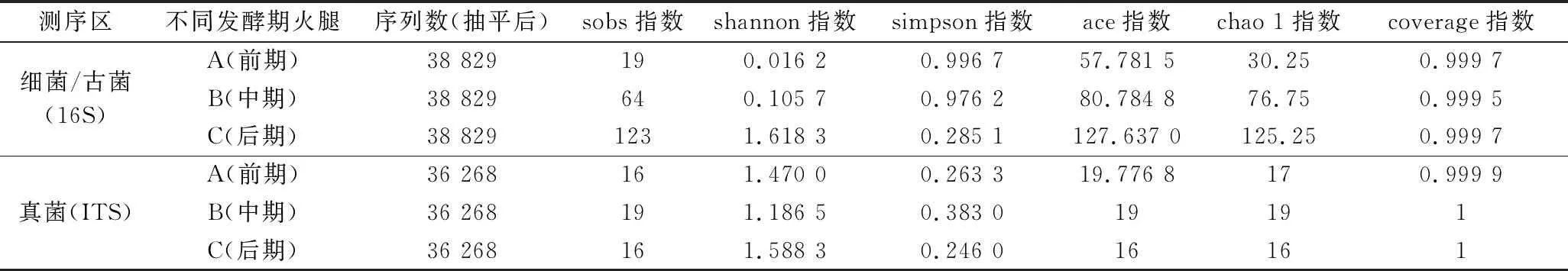
表1 樣本信息和Alpha多樣性指數Table 1 Sample information and Alpha diversity index
2.2 宣恩火腿微生物群落Alpha多樣性分析
宣恩火腿表面微生物群落Alpha多樣性指數見表1。多樣性指數中,sobs指數、ace指數和chao1指數可顯示群落物種的豐富度,數值越大代表物種豐富度越高;shannon指數和simpson指數綜合反映群落多樣性,shannon指數越大代表群落多樣性越高,simpson指數越小代表群落多樣性越高;coverage指數顯示物種覆蓋度,值越接近1(即100%)表明測序越全面,越能代表樣本真實物種組成情況[19-20]。
表1中,所有樣本真菌和細菌測序的覆蓋率(coverage)指數均大于99.9%,同時樣品稀釋曲線趨于平緩(圖1),說明增加測序量只能發現極少量的物種,即測序結果能很好地反應樣本微生物多樣性。宣恩火腿發酵過程中,細菌(含古菌)的sobs、ace和chao1指數持續增加,顯示物種豐富度不斷提高;shannon指數持續顯著增加,simpson指數持續顯著減小,顯示細菌多樣性持續提高。發酵過程中,真菌sobs、ace和chao1指數變化較小,顯示真菌物種豐富度變化相對較小;真菌的shannon指數先減小后增大,simpson指數先增大后減小,但變化幅度均較小,顯示真菌多樣性波動較小,呈現微弱的先減小后增大的趨勢。另外,比較sobs指數、ace指數和chao 1指數可發現:細菌物種豐富度整體大于真菌。因此,在發酵過程中,宣恩火腿表面細菌豐富度和多樣性持續增加,真菌的豐富度和多樣性波動較小,且整個發酵過程中細菌物種豐富度大于真菌。

圖1 樣品稀釋曲線Fig.1 Rarefaction curves of samples注:A-b、B-b、C-b依次為發酵前期、中期、后期火腿樣本的細菌群落;A-f、B-f、C-f分別為發酵前期、中期、后期火腿樣本的真菌群落
2.3 宣恩火腿微生物群落組成分析
2.3.1 細菌(含古菌)群落組成分析
在97%的相似水平上對宣恩火腿表面細菌16S rRNA V3~V4區測序結果進行OTU分析,通過Venn圖展示宣恩火腿發酵過程中共有和獨有的細菌(含古菌)物種數如圖2-a所示。所有樣本中共測得133個細菌OTUs,其中發酵前期、中期、后期的火腿樣本依次測得19、64、123個OTUs;說明宣恩火腿發酵過程中,細菌群落豐富度逐漸增加,不斷有新的細菌種類參與發酵過程。
