不同樹種林下套種金花茶土壤微生物多樣性分析
2021-05-14 02:18:40伍思宇李寶財李紀元廖健明韋曉娟傅鏡遠李開祥
廣西林業科學 2021年2期
伍思宇,李寶財,李紀元,廖健明,韋曉娟,傅鏡遠,李開祥
(1.廣西壯族自治區林業科學研究院 廣西特色經濟林培育與利用重點實驗室,廣西南寧530002;2.中國林業科學研究院亞熱帶林業研究所,浙江杭州 311400;3.合浦佳永金花茶開發有限公司,廣西合浦 536100)
金花茶(Camellia nitidissima)屬小喬木或常綠灌木,高度為2 ~6 m[1],主要分布在中國廣西南部和越南北部[2],原始生境的伴生植物主要有白櫟(Quer?cus fabri)、水冬哥(Saurauia tristyla)和白顏樹(Giron?niera subaequalis)等[3]。金花茶是山茶屬稀有的黃色種質資源,含有豐富的黃酮類、多糖和茶多酚等化學活性成分以及鍺(Ge)和砷(Se)等微量元素[4-5],具有極高的觀賞和科研價值[6]。
土壤微生物是森林生態系統的重要組成部分,在養分轉化和循環上發揮重要的作用[7],其多樣性受林分、枯落物和土壤因子等因素共同影響[8-10]。Waid[11]指出植被類型和數量不同,林下凋落物及根系分泌物也不同,使土壤微生物發生變化。宋賢沖等[12]研究表明,土壤有機質、全磷(P)和全鉀(K)含量等土壤理化性質可能是造成不同季節常綠闊葉林土壤微生物群落功能多樣性差異的主要原因。
林下套種是通過空間將目標樹種與其他植物組合在一起的栽培模式,不僅能充分利用林地空間,增加森林單產,還能促進養分循環,提高土壤肥力和土壤微生物數量[13]。吉艷芝等[14]研究表明與落葉松(Larix gmelinii)純林相比,落葉松混交林的土壤養分、微生物多樣性和酶活性均得到了顯著提高。由于金花茶耐陰的特性[3],研究人員采取林下套種金花茶的栽培模式,研究金花茶的生長和光合特性[15-16],但鮮少關注林下的土壤特性。本研究對不同樹種林下套種金花茶的土壤微生物多樣性進行研究,探討土壤微生物群落結構的差異,以及枯落物和土壤理化性質對土壤微生物多樣性的影響,以期為林下套種金花茶選擇適宜的上層樹種提供理論依據和指導。
