膽堿對奶牛肝臟脂肪代謝的調控作用及機制
2021-05-12 14:09:16劉喆佳王弘浩姚軍虎曹陽春
草業科學 2021年4期
劉喆佳,王弘浩,姚軍虎,曹陽春
(西北農林科技大學動物科技學院, 陜西 楊凌 712100)
隨著科學技術在畜牧業領域的不斷創新和發展應用,集約化、現代化、規模化養殖已十分普遍,畜禽生產周期大幅度縮短,生產效率快速提高,而動物疾病的發生制約著畜牧業生產的進一步發展,因此,動物常見疾病的防控與治療對于進一步提高畜牧業生產效益十分重要,仍是畜牧業生產過程中急需解決的問題[1]。在奶牛生產養殖過程中,處于圍產期的奶牛易發生酮病和脂肪肝等肝臟脂肪代謝疾病[2],導致肝臟功能受損,極大地降低了奶產品品質和生產效益,嚴重制約我國奶業的高速發展[3]。
膽堿作為動物生長發育所必需的營養素,在動物體內可構成多種生理活性物質,參與機體代謝。一般情況下,奶牛體內可以自身合成一定量的膽堿滿足基礎需要,但對于高產奶牛尤其是在泌乳高峰期,需要更多的膽堿維持其生產性能,因此,奶牛生產往往需要額外補充適量膽堿[4]。由于膽堿在瘤胃的易降解性,市場上主要以過瘤胃膽堿(rumen-protected choline,RPC)的形式添加到奶牛飼料中[5],飼糧中補充膽堿能夠有效降低奶牛肝臟脂肪沉積,達到治療和預防脂肪肝的效果,從而提高生產性能[6]。大量研究已證明膽堿在調節奶牛肝臟脂肪代謝方面具有重要作用,對其作用機制的探索也受到越來越多的關注,主要集中在AMPKα (adenosine monophosphateactivated protein kinase α)信號通路的激活和膽堿的一碳單位循環兩種途徑。本研究闡述了膽堿的基本性質及生理功能,圍繞膽堿在奶牛肝臟脂肪代謝中的調控作用及機制進行綜述。
1 膽堿的基本性質和生理功能
膽堿(三甲基乙醇胺),分子式為(CH3)3N(CH2)2OH,是一種吸濕性較強的季胺堿,質地是無色粘稠,并略帶魚腥味的白色易潮解的針狀晶體。膽堿在動植體內多以種不同的形式存在,主要有乙酰膽堿、磷脂酰膽堿、神經鞘磷脂等形式。在飼料中也包含多種形式的膽堿,主要以磷脂酰膽堿的形式存在[7],以衍生物和游離態形式存在的膽堿比較少[8]。正常情況下,動物體內可自身合成膽堿,在一些特殊的生理狀態下,如奶牛圍產期能量負平衡狀態,需要在飼料中額外補充適量膽堿,以保證奶牛生理功能正常。為保證穩定性,動物生產上通常以氯化膽堿的形式作為添加劑添加到日糧中[9]。