在門水平上,所有樣本共檢出11個門的細菌,除1個豐度很低的古細菌門(Euryarchaeota,廣古菌門,占比<0.01%)外,其余10個為真細菌門,主要包括厚壁菌門(Firmicutes)、變形菌門(Proteobacteria)、擬桿菌門(Bacteroidetes)、放線菌門(Actinobacteria)等。發酵前、中期,厚壁菌門占絕對優勢(>99%);發酵后期,變形菌門占69%,而厚壁菌門下降到27%(圖2-b)。宣恩火腿表面細菌在屬水平(共96個)、種水平(共125個)上的組成分別見圖2-c和圖2-d。圖2-d為Circos圖,是一種描述樣本與物種之間對應關系的可視化圈圖,其不僅反映了每個樣本的優勢物種組成比例,同時也反映了各優勢物種在不同樣本(分組)中的分布比例[21]。發酵前、中期,葡萄球菌屬的木糖葡萄球菌(Staphylococcusxylosus)占據絕對優勢(>98%);發酵后期,沙雷氏菌屬(Serratia)未知種unclassified_g_Serratia(OTU61)占比最大,為42.2%,前期占優勢的木糖葡萄球菌下降到24.9%,根瘤菌科未知屬未知種s_uncultured_Rhizobiales_bacterium(OTU113)占20.8%,甲基菌屬的藤黃亞甲基桿菌(Methylobacteriumfujisawaense)占3.9%,Solitalea屬未培養菌s_uncultured_bacterium_g_Solitalea占1.9%(OTU72),芽孢桿菌科未知種s_unclassified_f_Bacillaceae占1.6%(OTU37),其他占4.7%。
綜上,宣恩火腿發酵中,細菌群落豐富度不斷增加,各物種相對豐度呈動態變化,發酵前、中期主要為木糖葡萄球菌,發酵后期的主要優勢屬為葡萄球菌屬、沙雷氏菌屬和甲基菌屬。
2.3.2 真菌群落組成分析
在97%的相似水平上對真菌ITS1-ITS2測序結果進行OTU分析,通過Venn圖展示宣恩火腿發酵過程中共有和獨有的真菌物種數目信息(圖2-f)。3個發酵期樣本中真菌群落共測得21個OTUs,其中發酵前、中、后期的火腿樣本依次測得16、19、16個OTUs。所有發酵期共有OTUs為12個,占57%;發酵前期沒有獨有OTUs,發酵中、后期分別獨有1、2個OTUs。因此,宣恩火腿發酵過程中真菌種類較為穩定,57%的種類在整個發酵期始終存在。
宣恩火腿真菌包括子囊菌門(Ascomycota)、擔子菌門(Basidiomycota)和未分類的門(unclassified_k_Fungi)。發酵過程中,子囊菌門占比持續增加,擔子菌門占比不斷減少,未分類門呈先增加后減少的趨勢(圖2-g)。宣恩火腿表面真菌在屬水平(共8個)、種水平(共12個)上的組成分別見圖2-h和圖2-e。