基于形態(tài)特征和SSR標(biāo)記的斑茅繁殖策略的研究
2021-05-07 07:37:04安金嬋白史且陳智華鄢家俊張建波李達(dá)旭游明鴻毛德才楊金平李英主邱文靜
草地學(xué)報(bào) 2021年4期
關(guān)鍵詞:系統(tǒng)
安金嬋, 常 丹, 白史且, 陳智華, 鄢家俊, 張建波, 李達(dá)旭,游明鴻,毛德才, 王 艷, 楊金平, 李英主, 邱文靜
(1.西南民族大學(xué), 四川 成都 610041;2.四川省草原科學(xué)研究院, 四川 成都 611731)
繁育系統(tǒng)是指所有直接影響其后代遺傳組成的有性特征,包括花的綜合特性、花部各器官的壽命、傳粉方式、自交親和程度和交配系統(tǒng)等[1]。其中交配系統(tǒng)至關(guān)重要,它控制基因通過配子體傳遞了兩代間遺傳信息,決定了后代的基因型分布,對物種的遺傳變異水平和自然種群的遺傳組成產(chǎn)生深遠(yuǎn)的影響[2]。因此了解植物繁殖特性對于確定遺傳育種和植物保護(hù)制定至關(guān)重要[3-4]。
人們研究植物繁殖性狀主要是從物候?qū)W、形態(tài)特征和人工授粉試驗(yàn)中推斷出來的[5-6]。如觀測其物候期、傳粉特性、雜交指數(shù)以及套袋試驗(yàn)等指標(biāo)來推測植物的交配系統(tǒng)[7-9]。但是,這些指標(biāo)的測量只能推測異交和自交的相對程度,不能反映植物自然群體的真實(shí)異交率。隨著科技的發(fā)展,分子標(biāo)記的使用克服了這些困難,分子標(biāo)記不受外界環(huán)境和植物生長發(fā)育階段的影響[10],被廣泛用于研究和確定植物交配系統(tǒng)。簡單重復(fù)序列(simple sequence repeats,SSR)標(biāo)記因其重復(fù)性好、操作簡便、高效和多態(tài)性,已成為一種應(yīng)用廣泛的分子標(biāo)記工具[11]。根據(jù)遺傳標(biāo)記估計(jì)的交配系統(tǒng)參數(shù)有助于特定植物物種保護(hù)策略和育種選擇方式的發(fā)展。
斑茅(Saccharumarundinaceum)是甘蔗屬(Saccharum)密叢高大草本植物,適應(yīng)性強(qiáng),廣泛種植在我國南方荒山荒坡、鹽堿地、沿海灘涂等地。其種內(nèi)變異豐富、抗旱抗蟲耐貧瘠性強(qiáng)、分蘗多、生長快,有較強(qiáng)的生態(tài)競爭能力[12-13],在緩解能源供給緊張的同時(shí),對改善我國生態(tài)環(huán)境具有重要意義。