膽堿是動物生長發育所必需的營養素,在動物體內以多種形式存在并發揮不同的作用,其在代謝中的主要生理功能包括[10-12]:1) 維持生物膜組成結構完整。膽堿能夠在機體內合成卵磷脂和神經鞘磷脂,參與構成細胞膜結構和脂蛋白,維持細胞正常生理功能和結構完整性;2) 促進體內甲基代謝。膽堿作為甲基供體,可提供不穩態甲基,與胍基乙酸生成肌酸,還可生成肉堿、腎上腺素等活性物質,參與機體甲基代謝。膽堿在機體內,能夠氧化轉化為甜菜堿,甜菜堿作為甲基供體可通過甲基化反應生成蛋氨酸,蛋氨酸可以繼續將甲基傳遞給S-腺苷甲硫氨酸(S-adenosylmethionine,SAM-e),在機體內參與一碳單位的代謝循環。膽堿、甜菜堿和蛋氨酸作為主要的甲基供體,三者多數情況下能夠在體內進行補充或替代;3) 保證信號分子的傳遞。膽堿參與乙酰膽堿和磷脂酰膽堿的合成,作為神經遞質維持基本的神經信號傳遞過程;4) 促進腦部正常生長發育和提高記憶力;5) 促進脂肪代謝。膽堿能夠通過促進載脂蛋白的合成,加快肝臟脂肪向外周轉運,進而降低脂肪在肝臟的沉積;6) 調控細胞凋亡[11]。
2 膽堿對奶牛肝臟脂肪代謝的調控作用
圍產期奶牛受特殊生理階段的生理應激影響,干物質采食量明顯下降,而圍產前期胎兒的快速發育和圍產后期奶牛的泌乳均需要大量的能量。為滿足圍產期的能量需求,奶牛主要通過動員體脂來進行自身代謝的調控,采食量的嚴重不足和過量的體脂動員極易使奶牛處于能量負平衡(negative energy balance,NEB)狀態[13]。奶牛體脂動員會分解產生的大量非酯化脂肪酸(non-esterified fatty acids,NEFA)通過血液進入到肝臟進行代謝,主要有以下3 條途徑:1) 完全氧化進行供能,生成二氧化碳和水;2) 當生成的NEFA 的量超出肝臟的氧化能力時,一部分NEFA 會不完全氧化生成酮體,包括β-羥丁酸(βhydroxybutyric acid,BHBA)、丙酮和乙酰乙酸,酮體積累過多易引發酮病;3) 再酯化生成甘油三酯(triglycerides, TG),與載脂蛋白結合以極低密度脂蛋白(very low density lipoprotein,VLDL)形式轉運出肝臟,由于肝臟轉運TG 能力有限,導致過量的TG 積累在肝臟,易形成脂肪肝。酮病和脂肪肝等肝臟代謝性疾病會造成肝臟功能受損,加劇奶牛NEB,直接影響奶牛的生產性能,降低奶牛的生產效率和奶產品品質,增加了飼養成本,減少了企業生產效益[14]。