曲霉屬(Aspergillus)在發酵過程中較為穩定,發酵前、中、后期占比依次為37.9%、43.2%和41.0%,但在種水平上有物種更替,發酵前、中期Aspergilluscibarius占優勢,而發酵后期帚狀曲霉(Aspergilluspenicillioides)占優勢;未分類真菌s_unclassified_k_Fungi(OTU7)在發酵前、中、后期占比依次為24.3%、46.5%和25.6%;節菌屬(Wallemia)在發酵過程中逐漸下降,發酵前、中、后期占比依次為37.6%、9.9%和1.5%;未分類酵母unclassified_o_Saccharomycetales(OTU20)由發酵前期未檢出、中期占0.01%,到發酵后期占31.1%,說明酵母是發酵后期優勢類群之一。
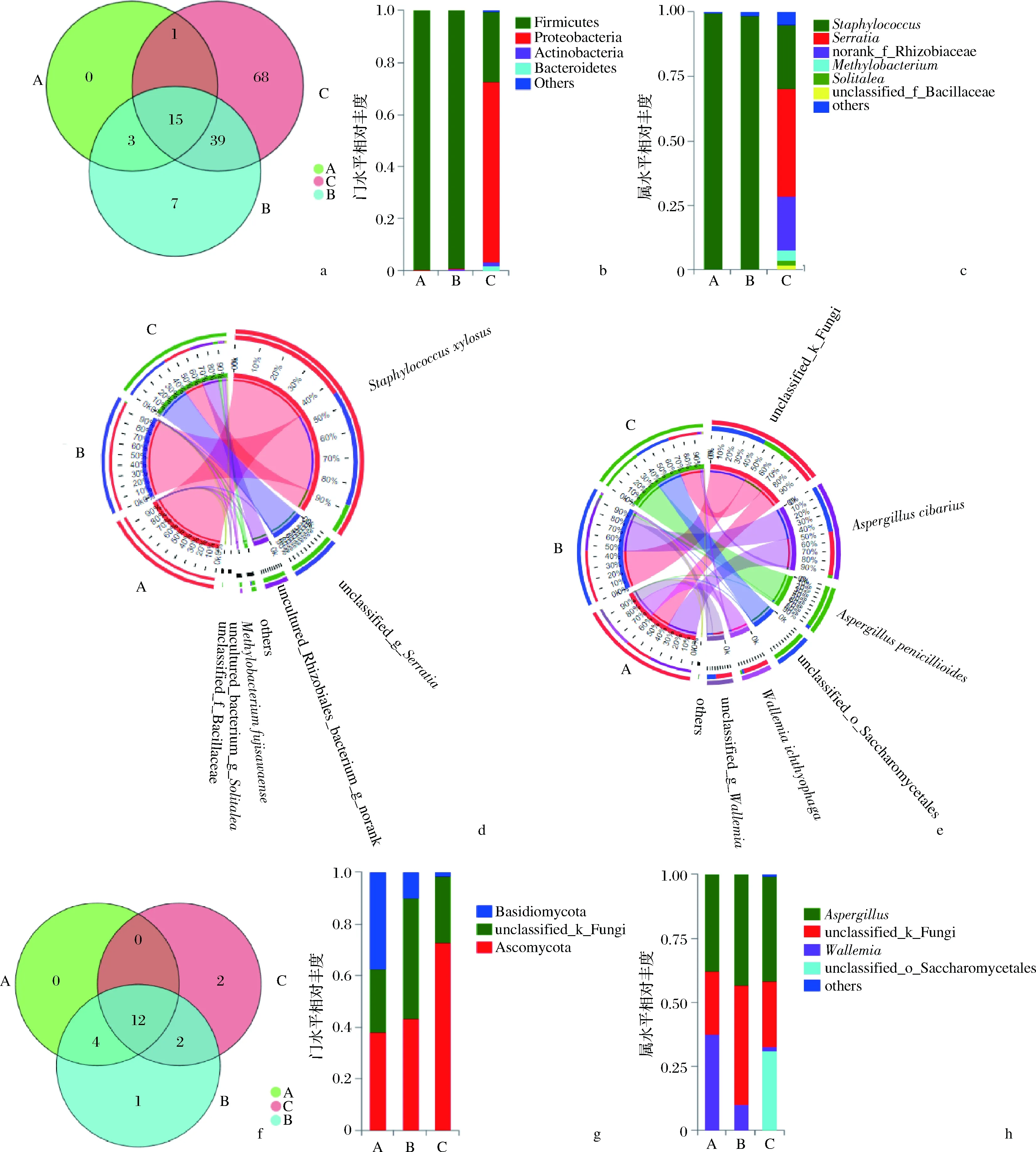
a-細菌OTU水平Venn圖;b-細菌門水平相對豐度;c-細菌屬水平相對豐度;d-細菌種水平circos圖;e-真菌種水平Circos圖;f-真菌OTU水平Venn圖;g-真菌門水平相對豐度;h-真菌屬水平相對豐度圖2 宣恩火腿的微生物區系Fig.2 The microbiota of Xuanen ham注:A、B、C依次為發酵前、中、后期的火腿樣本
綜上,宣恩火腿發酵過程中,真菌種類相對穩定,但各物種相對豐度存在動態變化,發酵前、中期主要是Aspergilluscibarius、未分類真菌(OTU7)、節菌屬占優勢,發酵后期主要優勢種為帚狀曲霉、未分類真菌(OTU7)、未分類酵母(OTU20)等。
2.4 物種相關性網絡分析
2.4.1 單因素網絡分析
對宣恩火腿表面細菌、真菌優勢種分別進行單因素網絡分析,當物種之間的相關系數符合某一閾值(P<0.05)時就用線連接,用不同顏色表示物種之間正、負相關性[22],結果見圖3-a和圖3-b。細菌群落中,木糖葡萄球菌與其他種類呈負相關,其他種之間互為正相關(圖3-a)。真菌群落中,曲霉屬Aspergilluscibarius和節菌屬s_unclassified_g_Wallemia呈正相關;這兩者與發酵后期占優勢的帚狀曲霉(A.penicillioides)、巴恩曲霉菌(A.baarnensis)、三角山氏酵母(Yamadazymatriangularis)和未分類酵母s_unclassified_o_Saccharomycetales(OTU20)均為負相關;后4種真菌互為正相關(圖3-b)。雖然發酵過程中,未分類真菌(OTU7)豐度一直很高,但它與其他種類真菌的豐度沒有顯著相關性,故未在圖3-b中顯示。
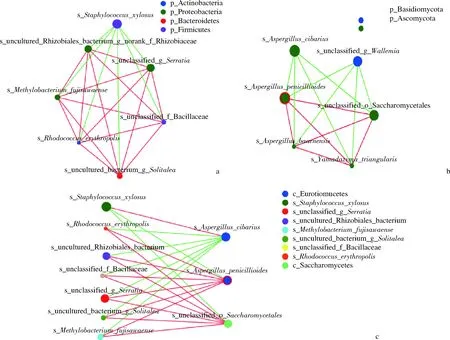
a-細菌種水平單因素網絡分析;b-真菌種水平單因素網絡分析;c-細菌與真菌種水平雙因素網絡分析圖3 宣恩火腿微生物相關性網絡分析Fig.3 Network analysis for microbiota of Xuanen ham注:節點大小與種的豐度成正比,紅色連線代表正相關,綠色連線代表負相關;a與b中節點顏色代表門類別,c中節點顏色代表綱類別
2.4.2 雙因素網絡分析