其優(yōu)異性狀特點(diǎn)也倍受甘蔗(Saccharumofficinarum)育種者的普遍關(guān)注,對高產(chǎn)高糖甘蔗品種的培育有重要意義。但是,由于斑茅花期普遍比甘蔗早3~6個(gè)月[14],使得斑茅優(yōu)良特性的利用率低下,且斑茅-甘蔗雜交困難。目前國內(nèi)外對斑茅植物的研究主要集中在種質(zhì)資源的調(diào)查及評(píng)價(jià)、形態(tài)學(xué)、細(xì)胞學(xué)、蛋白質(zhì)標(biāo)記、遺傳多樣性及種質(zhì)資源抗逆性的研究等方面[15],對斑茅植物生殖生物學(xué)方面試驗(yàn)性研究還未見報(bào)道。而植物學(xué)性狀與分子標(biāo)記兩者相結(jié)合才能選出最佳育種材料,達(dá)到預(yù)期的育種目標(biāo)[16]。故本研究以收集自我國7省的45份野生斑茅為研究對象,對斑茅表型特征和交配系統(tǒng)進(jìn)行研究,以期闡明斑茅的繁殖特性及其有性繁殖力的情況,為斑茅資源的開發(fā)利用、保護(hù)、雜交育種及繁殖技術(shù)提供基礎(chǔ)依據(jù)。
1 材料與方法
1.1 材料
試驗(yàn)材料來自四川省成都市大邑縣韓場鎮(zhèn)種質(zhì)資源圃(30°25′ N,103°45′ E),該資源圃收集了來自我國陜西、四川、貴州、廣西、云南、廣東、海南7省的野生無性系斑茅種質(zhì)資源(四川省草原科學(xué)研究院提供),于2012年移栽,植株高大健壯,平均株高 4.5 m。共有45個(gè)居群(如表1),每個(gè)居群種于一個(gè)長8 m寬5 m的地塊中,每個(gè)地塊包含12個(gè)無性系單株。
1.2 方法
1.2.1花粉量與花粉-胚珠比P/O 采用血球計(jì)數(shù)法[17]統(tǒng)計(jì)斑茅花粉量。從每個(gè)斑茅居群中隨機(jī)選取3個(gè)單株進(jìn)行標(biāo)記,每株選擇3個(gè)即將開放的小花,放入1.5 mL離心管中,帶回實(shí)驗(yàn)室。將離心管中的花藥搗碎,加入0.1 mL 1.0 mol·L-1的鹽酸對花藥進(jìn)行水解一個(gè)小時(shí),用移液槍吸取約0.01 mL 進(jìn)行制片,在顯微鏡(10×)(OLYMPUS-BX53F,日本)下直接觀察記錄5個(gè)大方格的數(shù)值(血球計(jì)數(shù)板計(jì)數(shù)池長寬各3.0 mm,分為9個(gè)大方格,每個(gè)大方格容積為0.1 mm3),記錄數(shù)值A(chǔ),1朵花的花粉總數(shù)=0.1×A/(0.5×10-3×3)=200A/3。