1 材料與方法
1.1 試驗地概況
試驗地位于廣西合浦縣佳永金花茶開發有限公司龍門江基地(109°21'E,21°67'N),海拔12 m,屬亞熱帶海洋季節性氣候,年均氣溫22.4 ℃,年均降水量1 667 mm,年均相對濕度75% ~86%。選擇造林時間相近的3 種樹種套種金花茶,對枯落物和土壤進行調查研究。3 種樹種分別為印度紫檀(Ptero?carpus indicus)、荔 枝(Litchi chinensis)和 濕 地 松(Pinus elliottii)(表1)。印度紫檀為高大落葉喬木[17],林下透光度在春季、夏季、秋季和冬季分別為20%、12.5%、44.8%和100%;荔枝為常綠喬木[18],林下透光度常年為5%左右;濕地松為常綠針葉喬木[19],林下透光度常年為60%左右。
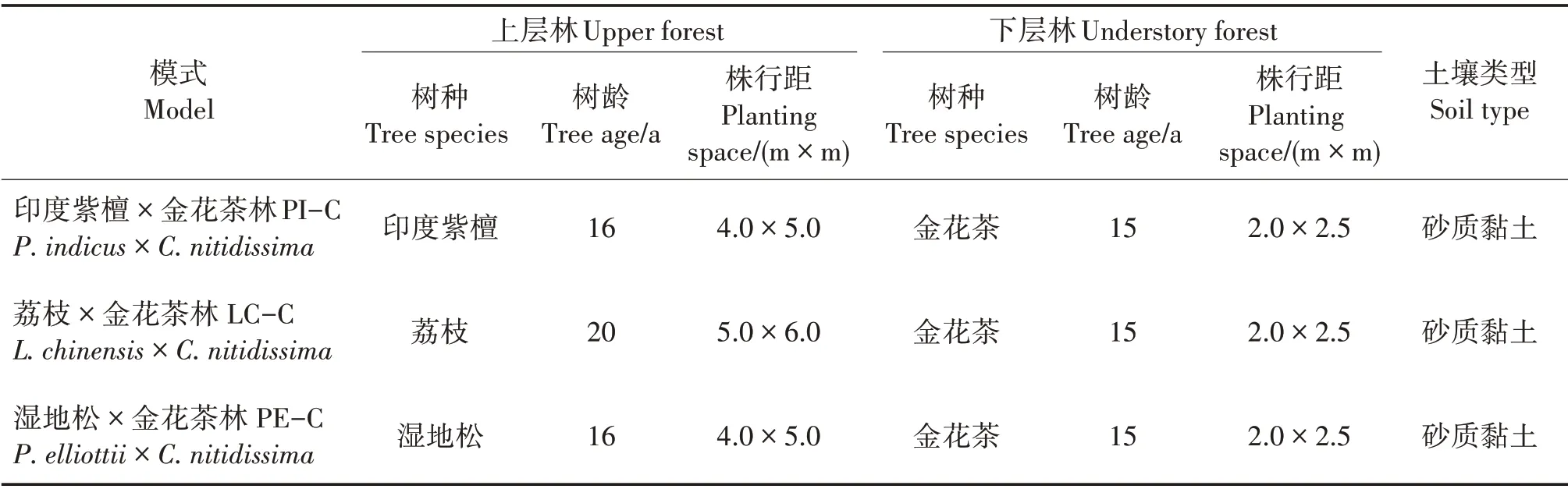
表1 不同套種模式基本概況Tab.1 General situation of different interplanting models
1.2 樣品采集
2018年12月—2019年12月,每3 個月采集1次枯落物。收集面積為1 m2的枯枝落葉,風干后稱重,用于測定枯落物蓄積量,另取部分枯枝落葉樣品帶回室內測定養分。
2019年6月采集土壤,測定土壤理化性質和微生物。在3 種樹種的林地分別隨機選取3 個采樣點,清除表層雜質后,用環刀取出深度為0 ~10 cm的土壤,用于土壤物理性質分析。于平行斷面下鏟取土樣,剔除石礫和植物殘根等雜物后,將采集的土壤分別混合均勻,采用四分法,將一部分裝入15 mL 無菌離心管中冷藏,用于土壤微生物分析;另一部分置于自封袋中,用于土壤化學性質分析。
1.3 樣品測定和數據分析
