赤狐ESR1基因啟動子的克隆及生物信息學分析
2021-05-10 03:15:24周瑞紅彭永東李祥龍
河北科技師范學院學報 2021年1期
周瑞紅,彭永東,李祥龍
(河北科技師范學院動物科技學院 河北省特色動物種質資源挖掘與創新重點實驗室,河北 秦皇島,066600)
赤狐毛皮經濟價值較高,能給養狐業帶來巨大經濟效益。然而,作為被保護的動物之一,赤狐數量在不斷減少,其產仔數少是造成這種現象的主要原因之一,且狐貍的產仔數遺傳力低,采用常規育種方法很難在短時間內取得較大進展。另外,赤狐是季節性發情動物,在每年的12月至2月發情。雌激素受體1(estrogen receptor 1,ESR1)是哺乳動物雌激素受體的兩種亞型之一[1,2]。ESR1基因是一種核酸受體,具有轉錄調控蛋白質的功能,參與雌性動物性腺基因的表達與調控,且能影響胚胎發育和系統進化,最終能夠影響動物的繁殖表現[3,4]。目前已有研究發現,ESR1基因不僅可以調節性腺生長和分化,且能在機體能量代謝方面發揮重要作用[5,6],可以影響動物的繁殖性能。自雌激素受體的氨基酸序列公布后,對ESR1基因的研究也取得了較大進展[7]。王宵燕等[8]利用ESR1基因上的PvuⅡ酶切位點作為蘇姜豬繁殖相關性狀遺傳標記可以提高產仔數。有試驗結果表明,綿羊ESR1基因主要位于細胞核中,并且參與機體的轉錄調控機制[9],也有試驗以大白母豬進行分析,表明B基因較A基因而言是優勢基因,可以應用于優化繁殖性能[10]。綜合分析表明,ESR1基因對動物育種具有一定應用前景。因此,對ESR1基因啟動子區進行研究以便為今后的育種工作提供理論依據。
近年來,ESR1基因在家畜繁殖性能的研究和利用方面已經成為熱點,前人的研究為其生物信息學分析提供了有價值的理論依據和研究意義。但國內ESR1基因的研究集中在豬和綿羊,未見狐貍ESR1基因相關研究。為此,筆者利用PCR技術擴增ESR1基因啟動子區序列,并將其構建至雙熒光素酶報告基因載體,通過在線軟件對預測結果進行深入分析,以期擴充已有數據庫,為探究ESR1基因啟動子區的作用提供依據。
1 材料與方法
1.1 試驗材料與試劑
試驗所用赤狐來自河北省昌黎縣金島育種場,取肌肉組織-20 ℃保存,用于DNA提取。
試驗用到的pMDTM18-T Vector Cloning Kit,T4-DNA Ligase,限制性內切酶MluⅠ和XhoⅠ以及DNA Marker購自Takara寶生物工程有限公司;DH5α感受態細胞購于博邁德生物技術有限公司;2×Es Taq MasterMix(Dye)購于康為世紀生物有限公司;DNA膠回收試劑盒購自OMEGA公司(美國);質粒DNA小量提取試劑盒購自天根生化科技有限公司。引物合成和測序由北京六合華大基因公司完成。
1.2 試驗方法