許多研究表明,飼糧中補充RPC 對奶牛有益,能夠增加奶牛圍產期采食量[15],降低肝臟脂肪沉積[16],調控能量代謝[17],增強機體抗氧化應激能力[18],提高生產性能[19]。Cooke 等[20]在誘導奶牛脂肪肝過程中每天補充15 g RPC,處理組血液中NEFA 濃度降低,肝臟中TG 含量降低;馬晨等[21]發現,圍產期奶牛基礎日糧中添加25 g·d-1膽堿效果最好,顯著提高了奶牛產奶量,降低了血液中NEFA 和BHBA 的含量,提高奶牛繁殖性能;其他研究人員[22-23]也證實補充RPC 一定程度上可以提高產奶量,降低肝臟脂肪沉積。最新研究發現,飼喂RPC 的起始時間和持續時間對泌乳奶牛的生產性能會產生不同的影響,當RPC 補充發生在圍產期時,提供12.9 g·d-1的膽堿有利于奶牛的生產性能,但在產后22 d 后補充RPC 沒有觀察到額外的益處[24]。以上研究均表明膽堿對奶牛肝臟脂肪代謝有調控作用,能夠緩解能量負平衡狀態,達到預防和治療脂肪肝的效果,但對于膽堿的最適添加量和最佳飼喂時間還需要進一步研究。
膽堿主要通過上調脂肪分解和下調脂肪生成途徑的基因和蛋白的表達水平來調節脂肪代謝,降低奶牛肝臟脂肪沉積,分別從脂肪代謝的3 個方面進行,包括減少脂肪合成,促進脂肪氧化,促進脂肪轉運。
2.1 對脂肪合成的調控
膽堿可能通過調節脂肪分解基因減少脂肪合成。過氧化物酶體增殖物激活受體γ (peroxisome proliferator-activated receptor gamma, PPARγ)是與脂肪生成有關的轉錄因子,已知脂蛋白酯酶(lipoprotein lipase,LPL)基因是其下游基因[25],LPL能夠水解乳糜微粒和極低密度脂蛋白中甘油三酯的生成脂肪酸和甘油的關鍵限速酶,促進細胞從極低密度脂蛋白中攝取脂肪,Goselink 等[26]在奶牛活體采樣試驗中發現,由于沒有觀察到LPL 基因表達的變化,補充RPC 誘導的肝脂質沉積減少不能歸因于在脂肪組織中改變脂肪分解。硬脂酰輔酶A 去飽和酶(stearyl coenzyme A dehydrogenase,SCD)是催化飽和脂肪酸向單不飽和脂肪酸轉化的重要限速酶,在脂肪合成和分解及能量代謝中起重要的調控作用,在王艷輝[27]的試驗中證明了添加膽堿組相對于對照組硬脂酸含量下調,說明膽堿以通過調控SCD 基因表達,減少脂肪沉積。膽堿還可下調脂肪合成,減少脂肪沉積。乙酰輔酶A 羧化酶(acetyl-CoA carboxylase,ACCα)和脂肪酸合成酶(fatty acid synthase,FAS)參與脂肪酸的合成過程,是脂肪酸合成的關鍵酶[28]。孫菲菲[29]在研究中發現,添加膽堿可以明顯抑制ACC和FAS 的基因表達量,減少肝細胞脂肪蓄積。
2.2 對脂肪氧化的調控