檢測一朵小花的胚珠數(shù),P/O=1朵小花花粉量/一朵小花的胚珠數(shù)。根據(jù)Cruden[18]標(biāo)準(zhǔn)(P/O為2.7~5.4時(shí),其繁育系統(tǒng)為閉花受精;P/O為18.1~39.0時(shí),繁育系統(tǒng)為專性自交;P/O為31.9~396.0時(shí),繁育系統(tǒng)為兼性自交;P/O為244.7~2 588.0時(shí),繁育系統(tǒng)為兼性異交;P/O為2 108.0~195 525.0時(shí),繁育系統(tǒng)為專性異交)來判斷斑茅的交配系統(tǒng)。
1.2.2花粉活力與花粉壽命 采用TTC法[19]測定花粉活性:選擇即將開放的斑茅小花采集花藥,將花藥撒于載玻片上,滴加0.1%的TTC染液進(jìn)行染色,后于顯微鏡下統(tǒng)計(jì)被染色和未被染色的花粉數(shù)目。計(jì)算花粉活力百分比。
隨機(jī)選取17個(gè)居群,每個(gè)居群選取3個(gè)單株抽穗2/3但未盛開的斑茅花穗無性系材料,剪取單株穗莖(帶葉片)帶回室內(nèi)進(jìn)行自來水養(yǎng)莖,從正在開放的花上收集一定量的花粉,每隔0.5 h用TTC染色,染成紅色的記為有活力的花粉,其余均記為無活力的花粉(由同一個(gè)人計(jì)數(shù)),并計(jì)算花粉活力百分比,求出花粉最長存活時(shí)間,得出花粉壽命。
1.2.3雜交指數(shù)OCI(outcrossingindex)測定 每個(gè)居群選擇1個(gè)單株上的一條斑茅花穗進(jìn)行花序直徑、花朵大小以及對雌雄蕊開花時(shí)期形態(tài)的觀測,按照Dafni[20]的標(biāo)準(zhǔn)進(jìn)行OCI值的評(píng)估,并進(jìn)行繁育系統(tǒng)評(píng)判。
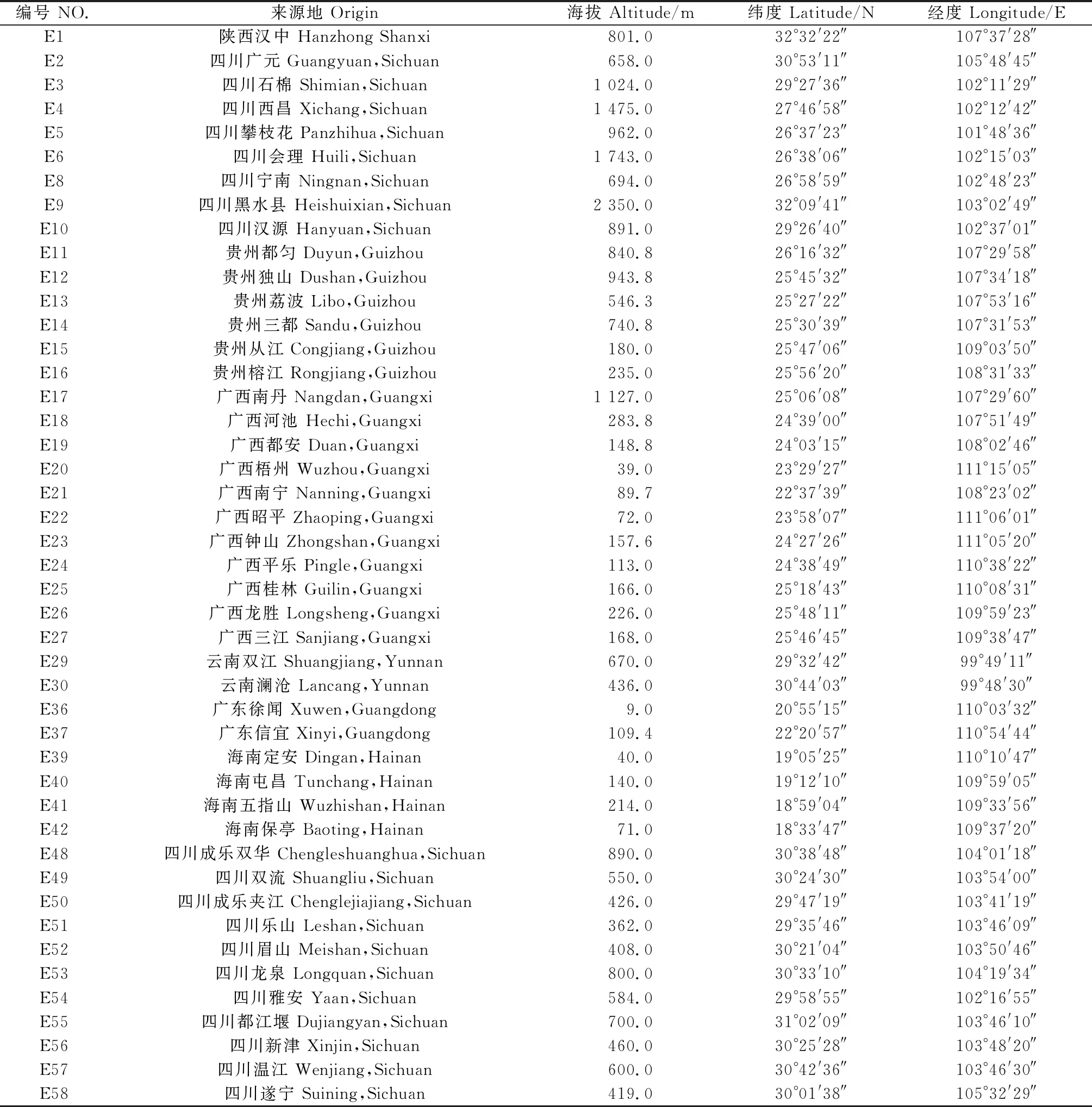
表1 斑茅材料采集地及其海拔、經(jīng)緯度
①采用游標(biāo)卡尺測量花序直徑,花序直徑<1 mm記為0;1~2 mm記為1;2~6 mm記為2;>6 mm記為3。
②花藥開裂時(shí)間與柱頭可授期之間的時(shí)間間隔,采用聯(lián)苯胺-過氧化氫法[18](V1%聯(lián)苯胺∶V3%過氧化氫∶V水=4∶11∶22)對開花前的柱頭進(jìn)行可授性的鑒定,同時(shí)于顯微鏡下觀測柱頭周圍是否有花粉粒,雌蕊先熟記為0;雄蕊先熟記為1。