用稱重法測枯落物蓄積量[20];凱氏定氮法測全氮(N)含量,鉬銻抗比色法測全P 含量,火焰原子分光光度法測全K 含量[21]。用烘干法測土壤含水量,環刀法測土壤容重和土壤孔隙度[22];用電位法測土壤pH 值;用重鉻酸鉀-硫酸消化法測土壤有機質含量,用凱氏定氮法測土壤全N含量,用鉬銻抗比色法測土壤全P 含量,用酸溶-火焰光度法法測土壤全K含量[23]。采用Excel和SPSS 19.0軟件進行數據處理。
用Illumina-MiSeq 高通量測序技術(上海美吉生物醫藥科技有限公司)進行土壤微生物群落基因組測序。采用E.Z.N.A.?soil DNA Kit 提取試劑盒(Omega Bio-Tek,Norcross,GA,U.S.)提取土壤總DNA。細菌V3 ~V4可變區采用338F(5’-ACTCCTACGGGAGGCAGCAG-3’)和806R(5’-GGACTACHVGGGTWTCTAAT-3’)引物進行PCR擴增,真菌18S rDNA 的V5 ~V7區采用SSU0817F(5’-TTAGCATGGAATAATRRAATAGGA-3’)和1196R(5’-TCTGGACCTGGTGAGTTTCC-3’)引物進行擴增。將PCR 擴增產物用2%瓊脂糖凝膠電泳檢測,使用AxyPrepDNA 凝膠回收試劑盒(AXYGEN 公司)切膠回收PCR 產物,Tris-HCl 洗脫,1%瓊脂糖電泳檢測。再將PCR 回收產物用QuantiFluor?-ST 藍色熒光定量系統(Promega公司)進行檢測定量,之后按照測序量,進行相應比例的混合。先連接“Y”字形連接器,再用磁珠過濾,自連接片段,然后通過PCR 擴增文庫模板,對NaOH變性,產生單鏈DNA片段。利用測序公司MiseqPE300平臺進行測序(上海美吉生物制藥有限公司)。將原始數據上傳至NCBI數據庫進行比對,并在美吉I-Sanger 云數據分析平臺上進行在線數據分析(www.majorbio.com)。
2 結果與分析
2.1 不同套種模式枯落物蓄積量和養分分析