脂肪酸氧化主要是在線粒體中進行β-氧化,游離脂肪酸進入線粒體基質的過程是脂肪酸進行β-氧化的限速環節,而肉堿棕櫚酰轉移酶1A(carnitine palmitoyltransferase,CPT1A)是脂肪酸進行β-氧化的關鍵限速酶,能夠催化細胞內脂肪酰輔酶A 轉化為脂肪酰肉堿進入線粒體[30-31]。肉堿是重要跨膜轉運載體,在脂肪酸氧化過程中起關鍵作用,能夠幫助長鏈脂肪酸進入到細胞線粒體內。Coleman 等[32]在奶牛能量負平衡狀態期間,給奶牛補充膽堿發現肝臟TG 含量降低,CPT1A 顯著增加,表明線粒體β-氧化能力增加。在日糧中添加膽堿,可通過增加肉堿轉運蛋白SLC22A5 的表達[26],促進肉堿轉運輸送,增加動物肝臟中肉堿水平[33],進而促進脂肪酸轉運到線粒體進行氧化分解[34]。
2.3 對脂肪轉運的調控
肝臟中的脂肪主要以VLDL 的形式向外周轉運[35],磷脂酰膽堿是合成VLDL 的重要成分[36],對脂肪轉運過程起重要作用。膽堿通過兩種途徑生成磷脂酰膽堿:肝臟中70% 的膽堿通過CDP 這一途徑合成,30% 通過PEMT 體內生成途徑合成,當膽堿外源攝入不足時,PEMT 途徑供應肝臟內的磷脂酰膽堿發揮主要作用,但同時需要SAM-e 的參與[37]。而VLDL 的合成、組裝與分泌還需要載脂蛋白B(apolipoprotein B100,Apo B)[38]和微粒體甘油三酯轉運蛋白(microsomal triglyceride transfer protein,MTTP)[39]的協助完成。ApoB 是合成VLDL 所必需的蛋白質,MTTP 能夠幫助脂質轉移到內質網中,使新生的ApoB 與其結合,并通過形成原始脂蛋白顆粒運輸到胞外。有研究發現,增加膽堿的添加量可以通過調節PPARα 通路上調MTTP 和ApoB-100 的表達水平,進而增加VLDL 的合成、組裝與分泌,促進脂肪轉運,減少肝臟脂肪沉積[40]。載脂蛋白A5 (apolipoprotein A5,ApoA5)與VLDL 和高密度脂蛋白相關,并且參與降低肝臟脂肪含量的過程[41]。已有研究表明,隨著膽堿供應增加,ApoA5 也隨之線性增加,說明膽堿可能通過提高ApoA5 的表達來減少肝臟脂肪沉積[32]。脂肪酸轉運蛋白(fatty acid transport protein,FATP)是參與調控脂肪酸的轉運和脂肪代謝的關鍵因子,能夠協助長鏈脂肪酸跨膜轉運,調節脂肪沉積。Goselink 等[26]發現補充膽堿可提高FATP5 的表達量,緩解奶牛產后能量負平衡,證明了添加膽堿能夠促進脂肪從肝臟向外周轉運,調節脂肪代謝。
3 膽堿對奶牛肝臟脂肪代謝的調控機制
膽堿具有很好地調節肝細胞脂代謝的作用,能夠顯著降低肝臟脂肪沉積,達到緩解奶牛能量負平衡的效果。目前,對膽堿調節奶牛肝細胞脂代謝機制的研究有限,主要通過激活AMPK 信號通路和膽堿的一碳單位循環兩個途徑(圖1)。