1.2.1引物設計與合成由于基因的啟動子區域通常在轉錄起始位點上游1.0~2.0 kb,故本次研究選取轉錄起始位點上游約1.8 kb為候選啟動子區。參考NCBI(https://www.ncbi.nlm.nih.gov/)中GenBank數據庫公布的赤狐ESR1基因序列(登錄號:XM_025998015.1),以5’ UTR上游序列約1 800 bp及轉錄起始位點下游78 bp為模板,利用Primer Primer 5.0軟件設計引物,利用DNASTAR7.1軟件進行限制性內切酶位點分析,上游引物和下游引物分別引入MluⅠ和XhoⅠ限制性酶切位點及其對應的保護堿基。上游引物為:5’-CGACGCGTGCTGTTCATAGTCCAGGGTT-3’,下游引物為:5’- CCCTCGAGGCTCGCATGTGCGTGTAG-3’。引物由北京六合華大基因有限公司完成。
1.2.2赤狐ESR1基因轉錄調控區生物信息學分析利用在線預測軟件Neural Network Promoter Predition,Promoter 2.0,TSSW和TSSG預測啟動子區;通過在線預測軟件MethPrimer預測CpG島;通過在線預測軟件Nsite,AliBaba 2.1,Cister和JASPAR數據庫預測核心啟動子區的轉錄因子結合位點。在線預測軟件網址見表1。

表1 分子生物學及數據分析軟件
1.2.3赤狐ESR1基因啟動子區PCR擴增以赤狐基因組DNA為模板進行PCR擴增。PCR擴增體系為40 μL:20 μL PrimeSTAR Max DNA Premix(2×),14 μL ddH2O,2 μL DNA模板,上下游引物各2 μL(引物濃度10 μmol/L)。PCR反應條件:94 ℃預變性5 min;經94 ℃變性30 s,57 ℃退火30 s,72 ℃延伸2 min,共30個循環;72 ℃終延伸10 min,12 ℃保存。PCR產物用質量濃度為15 g/L的瓊脂糖凝膠電泳檢測。
1.2.4赤狐ESR1基因候選啟動子區雙熒光素酶報告基因載體的構建將上述PCR產物進行瓊脂糖凝膠電泳并使用回收試劑盒回收目的片段,回收得到的擴增片段連接至pMDTM18-T載體;提質粒后連同雙熒光素酶載體pGL3-Basic一起分別用限制性內切酶MluⅠ和XhoⅠ進行雙酶切(37 ℃, 2~3 h),雙酶切反應體系為20 μL:2 μL Green Buffer(10×), 2 μL 質粒,14 μL ddH2O,MluⅠ和XhoⅠ各1 μL 。利用T4-DNA Ligase將雙酶切后的目的片段與pGL3-Basic載體16 ℃過夜連接,連接體系為20 μL:2 μL T4 DNA Ligase,2 μL T4 DNA Ligase Buffer(10×), 2 μL 酶切后的pGL3-Basic質粒,8 μL目的片段,6 μL ddH2O。通過熱激法將連接產物轉化至大腸桿菌DH5α感受態細胞;后進行涂板培養,挑取陽性單克隆菌落進行鑒定、測序,測序成功后提質粒,-20 ℃保存備用。