枯落物累計年限相近時,不同林分枯落物蓄積量主要與枯落物的輸入量和分解速率有關[20]。3 種套種處理的枯落物單位面積蓄積量均呈先減少后增加的趨勢,說明枯落物隨著時間的推移會逐步分解,待到新一輪落葉期時,枯落物又會再次累積(圖1a)。在枯落物分解的過程中,枯落物的養分貯量也會隨之變化。PI-C 枯落物的全N、全P 和全K 含量隨時間呈波動變化(圖1b ~d),LC-C 和PE-C 全年變化趨勢均不太明顯。PI-C枯落物全N含量呈先增多后減少再增多的趨勢,年變化量為16.36 g/kg,全年均高于LC-C和PE-C,PI-C、LC-C和PE-C凋落物全N含量 分別 為13.39 ~29.75、10.68 ~14.36 和4.90 ~8.38 g/kg。PI-C 枯落物全P 含量呈先增多后減少的趨勢,年變化量為0.51 g/kg。PI-C、LC-C 和PE-C 枯落物全P含量分別為0.55 ~1.06、0.49 ~0.68和0.20 ~0.31 g/kg,PI-C 枯落物全P 含量存在明顯優勢。PIC 枯落物全K 含量呈先減少后增多的趨勢,年變化量為3.52 g/kg,全K 含量最大值為5.15 g/kg,分別為LC-C和PE-C全K含量最大值的1.40和2.83倍。
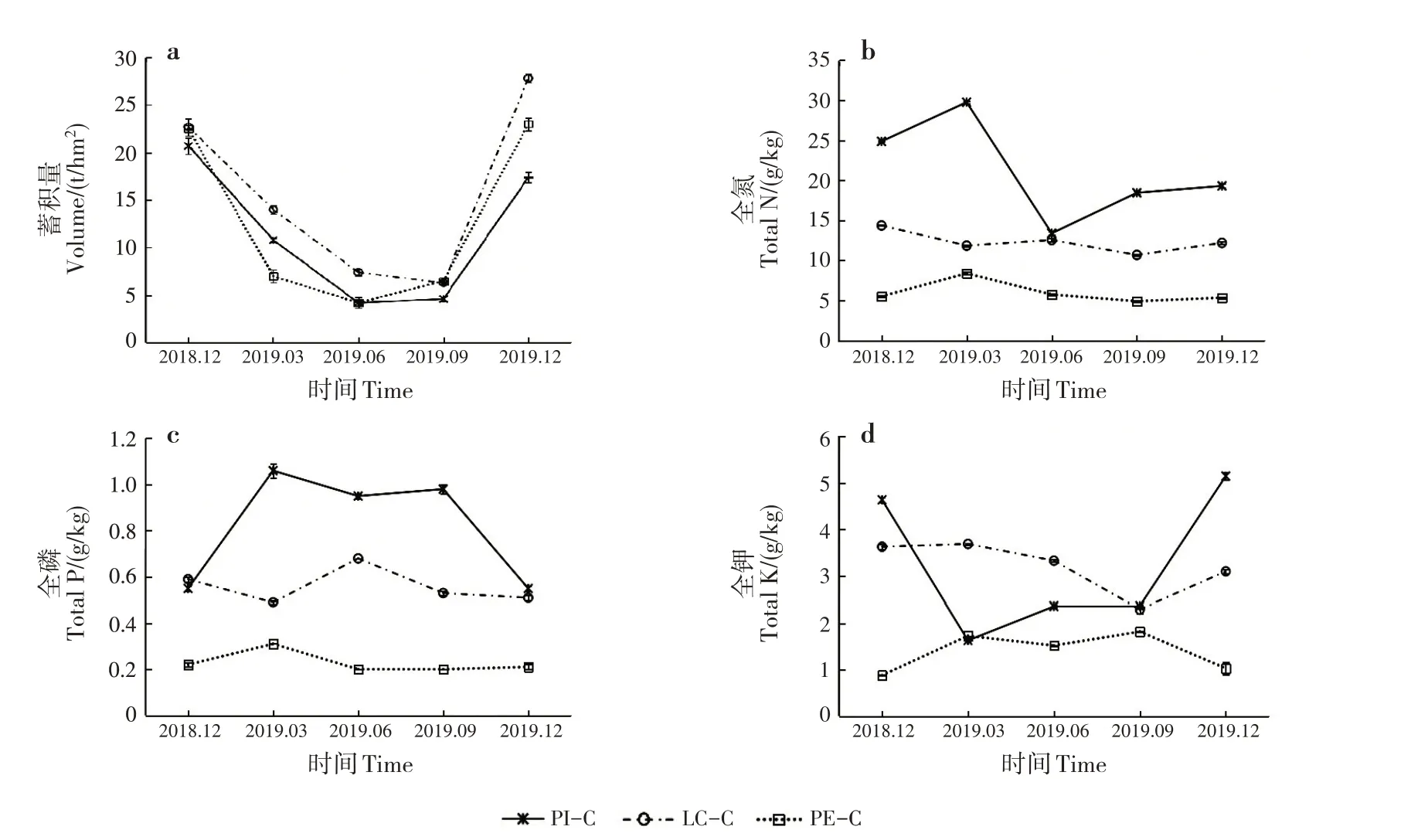
圖1 不同套種模式的枯落物蓄積量和養分貯量動態變化Fig.1 Dynamic changes of litter volume and nutrient storage of different interplanting models
2.2 不同套種模式土壤理化性質分析
不同套種模式對土壤的最小持水量、毛管持水量、非毛管孔隙、毛管孔隙、總孔隙度、pH、全P 和全K 含量影響顯著(P< 0.05),對土壤容重、最大持水量、有機質和全N含量影響不顯著(表2)。土壤孔隙度能反映出土壤透水通氣性能是否良好[24],PI-C 土壤總孔隙度最大,與LC-C 和PE-C 差異顯著(P<0.05),說明PI-C 土壤透水通氣性最好。3 種土壤均為酸性土壤,其中PE-C 土壤pH 為4.70,與PI-C 和LC-C 差異顯著(P<0.05)。LC-C 土壤全P 含量最高(1.08 g/kg),與PE-C差異顯著(P<0.05),與PI-C差異不顯著;LC-C 土壤全K 含量最高(2.99 g/kg),與PI-C和LC-C均差異顯著(P<0.05)。