③柱頭與花藥的空間位置,同一高度記為0;空間分離記為1。
三者之和為OCI值。之后依據(jù)OCI值進(jìn)行繁育系統(tǒng)的判定。其標(biāo)準(zhǔn)為:OCI=0,閉花受精;OCI=1時(shí),專性自交;OCI=2時(shí),兼性自交;OCI=3時(shí),自交親和,有時(shí)需要傳粉者;OCI=4時(shí),部分自交親和、異交、需要傳粉者;OCI=5時(shí)為專性異交。
1.2.4柱頭可授性檢測 用聯(lián)苯胺-過氧化氫法[21](V1%聯(lián)苯胺∶V3%過氧化氫∶V水=4∶11∶22)對斑茅開花前一天、開花時(shí)刻、開花后幾天進(jìn)行柱頭可授性測定。取斑茅柱頭浸入含有聯(lián)苯胺-過氧化氫溶液的凹面載玻片上,其周圍溶液出現(xiàn)藍(lán)色及有氣泡產(chǎn)生,則證明柱頭具有可授性。
1.2.5SSR分子標(biāo)記研究交配系統(tǒng) 在45個(gè)居群中,對有30個(gè)子代以上的母株及其子代進(jìn)行SSR比較分析。母株取野生無性系斑茅居群中的單株進(jìn)行樣本測定,子代由母株收集而來的種子經(jīng)發(fā)芽育苗后進(jìn)行測定。共采集15個(gè)居群39個(gè)半同胞家系材料(39個(gè)單株在開放授粉狀態(tài)下每個(gè)單株上收獲的種子形成的個(gè)體集合),每個(gè)家系30個(gè)子代,除去育苗及試驗(yàn)過程中的損耗,共獲得1 158個(gè)子代苗。該15個(gè)居群均為6倍體無性系材料。
基因組DNA的提取:DNA采用Plant Gneomic DNA Kit方法提取。采用2%的瓊脂糖凝膠測定其質(zhì)量,紫外分光光度計(jì)測定濃度,用無菌水稀釋濃度為20 ng·μL-1,存放至-80℃冰箱備用。
引物的篩選:參照Zhang等[22]發(fā)表的SSR引物,選取5個(gè)樣本對引物進(jìn)行篩選,篩選出如表2所示11對擴(kuò)增條帶清晰、多態(tài)性好、穩(wěn)定性高的引物進(jìn)行試驗(yàn)。

表2 用于斑茅DNA擴(kuò)增的11對SSR引物及其退火溫度
PCR擴(kuò)增:PCR擴(kuò)增采用20 μL體系,包括0.5 μL上游引物,0.5 μL下游引物,10 μL 2×Reaction Mix,1.5 μL 0.1 μg·μL-1模板DNA,最后用ddH2O補(bǔ)足。