3 結果與分析
3.1 赤狐ESR1基因啟動子區序列的擴增及載體構建
通過PCR擴增出赤狐ESR1基因啟動子區約1 880 bp的片段,PCR產物經質量濃度為15 g/L的瓊脂糖凝膠電泳檢測,其結果與預期片段大小一致(圖1),經pMDTM18-T中間載體后構建至表達載體pGL3-Basic上,經菌液PCR、雙酶切及測序鑒定后,目的片段大小一致且序列正確,表明重組克隆載體pGL3-Basic-ESR1構建成功(圖2),該重組質粒可用于后續試驗。
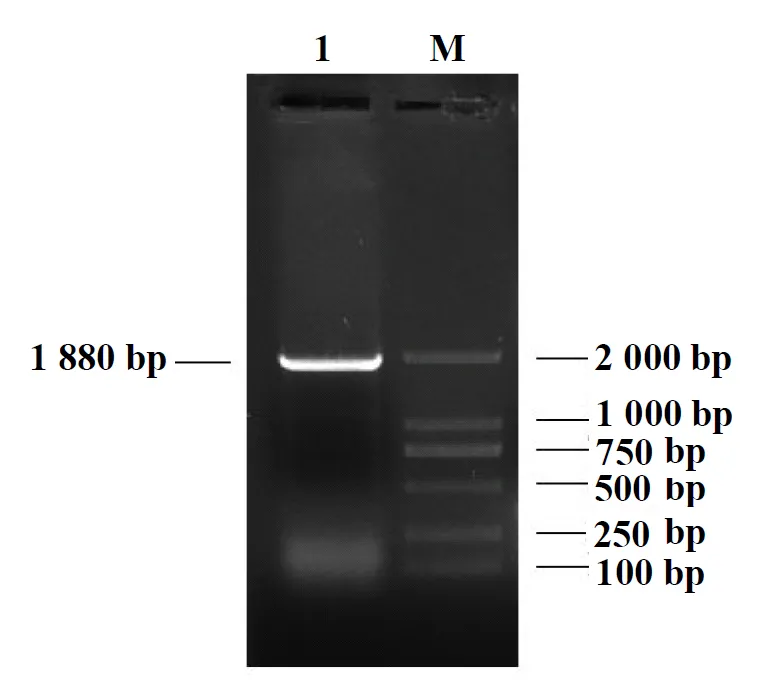
1:目的片段;M:D2000 DNA Marker圖1 PCR電泳檢測結果
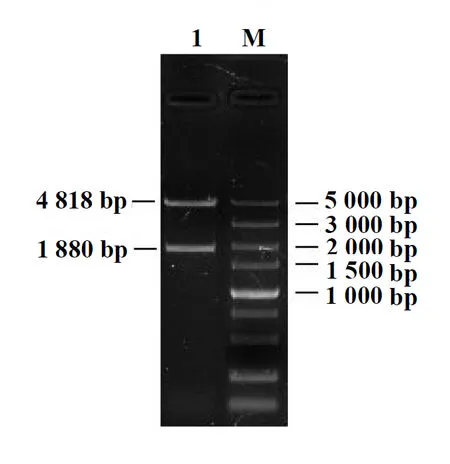
1:目的片段;M:D2000 Plus DNA Marker圖2 重組載體雙酶切檢測結果
3.2 赤狐ESR1基因啟動子區序列生物信息學分析
運用在線軟件對測序得到的結果進行序列分析,GC含量占49.89%,AT含量占50.11%。用在線軟件Promoter 2.0預測出2個啟動子臨界位置,分別為-1 602 bp(得分0.583)和-502 bp(得分0.671);TSSW程序預測啟動子位置在-1 702 bp(得分1.04);TSSG預測啟動子位置在-1 548 bp(得分6.81)。在線預測軟件Neural Network Promoter Predition分析發現2個核心啟動子(表2)。
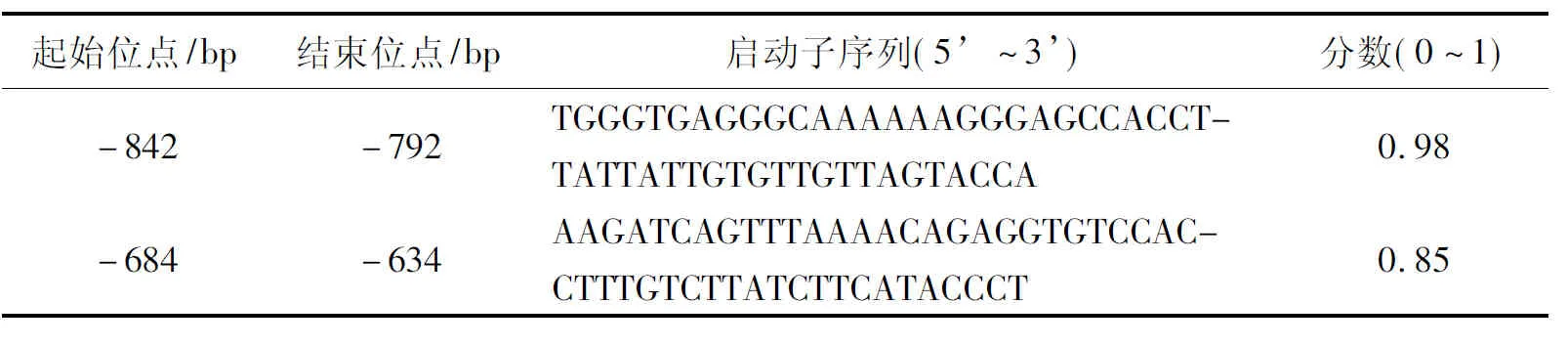
表2 赤狐ESR1基因啟動子區Neural Network Promoter Predition程序預測結果
3.3 赤狐ESR1基因CpG島及轉錄因子結合位點預測
所獲得的赤狐ESR1基因啟動子區存在多個轉錄因子作用元件,如AP-1,SRF,GATA-1,c-Jun,Oct-1,SP1,NF-1和cEBP等識別位點,其中Sp1家族結合位點、NF-1和cEBP的結合位點出現頻率較高且在多個預測軟件中出現。綜合多個軟件對轉錄因子結合位點預測分析,單個軟件預測結果不做考慮,結果見表3。
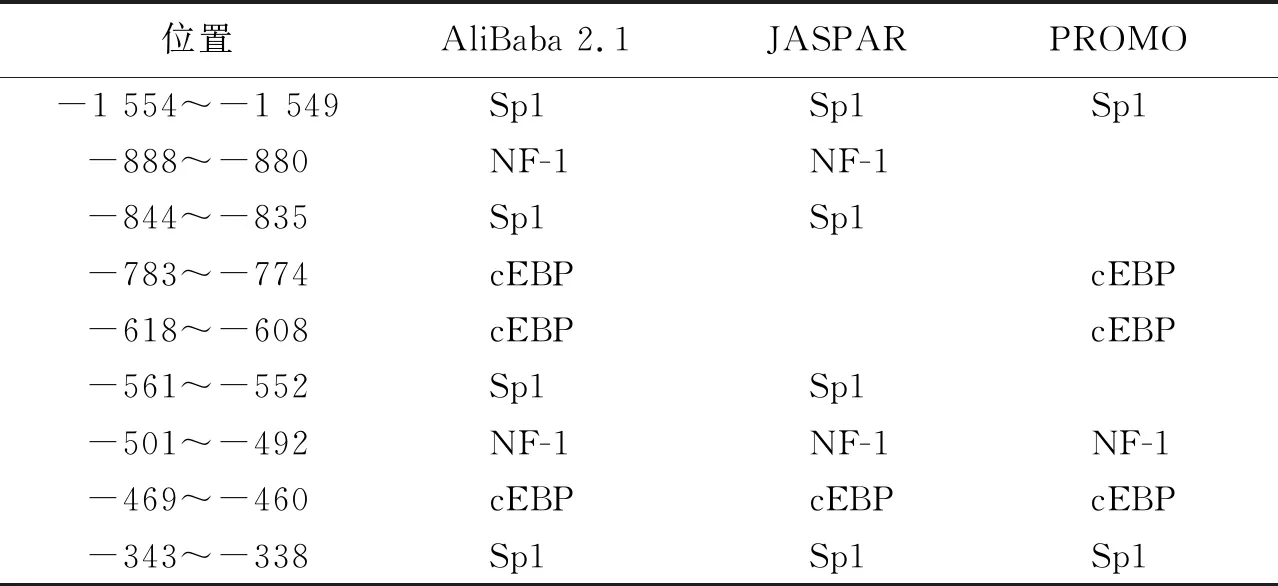
表3 赤狐ESR1基因啟動子區轉錄因子結合位點預測結果
一般認為,CpG島大于100 bp,且GC含量大于50%,使用MethPrimer軟件預測分析顯示ESR1基因啟動子區存在一個CpG島(圖3),長度為148 bp,位于-1 749~-1 602 bp。

圖3 赤狐ESR1基因CpG島預測結果