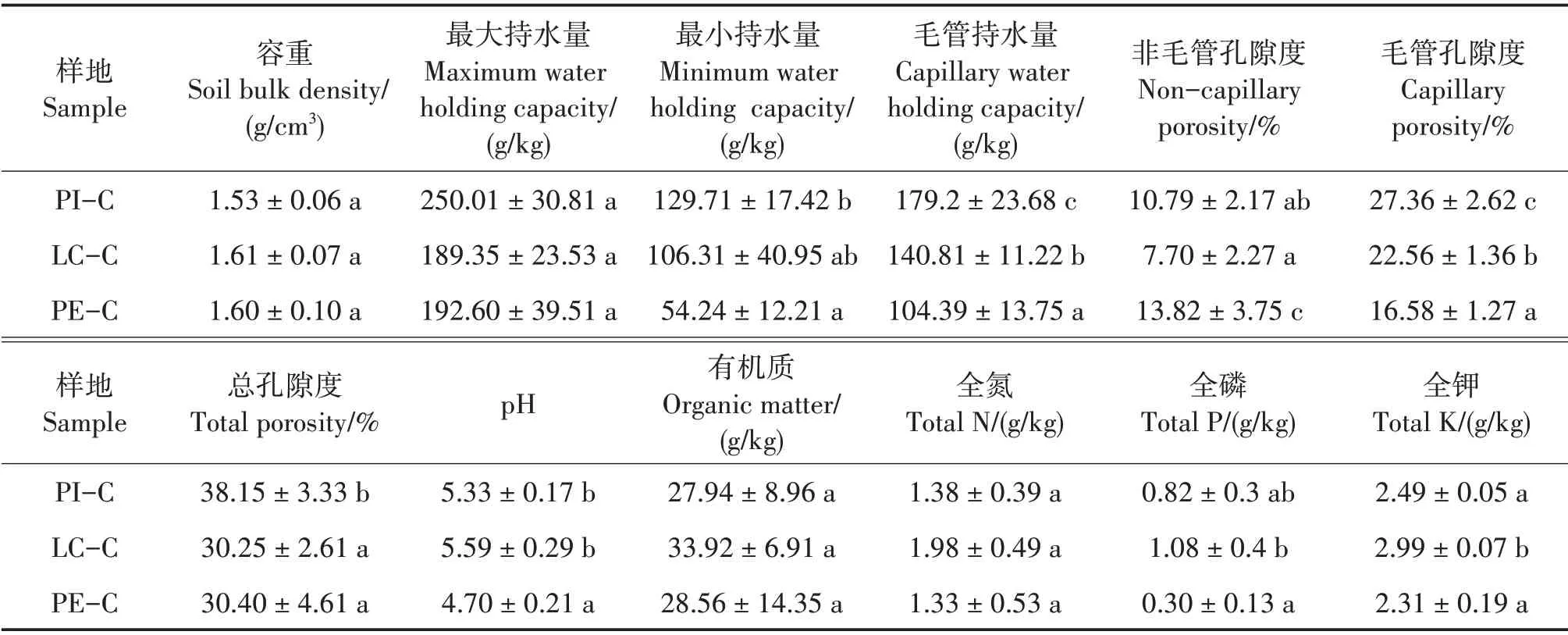
表2 不同套種模式的土壤理化性質Tab.2 Soil physical and chemical properties of different interplanting models
2.3 不同套種模式土壤微生物多樣性分析
PI-C、LC-C 和PE-C 土壤細菌群落各獲得4 471、4 310 和2 897 個OTU,真菌群落各獲得243、300 和259 個OTU,真菌細菌比分別為0.054、0.070 和0.089。對0.97%相似水平的OTU 代表序列進行分類學分析,PI-C、LC-C 和PE-C 土壤細菌群落分別歸為32 個門734 個屬、28 個門638 個屬和26個門465個屬,真菌群落分別歸為36個門124個屬、39個門136個屬和32個門117個屬。
2.3.1 不同套種模式土壤微生物Alpha多樣性分析
Chao1和Ace指數越大,說明樣本中微生物豐富度越高;Shannon 指數越大,Simpson 指數越小,說明樣本中微生物群落多樣性越高[25];Coverage 指數越高,說明樣本中序列被測出的概率越高。PI-C 和LC-C土壤細菌的Chao1、Ace和Shannon指數顯著大于PE-C(P< 0.05),且Simpson 指數顯著小于PE-C(P<0.05),說明PI-C 和LC-C 土壤細菌豐富度和多樣性明顯優于PE-C,PI-C 與LC-C 差異不顯著;PIC、LC-C 和PE-C 土壤真菌的Chao1、Ace、Shannon和Simpson 指數差異不顯著,說明3 種土壤真菌豐富度和多樣性差異不大(表3)。
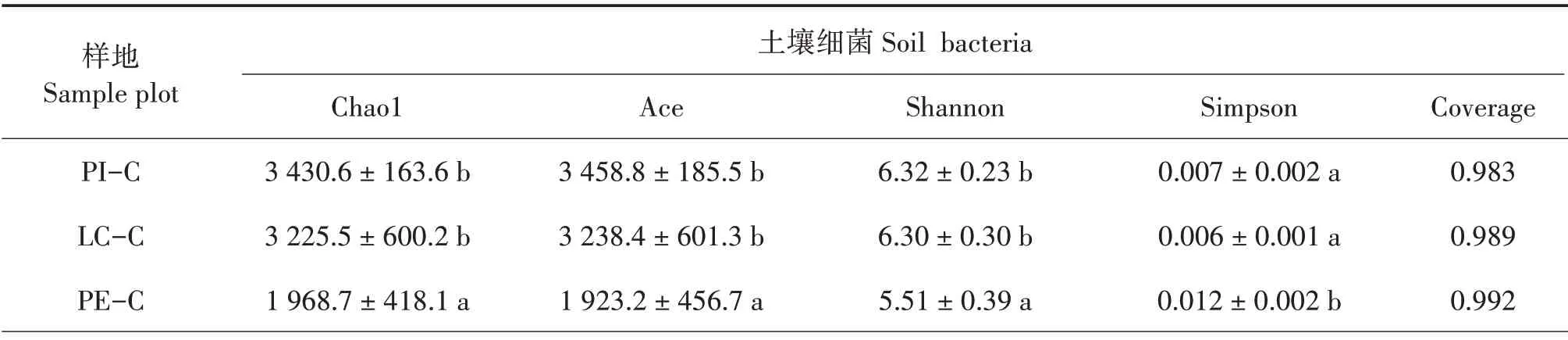