3.1 AMPKα 介導的膽堿調控脂肪代謝
AMPK 是重要的細胞能量代謝調控元件,激活AMPK 通路可通過磷酸化或去磷酸化調節下游分解和合成代謝相關的調節因子或蛋白的表達,進而實現對細胞能量代謝的調控[42],PPARα、LXRα 和SREBP-1c 的表達均受到AMPKα 的調控[43]。有研究發 現,1.5 mmol·L-1的NEFA 誘 導 的LO2 肝 細 胞 損傷模型可抑制細胞能量和脂質代謝,增加細胞內TG 的沉積,降低CPT-1α、ACC、FAS 的mRNA 表達量,并且能夠抑制AMPKα 的表達[44],當在此模型中添加膽堿后,促進了AMPKα 磷酸化,該作用能被BML-275 (一種AMPKα 抑制劑) 阻斷,并能夠增加PPARα、CPT-1α 和ApoB-100 的基因表達水平,降低LXRα 和SREBP-1c 的表達,同時抑制ACCα 和FAS的表達,而BML 只阻斷了LXRα 的表達,說明膽堿可以通過激活AMPKα 磷酸化調節PPARα 和LXRα的轉錄活性,并獨立于AMPKα 調節SREBP-1c,以促進NEFA 處理的肝細胞中脂質氧化和轉運,實現對肝臟細胞脂代謝調控作用[45](圖2)。以上研究證明AMPKα 可通過調節其下游PPARα、LXRα和SREBP-1c的表達水平介導膽堿調控肝臟細胞脂肪代謝。
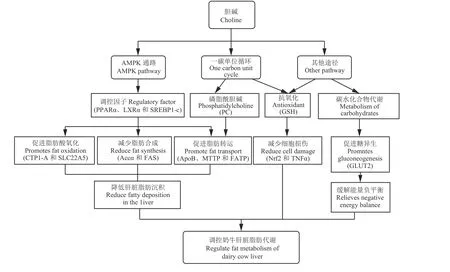
圖 1 膽堿對奶牛肝臟脂肪代謝的調控途徑Figure 1 The regulation of choline on fat metabolism in dairy cow livers
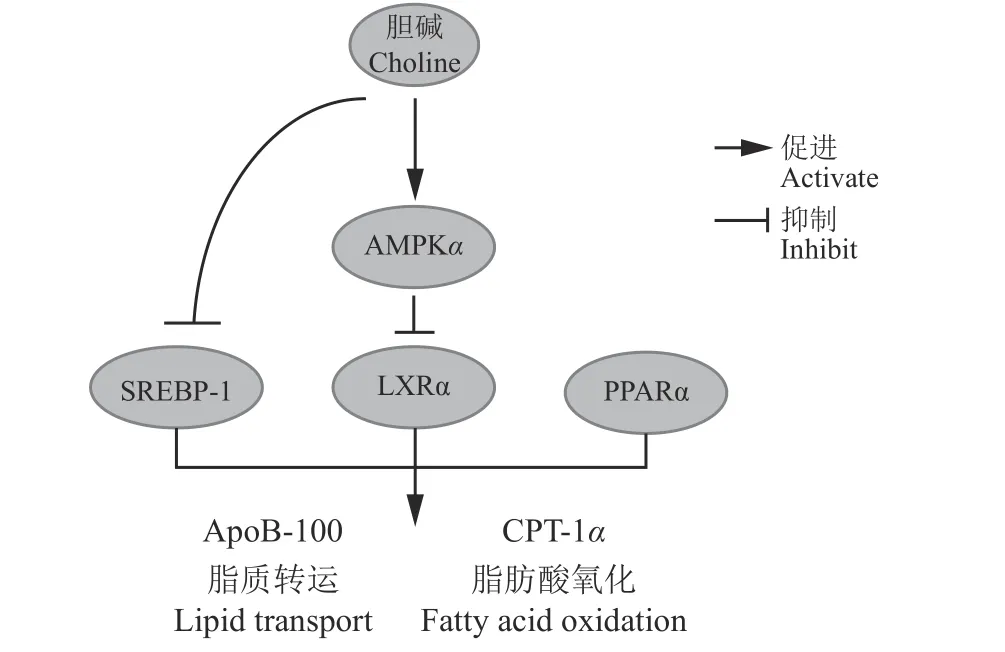
圖 2 膽堿和蛋氨酸對肝臟代謝的調節機制Figure 2 The regulatory mechanism of hepatic metabolism by choline and methionine
還有其他研究發現,膽堿對肝臟脂代謝的調節不能通過影響PPARα 激活來實現,而是通過提高PPARδ 的表達來降低脂肪在肝臟的沉積[26]。膽堿調控下游調控因子可能還存在其他途徑,LXRα-SREBP-1c通路可以不依賴 AMPKα 來調控肝細胞脂肪代謝和緩解脂肪肝[46-47]。膽堿通過PPAR、LXRα和SREBP-1c調控肝臟細胞脂代謝是否僅依賴AMPK 信號通路以及是否還有其他調節途徑還需進一步研究驗證。
3.2 膽堿的一碳單位循環