4 結論與討論
ESR1基因通過轉錄調控蛋白控制雌激素基因的轉錄過程進而對生殖發育等功能產生影響[11]。已經有研究證實ESR1基因在動物繁殖中發揮著重要的作用[12],因此在今后的育種工作可以利用相關的研究成果以提高產仔數,為養狐業提供經濟效益。基因表達調控中最重要的就是轉錄的起始,而轉錄調控的實質是通過上游調控序列與啟動子及轉錄因子相互作用來調控基因的表達[13],基因啟動子活性區域研究對揭示基因功能具有重要的意義[14]。然而,赤狐ESR1基因啟動子的序列結構、功能和調控機理等尚未報道。因此,本次研究對赤狐ESR1基因啟動子區的序列與結構進行生物信息學分析,有助于闡明其功能及轉錄調控機制,為進一步揭示赤狐ESR1基因作用機理和轉錄調控網絡鑒定提供參考依據。
本次研究成功克隆了ESR1基因啟動區序列,并將其成功構建至雙熒光素酶報告基因載體。有研究發現,若克隆片段中只含轉錄起始位點上游區域可能會導致實驗結果的假陰性,而克隆片段中包含轉錄起始位點、ATG或部分CDS區序列會很大的提高實驗結果的陽性率[15]。故本次研究克隆的片段中包含轉錄起始位點下游78 bp序列,符合上述要求,實驗擴增出的ESR1基因啟動子區序列與預測基本一致。基因表達是靠轉錄因子來進行調控的,不同的轉錄因子有不同的轉錄調控元件,靠與特定的啟動子區域結合而改變mRNA的轉錄水平[13]。Sp1是哺乳動物基因轉錄活化的必需因子,可通過識別基因啟動子上富含GC序列的位點發揮作用,其結合位點在轉錄激活的結構域中分布非常廣泛,且在哺乳動物大量的基因表達和細胞生物學的調控進程中發揮作用[16~19]。研究已證明,Sp1對于小鼠胚胎發育表達的第一個基因HSP70.1的轉錄來說是必需的[20];除此之外,在基因的表達調控中,Sp1具有協同激活轉錄的功能,即轉錄水平在有兩個Sp1轉錄結合位點存在時會大大提高[21]。NF-1轉錄因子是重要的轉錄因子之一,通過與CAAT box結合從而影響基因的表達,目前已有研究表明,在NF-1與以RNA聚合酶Ⅱ為轉錄起始復合物的形成有緊密聯系[22],這說明NF-1可能在ESR1基因的表達中發揮作用。
本次研究運用多個生物信息學軟件對赤狐雌激素受體ESR1基因候選啟動子區域和轉錄因子結合位點進行預測分析,增加了預測結果的可靠性,為后續的ESR1基因的研究奠定理論基礎,后續還需要對啟動子核心區域及轉錄因子的調控作用等進行瞬時轉染(Transient Transfection)試驗驗證,對轉錄因子結合位點還可進行染色質免疫共沉淀(Chromatin Immunoprecipitation, ChIP)技術、凝膠阻滯分析(Electrophretic Mobility Shift Assay, EMSA)試驗和DNA足紋分析(DNase footprinting)等驗證其具體作用及功能。