擴(kuò)增反應(yīng)在PCR東勝(ETC811)擴(kuò)增儀上進(jìn)行,擴(kuò)增程序?yàn)?5℃預(yù)變形5 min,94℃變性30 S,退火45 S,72℃延伸45 S,33個(gè)循環(huán);72℃延伸5 min,4℃保存。擴(kuò)增產(chǎn)物以DL2000 Marker對照,用1.5%的瓊脂糖凝膠電泳分離,150 V電壓下電泳2 h,最后拍照。
SSR數(shù)據(jù)分析:本研究采用多位點(diǎn)混合交配系統(tǒng)模型估計(jì)多位點(diǎn)異交率和單位點(diǎn)異交率[23]。多位點(diǎn)異交率又稱為異交率的多位點(diǎn)估計(jì),在估計(jì)時(shí)它考慮到了確定性的異交,也考慮到了檢測不到但確定已出現(xiàn)的異交。使用Excel對PCR擴(kuò)增數(shù)據(jù)進(jìn)行整理記錄其基因型。MLTR3.4[24]統(tǒng)計(jì)軟件對交配系統(tǒng)進(jìn)行各參數(shù)的計(jì)算。在親本基因型已知的情況下,采用最大期望值法,在95%的置信區(qū)間,選擇家系內(nèi)1 000次重復(fù)取樣,估算多位點(diǎn)異交率(tm),單位點(diǎn)異交率(ts),雙親近交系數(shù)(tm—ts)和親本近交系數(shù)(F)。
2 結(jié)果與分析
2.1 花粉量與胚珠比
隨機(jī)對20個(gè)斑茅居群的花粉量進(jìn)行了測定,發(fā)現(xiàn)斑茅1朵小花中的3個(gè)花藥的總花粉量平均為5 897,1朵小花的胚珠數(shù)為1。因此,斑茅的花粉量與胚珠比為5 897,根據(jù)Cruden[15]的標(biāo)準(zhǔn)判斷,斑茅屬于專性異交。
2.2 花粉活力與花粉壽命
如表3所示,種群上斑茅花粉活力從開花時(shí)刻到開花后3小時(shí),花粉活力逐漸減弱。從開花時(shí)刻的44.49%到開花后1小時(shí)35.61%再到開花后3小時(shí)的6.72%,結(jié)果表明斑茅花粉活力較低,且花粉壽命時(shí)間約為3個(gè)小時(shí),大大限制了花粉的傳粉效率。
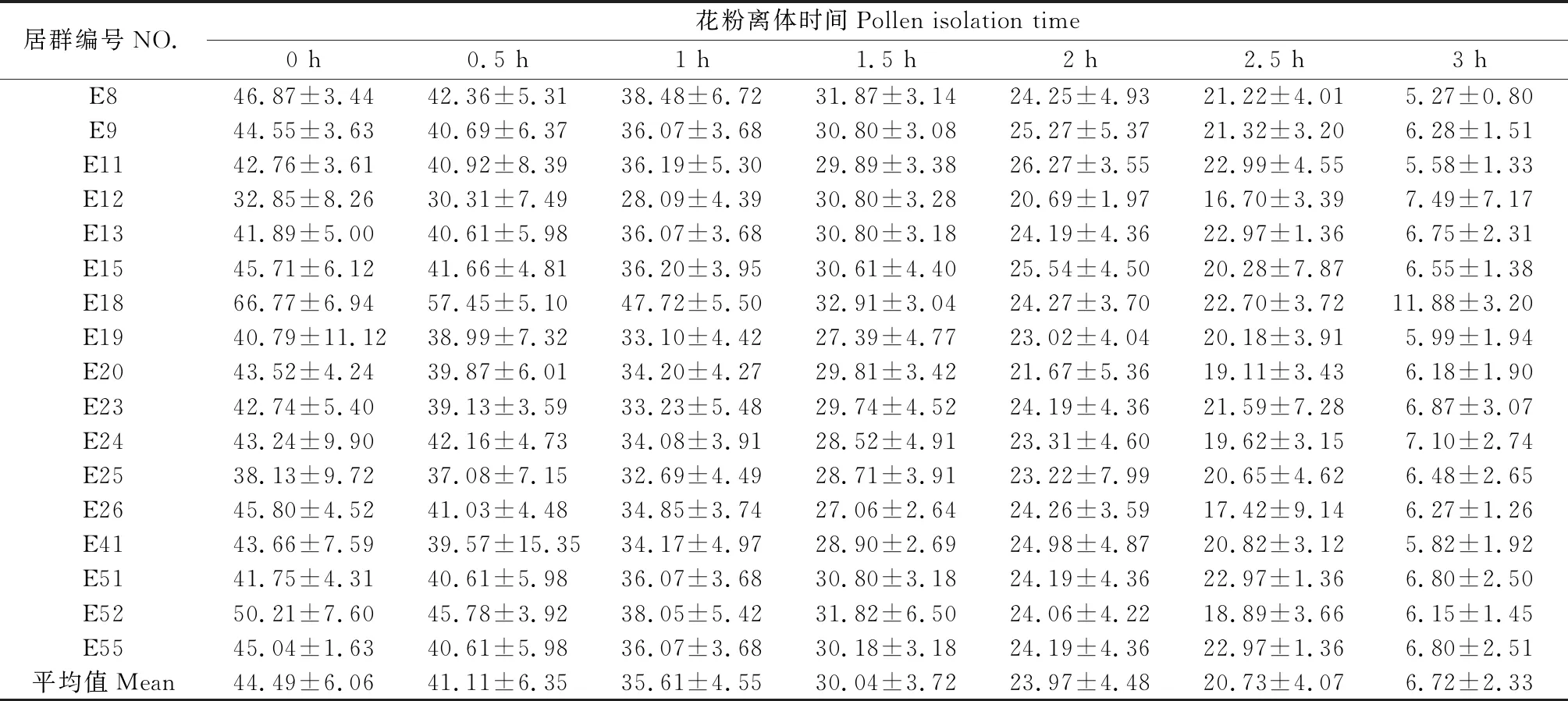
表3 花藥離體后花粉活性率
2.3 柱頭可授性檢測