膽堿在體內可以被氧化,轉化生成甜菜堿,甜菜堿作為甲基供體在甲基轉移酶的作用下,通過甲基化反應給同型半胱氨酸提供一個甲基生成蛋氨酸[48-49]。有研究發現,蛋氨酸還能在肝臟內各種酶的參與下合成肉堿,刺激 CPT-1A 的表達[50-51],進而能夠促進脂肪酸進入到線粒體進行β-氧化,減少脂肪沉積。蛋氨酸還能轉化為SAM-e[52],SAM-e 給磷脂酰乙醇胺提供游離甲基參與體內磷脂酰膽堿的合成[53],構成VLDL,幫助脂肪轉運出肝臟,降低肝臟脂肪沉積(圖3)。還有研究表明,膽堿通過一碳單位循環生成一些非酶抗氧化物質,如谷胱甘肽[54-55],提高細胞抗氧化能力,減少氧化應激和炎性損傷的發生,維持肝細胞代謝功能正常進而緩解能量負平衡,間接促進肝細胞脂質代謝,此結論還需要更多試驗驗證。
3.3 其他途徑
研究表明,補充了過瘤胃膽堿的母牛在產犢后,表現出肝臟葡萄糖轉運載體2 (GLUT2)的mRNA表達量顯著提高[26],并且丙酮酸羧化酶 (PC)的mRNA 表達水平降低,說明膽堿的補充很可能促進了肝臟的糖異生能力[56],促進碳水化合物在奶牛體內的代謝利用,減少脂肪過度動員,進而促進能量平衡,緩解奶牛的能量負平衡狀態,維持了肝細胞正常的脂代謝功能。試驗中還發現,膽堿提高了高濃度NEFA 模型下肝細胞氧化應激關鍵因子Nrf2[57]的表達量,并且降低了炎癥因子TNF-α 的表達[29],說明膽堿可能增強了肝細胞的氧化能力,緩解了能量負平衡狀態下的氧化應激和炎癥反應,減少肝細胞損傷,促進了肝細胞脂代謝作用。
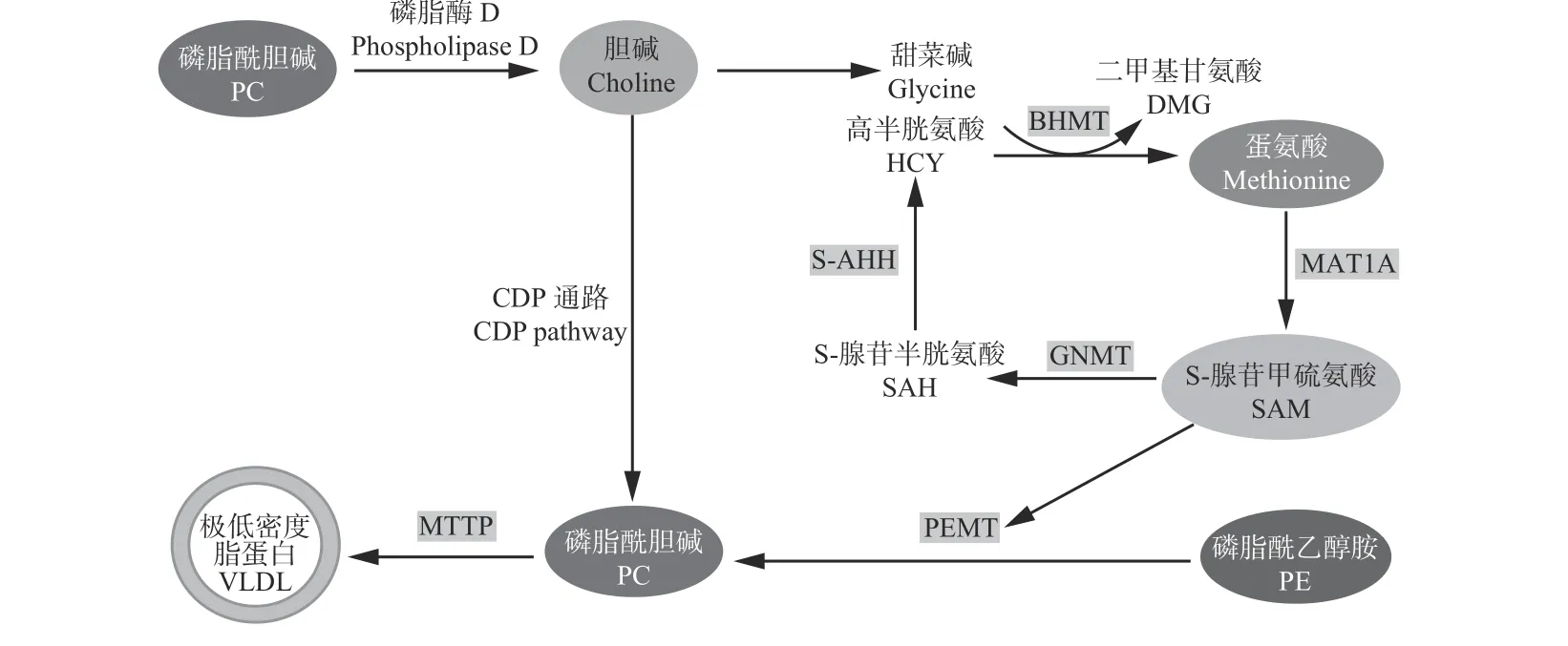
圖 3 轉甲基化循環中膽堿和蛋氨酸代謝途徑與控制甲基轉移的關鍵酶之間的交叉Figure 3 Intersection between pathways of choline and methionine metabolism in the transmethylation cycle and key enzymes that control methyl group transfer
4 小結
奶牛在能量負平衡狀態時,是脂肪肝等營養代謝性疾病的高發期,膽堿作為一種營養素,對調節奶牛肝臟脂肪代謝具有重要作用,補充膽堿可有效改善奶牛脂肪肝。膽堿主要是通過上調脂肪分解和下調脂肪生成途徑基因和蛋白的表達水平來調節脂肪代謝,其具體的調節機制已有研究,主要集中在AMPK 信號通路的激活和膽堿的一碳單位循環途徑(圖3),各調節因子的效應次序和模式還有待進一步研究確認,明晰膽堿調控奶牛肝臟脂代謝的核心關鍵通路及機制,從而確定合適的膽堿添加量及添加方式,對奶牛有效營養調控和提高生產效益具有重要價值。