表3 不同樹種套種模式的土壤微生物多樣性指數Tab.3 Soil microbial diversity index of different interplanting models
續表3Continued
2.3.2 不同套種模式土壤微生物群落組成分析
PI-C、LC-C 和PE-C 土壤優勢細菌門分別為13、13 和12 個,其中變形菌門(Proteobacteria)、放線菌門(Actinobacteria)、酸桿菌門(Acidobacteria)和綠彎菌門(Chloroflexi)在3 種土壤中均為優勢菌群,變形菌門在3 種土壤中占比為LC-C(34.67%)>PE-C(28.35%)>PI-C(22.40%);放線菌門為PE-C(30.50%)>LC-C(23.39%)>PI-C(11.52%);酸桿菌 門 為PE-C(23.31%)>PI-C(16.07%)>LC-C(12.16%);綠彎菌門為PI-C(29.81%)>LC-C(13.48%)>PE-C(6.81%)(圖2a)。在屬分類水平上,PI-C 土壤中細菌占比最多的3 個群落分別為norank_c_AD3(19.78%)、norank_f_Xanthobacterace?ae(5.73%)和norank_o_Subgroup_2(5.04%);LC-C土壤中細菌占比最多的3個群落分別為norank_f_Xan?thobacteraceae (9.97%) 、 norank_c_Subgroup_6(4.17%)和norank_c_AD3(3.99%);PE-C 土壤中細菌占比最多的3 個群落分別為熱酸菌屬(Acidother?mus,10.95%)、norank_f_Xanthobacteraceae(8.49%)和norank_o_Acidobacteriales(6.46%)(圖2b)。