用聯(lián)苯胺-過氧化氫法對斑茅開花前一天,開花時(shí)刻、開花后幾天進(jìn)行柱頭可授性測定。結(jié)果如圖1所示,斑茅在開花前一天就具有可授性,到開花時(shí)刻柱頭可授性最強(qiáng),之后幾天柱頭可授性持續(xù)下降,到開花后第5天,斑茅柱頭可授性趨近于無。表明斑茅柱頭可授期不是限制斑茅有性生殖的影響因素。

圖1 不同時(shí)期斑茅柱頭可授性
2.4 雜交指數(shù)
如表4所示,斑茅開花后平均花序直徑為1.18±0.08 mm。在開花前一天柱頭就具有可授性,開花前一天在柱頭周圍觀察到了少量的花粉粒,證明在開花前花藥就有輕微的開裂,故雌雄蕊同時(shí)成熟。柱頭與花藥空間位置分離,所以雜交指數(shù)(OCI)為2,根據(jù)Dafni[17]的標(biāo)準(zhǔn)判斷斑茅繁育系統(tǒng)屬于兼性自交。

表4 斑茅雜交指數(shù)OCI
2.5 SSR分子標(biāo)記研究交配系統(tǒng)
PCR擴(kuò)增產(chǎn)物采用1.5%的瓊脂糖凝膠電泳分離,部分電泳圖如圖2所示。對電泳條帶進(jìn)行數(shù)據(jù)讀取,利用MLTR3.4軟件對斑茅無性系隨機(jī)抽取的15個(gè)居群中的39個(gè)半同胞家系母本單株和1 158個(gè)子代進(jìn)行交配系統(tǒng)評(píng)估(表5),結(jié)果如下:11個(gè)位點(diǎn)估算出多位點(diǎn)異交率(tm)為0.864,單位點(diǎn)異交率(ts)為0.853,自交率(s)為0.136,表明斑茅種群具有較高的異交率水平,同時(shí)存在一部分的自交;多位點(diǎn)異交率相關(guān)系數(shù)(rt)為0.455,多位點(diǎn)父本相關(guān)性(rP(m))為0.132,有效花粉供體數(shù)目為7.6,也進(jìn)一步表明斑茅種群具有一定比例的自交;親本近交系數(shù)(F)為0.318,大于0,表明有過剩的純合子,近交優(yōu)勢較大;近交衰退系數(shù)(δ)為-4.144,表明斑茅沒有明顯的近交衰退現(xiàn)象。
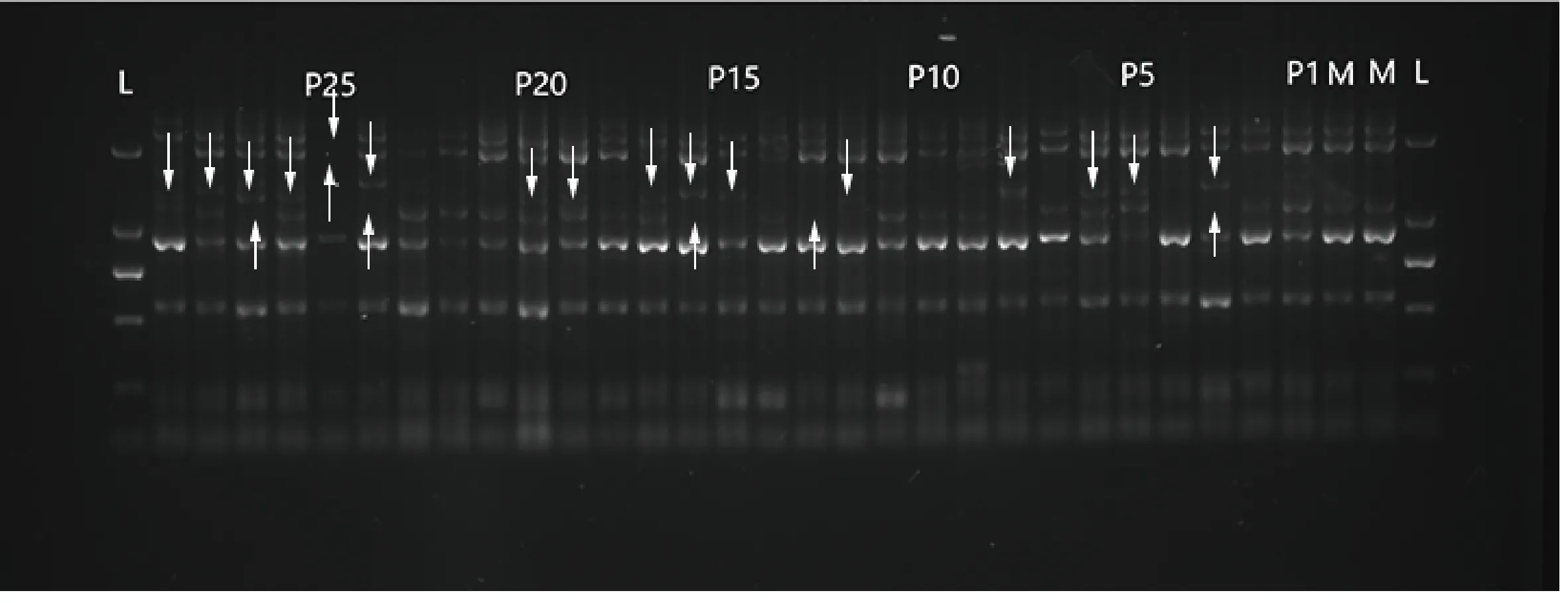
圖2 引物L(fēng)27在部分母本與其子代中的PCR擴(kuò)增結(jié)果

表5 斑茅種群水平上的交配系統(tǒng)參數(shù)評(píng)估