將主要細菌優勢種和真菌優勢種(作為其他因子)進行雙因素網絡分析,結果見圖3-c。曲霉屬Aspergilluscibarius與木糖葡萄球菌呈正相關,與其他細菌呈負相關;帚狀曲霉(A.penicillioides)、未分類酵母(OTU20)與木糖葡萄球菌呈負相關,與其他細菌呈正相關(圖3-c);其他主要真菌與細菌種類沒有顯著相關性,故未在圖3-c中顯示。
2.5 進化分析
在種水平上對宣恩火腿主要細菌和真菌分別進行進化分析,結果見圖4。主要細菌種類來自厚壁菌門、變形菌門、擬桿菌門和放線菌門等4個門(圖4-a);主要真菌種類來自子囊菌門、擔子菌門和未分類的門(圖4-c)。將OTU分析中沒有成功注釋到種的OTU代表序列,與NCBI數據庫中相似度最高的序列一起構建個性化系統發生樹(圖4-b、4-d),以對物種進行更精確的注釋。
細菌(圖4-b)中,根瘤菌科未知種uncultured_Rhizobiales_bacterium(OTU113)與Methylobacteriumbrachiatum和M.phyllostachyos聚為一類,16S rRNA序列一致度為100%,所以OTU113能進一步鑒定到甲基菌屬(Methylobacterium),可能為兩者之一;同理,芽孢桿菌科未知屬未知種unclassified_Bacillaceae(OTU37)可鑒定為Corynebacteriumtuberculostearicum,Solitalea屬未知種uncultured_bacterium_Solitalea(OTU72)可鑒定為Oceanobacilluspicturae,葉桿菌屬未知種uncultured_bacterium_Phyllobacterium(OTU84)可鑒定為Phyllobacteriummyrsinacearum。沙雷氏菌屬未知種unclassified_g_Serratia(OTU61)與未培養細菌序列KF632521聚為一類,序列一致度100%,其目前還不能鑒定到種;同理,腸桿菌科未知屬未知種unclassified_norank_ Enterobacteriaceae(OTU124)不能鑒定到屬。
真菌(圖4-d)中,未分類真菌unclassified_Fungi(OTU7)與未培養真菌ITS序列KF221761聚為一類,序列一致度99%,其目前還不能進一步鑒定到門。節菌屬未知種unclassified_Wallemia(OTU3、OTU4)分別與Wallemiasebi、Wallemiatropicalis一致度達99%,所以unclassified_Wallemia包括這2個種;酵母目未分類種unclassified_Saccharomycetales(OTU20)與嗜糖假絲酵母(Candidaglucosophila)、漢遜德巴利酵母(Debaryomyceshansenii)聚在一起,但序列一致度不高(分別為93%、83%),故不能確定該酵母的種屬。