利用MLTR3.4對斑茅家系進(jìn)行交配系統(tǒng)分析發(fā)現(xiàn)(表6),與在斑茅種群水平上的結(jié)果不同,雖然都具有較高的異交率,但居群家系中斑茅多位點(diǎn)異交率變化差異較大(tm=0.601~1.200),單位點(diǎn)異交率同樣表現(xiàn)出較大差異(ts=0.575~1.200)。1,13,25,27,48,54,57號(hào)家系雙親近交系數(shù)<0,說明居群家系間無近親交配現(xiàn)象。其余居群家系間有較輕程度的近交現(xiàn)象,其中E26號(hào)居群雙親近交系數(shù)最大。
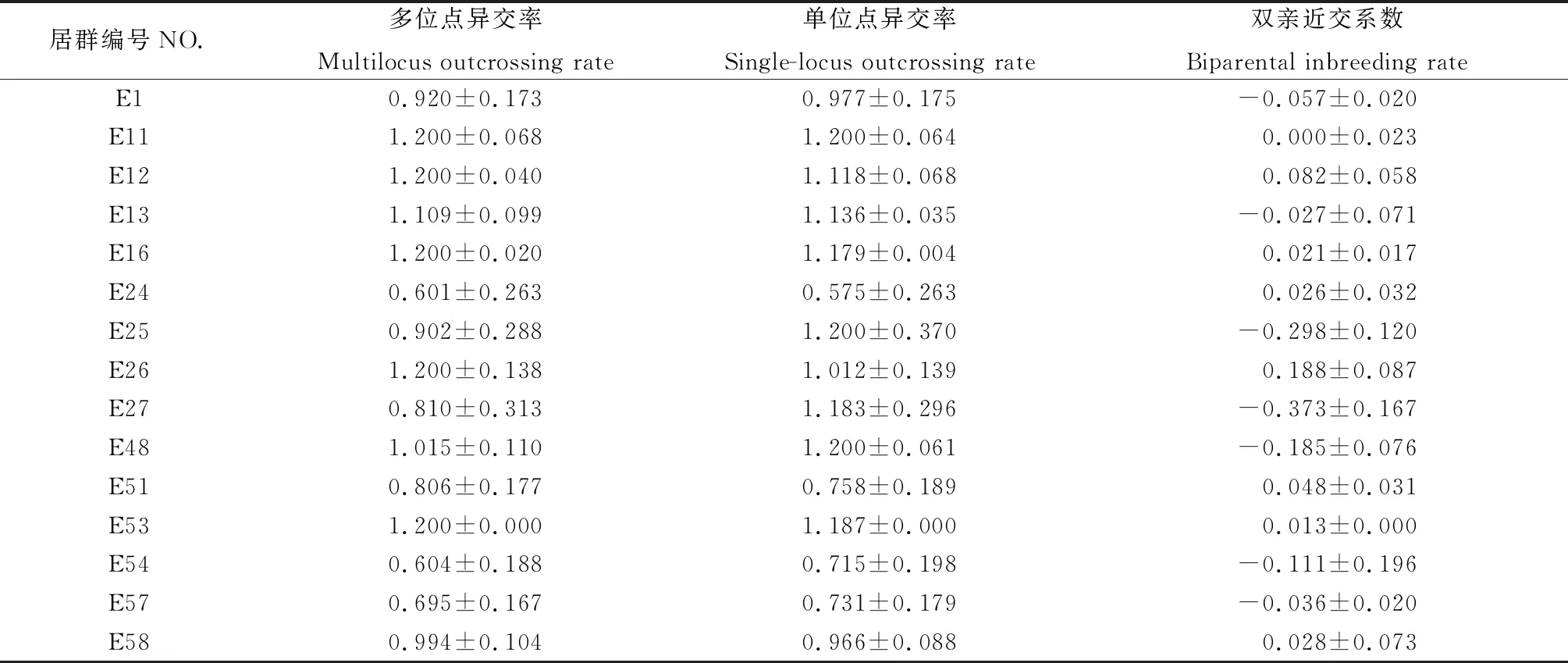
表6 斑茅15個(gè)居群交配系統(tǒng)參數(shù)
3 討論和結(jié)論
雜交指數(shù)、花粉量與胚珠比在研究植物繁育系統(tǒng)中廣泛使用。本研究中,P/O為5 897,根據(jù)Cruden的標(biāo)準(zhǔn)[24]進(jìn)行歸類分析,斑茅屬于專性異交。斑茅的大花粉量符合其多年生禾本科風(fēng)媒植物的特性[25],其結(jié)果也與楊等[14]統(tǒng)計(jì)斑茅花粉量大致相同。OCI指數(shù)為2,根據(jù)Dafni的標(biāo)準(zhǔn)[19],斑茅繁育系統(tǒng)為以自交為主的混合交配系統(tǒng),與本研究P/O結(jié)果具有一定的重合性。
斑茅花粉活力低,平均花粉活性百分率僅為44.49%,該結(jié)果與雷敬超[26]連續(xù)7年對同一屬的甘蔗花粉活力測定的結(jié)果相似。花粉離體后活力迅速下降,壽命約為3個(gè)小時(shí),大大限制了花粉的傳粉時(shí)間、距離和傳粉效率,所以花粉的初始活力可能是影響斑茅自交結(jié)實(shí)率低的一個(gè)因素。花的壽命是植物繁殖的一個(gè)關(guān)鍵特征,它平衡了花粉的傳播速度與花的維護(hù)成本。柱頭可授性的時(shí)間長短對于植物花期來說是極為重要的,它對植物的授粉成功率和結(jié)果有一定的影響[27-28]。柱頭可授性結(jié)果顯示斑茅柱頭在開花前就具有柱頭可授性,且柱頭可授性時(shí)間長,比同一屬的甘蔗柱頭可授性時(shí)間略短[29]。這一結(jié)果與趙元杰等[22]對芒的花粉壽命與柱頭可授性結(jié)果相一致。它彌補(bǔ)了花粉離體后活力迅速下降這一劣勢,對異花傳粉植物的傳粉成功率有一定的影響[30-31],不是影響斑茅有性生殖的限制因素。
本研究對斑茅居群的異交率進(jìn)行了評(píng)估。統(tǒng)計(jì)交配系統(tǒng)參數(shù)時(shí),Ritland和Jain[24]認(rèn)為多數(shù)情況下采用3~6個(gè)共顯性位點(diǎn)即可得到準(zhǔn)確的結(jié)果,我們比較了采用6個(gè)位點(diǎn)和11個(gè)位點(diǎn)統(tǒng)計(jì)斑茅異交率,結(jié)果發(fā)現(xiàn)兩種情況下異交率相差不大。這一結(jié)果與孫小涵等[32]采用3個(gè)位點(diǎn)與9個(gè)位點(diǎn)統(tǒng)計(jì)荊條的異交率結(jié)果相似。雙親近交系數(shù)(tm—ts)反映了雙親自交的程度,斑茅異交率處于較高水平(tm=0.853),tm—ts>0,表明斑茅居群存在自交。這一結(jié)果可能與斑茅的居群密度、風(fēng)力和花期不完全同步,先開花的植株未能更多地接受異源的花粉有一定聯(lián)系[33-34]。斑茅多位點(diǎn)異交率相關(guān)系數(shù)較大(rt=0.445),親本近交系數(shù)F>0,說明在該斑茅居群中異交占較大優(yōu)勢,且和隨機(jī)交配相比,近交會(huì)產(chǎn)生較多的純合子[35]。Tm>(1—F)/(1+F)表明斑茅種群偏離哈迪-溫伯格平衡,導(dǎo)致其偏離的原因可能與近交、氣候、單株密度、營養(yǎng)繁殖等有關(guān)[36]。不同斑茅家系也都具有較高的異交率,但異交率水平存在較大差異(tm=0.601~1.200),這可能是親本的自交可育程度,花粉的活性,資源圃中的位置以及其材料來源生境所致。
綜合花粉與胚珠比P/O、雜交指數(shù)、斑茅傳粉特征及交配系統(tǒng)結(jié)果,本研究初步認(rèn)為斑茅的繁殖特性為異交為主,自交為輔的兼性異交模式。
猜你喜歡
工業(yè)設(shè)計(jì)(2022年8期)2022-09-09 07:43:20
軍民兩用技術(shù)與產(chǎn)品(2021年10期)2021-03-16 06:05:30
北京測繪(2020年12期)2020-12-29 01:33:58
裝備制造技術(shù)(2019年12期)2019-12-25 03:06:46
制造技術(shù)與機(jī)床(2019年10期)2019-10-26 02:47:06
中國洗滌用品工業(yè)(2019年4期)2019-05-11 09:27:34
鐵道通信信號(hào)(2018年5期)2018-06-28 03:06:24
家庭影院技術(shù)(2017年9期)2017-09-26 03:41:45
知識(shí)經(jīng)濟(jì)·中國直銷(2017年5期)2017-06-15 20:28:19
通信電源技術(shù)(2016年6期)2016-04-20 06:21:32