a-細菌種水平系統發生樹及物種測序豐度圖;b-細菌未分類OTU的進化分析;c-真菌種水平系統發生樹及物種測序豐度圖;d-真菌未分類OTU的進化分析圖4 宣恩火腿主要微生物的進化分析Fig.4 Phylogenetic analysis of major microorganisms of Xuanen ham注:a~d中,樹枝長度為進化距離,分支節點處顯示1 000次重復檢驗的置信值(%);a和c中,進化樹樹枝顏色代表不同門,右邊柱狀圖展示的是物種在不同樣本中的測序數(reads),A、B、C依次為發酵前、中、后期的火腿樣本
2.6 功能預測分析
通過PICRUSt對16S rRNA測序結果進行細菌群落功能的預測,獲得OTU對應的COG家族相對豐度,結果見圖5。忽略未知功能后,宣恩火腿表面細菌功能主要包括氨基酸轉運及代謝(相對豐度11.2%~11.6%)、碳水化合物運輸及代謝(7.3%~8.0%)、能量產生與轉換(6.4%~6.6%)、無機離子轉運及代謝(7.1%~7.5%)、脂類運輸及代謝(3.4%~3.5%)等方面,這表明細菌群落對火腿蛋白質、脂肪的降解有重要作用。
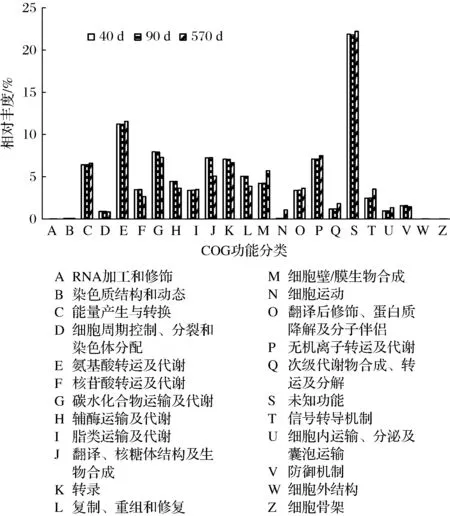
圖5 PICRUSt推斷的細菌COG功能相對豐度Fig.5 Relative abundance of inferred bacterial COG functions by PICRUSt
通過PICRUSt對ITS測序結果進行真菌群落的功能預測,獲得真菌MetaCyc pathway豐度(表2)。根據功能預測的豐度值,真菌群落主要功能包括有氧呼吸、乙醛酸循環、脂肪酸β氧化、GDP甘露糖生物合成、三羧酸循環、鳥氨酸降解Ⅱ等途徑。
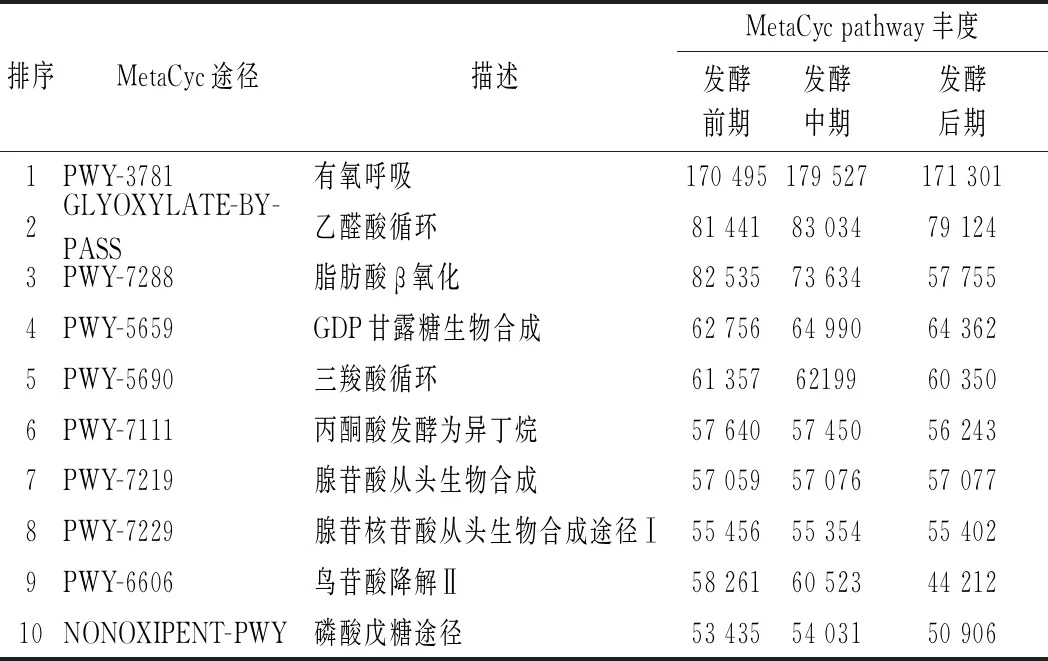
表2 PICRUSt 推斷的真菌MetaCyc pathway豐度(前10)Table 2 Abundance of inferred MetaCyc pathways by PICRUSt
其中,乙醛酸循環和脂肪酸β氧化都與脂肪降解密切相關,脂肪酸β氧化分解為乙酰CoA之后,在乙醛酸體內生成琥珀酸、乙醛酸和蘋果酸,從而進一步代謝;鳥氨酸降解Ⅱ與蛋白質降解途徑相關。因此,宣恩火腿表面真菌群落對脂肪、蛋白質的降解有重要作用。
3 結論與討論
發酵食品雖然種類繁多,但一般都具有各自相對穩定的微生物生態系統[23]。本文通過高通量測序技術研究了宣恩火腿發酵過程中表面細菌和真菌的多樣性,揭示了群落演替規律。
實驗中發現,宣恩火腿表面細菌主要為葡萄球菌屬(Staphylococcus)、沙雷氏菌屬(Serratia)、甲基菌屬(Methylobacterium)(包括根瘤菌科未知屬g_norank_f_Rhizobiaceae,其經進化分析也鑒定為甲基菌屬)等;其中,木糖葡萄球菌(S.xylosus)在發酵前、中期占絕對優勢(>98%),發酵后期下降到24.9%。有研究表明,金華火腿發酵中葡萄球菌是優勢細菌[24-25];云腿主要優勢細菌有表皮葡萄球菌(S.epidermidis)、肉葡萄球菌(S.carnosus)、木糖葡萄球菌(S.xylosus)等[26];木糖葡萄球菌能轉換氨基酸,使得亮氨酸和其他游離氨基酸含量增加從而提升火腿風味[27],發酵后期細菌群落多樣性的增加可能有利于火腿形成更加豐富的風味。
本實驗發現宣恩火腿表面真菌主要為曲霉屬(Aspergillus)、節菌屬(Wallemia)、未分類真菌unclassified_Fungi(OTU7)和酵母;曲霉屬在整個發酵過程中均有較高豐度,節菌屬在發酵中豐度逐漸降低,而酵母僅在發酵后期占據優勢。有研究表明,霉菌和酵母是眾多火腿的優勢真菌。宣威火腿表面和內部主要真菌為曲霉屬、酵母和節菌屬[7];云腿中的優勢真菌是青霉屬(Penicillium)、曲霉屬(Aspergillus)和酵母[26];金華火腿發酵前期青霉屬占優勢,后期曲霉屬占優勢,酵母菌也發揮了很重要的作用[24];Istrian火腿中分離的真菌主要來自散囊菌屬(Eurotium)、曲霉屬(Aspergillus)和青霉屬(Penicillium)[28]。
以上結果顯示,不同產地火腿的微生物群落組成雖然存在差異,但在屬水平上具有很高的一致性。這些微生物會分泌蛋白酶、脂肪酶等,促進蛋白質、脂肪水解產生游離氨基酸、脂肪酸,同時代謝產生各種特殊風味成分和保護火腿顏色等[29]。本文通過功能預測也發現,細菌和真菌均對宣恩火腿中蛋白質、脂肪的降解有重要作用。
通過高通量測序研究微生物多樣性時,常常會出現因物種數據庫序列信息不足而導致物種不能準確注釋的情況。本研究中,未分類真菌unclassified_Fungi(OTU7)無法準確注釋到門,進一步的進化分析顯示其與未培養真菌ITS序列的一致度為99%。隨著時間的推移,物種注釋的相關數據庫會不斷更新,這種問題應該會逐漸解決。后續試驗中也可嘗試利用各種培養基對unclassified_Fungi(OTU7)進行分離培養和進一步鑒定。
本文對宣恩火腿發酵過程中表面細菌和真菌多樣性進行了研究,對細菌、真菌優勢種之間的相關性進行了分析,有利于了解主要微生物種類及共存關系,可為后續研發宣恩火腿發酵劑和開展人工接種生產火腿作參考,以便進一步改善火腿品質、縮短加工周期和提升安全性。本文沒有研究火腿內部微生物,雖然內部微生物會受到表面微生物的抑制,但其也發揮了重要作用[24],后續可以開展相關工作。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26