PI-C、LC-C 和PE-C 土壤優勢真菌為子囊菌門(Ascomycota)、擔子菌門(Basidiomycota)、毛霉門(Mucoromycota)和纖毛門(Ciliophora)。子囊菌門在3 種土壤中占比為PI-C(77.61%)>LC-C(62.19%)>PE-C(28.45%);擔子菌門為PE-C(63.55%)>LC-C(26.35%)>PI-C(11.91%);毛霉門 為 PI-C (5.22%) >LC-C (4.80%) >PE-C(2.49%);纖毛門為PI-C(1.46%)>LC-C(1.10%)>PE-C(0.79%)(圖2c)。PI-C、LC-C 和PE-C 土壤中占比最多的細菌和真菌群落分別為綠彎菌門和子囊菌門、變形菌門和子囊菌門、放線菌門和擔子菌門。PI-C 真菌占比最多的3 個群落為unclassi?fied_f_Aspergillaceae(16.46%)、unclassified_f_Hypo?creaceae(14.12%)和毛殼菌屬(Chaetomium,13.57%);LC-C 真菌占比最多的3 個群落為unclas?sified_f_Aspergillaceae(17.45%)、Saitozyma(15.42%)和unclassified_f_Hypocreaceae(7.75%);PE-C 真菌占比最多的3 個群落為Saitozyma(55.68%)、unclas?sified_f_Hypocreaceae(8.26%)和毛殼菌屬(6.10%)(圖2d)。
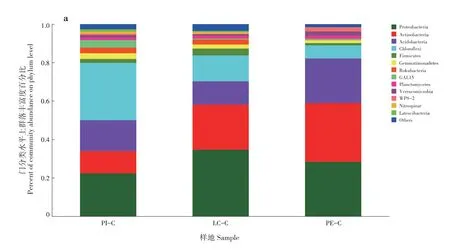
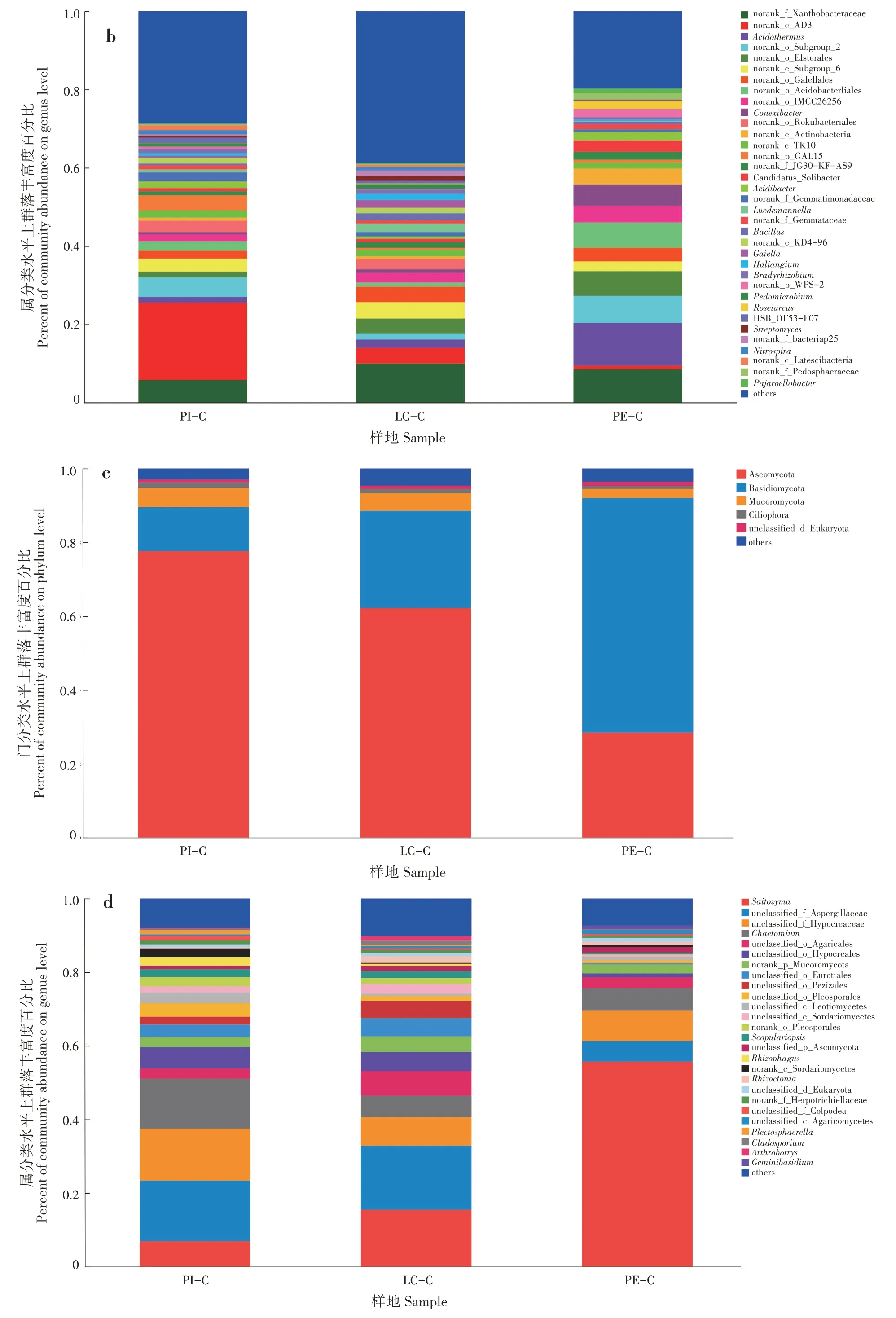
圖2 不同套種模式土壤微生物群落(a和b:細菌;c和d:真菌)Fig.2 Soil microbial community of different interplanting models(a and b:bacteria;c and d:fungi)
3 種套種模式土壤中共有的細菌屬和真菌屬分別為351個和82個(圖3)。PI-C 土壤中特有的細菌屬為176 個,多于LC-C(25 個)和PE-C(14 個)(圖3a)。PI-C、LC-C 和PE-C 土壤中特有的真菌屬分別為17個、25個和11個(圖3b)。
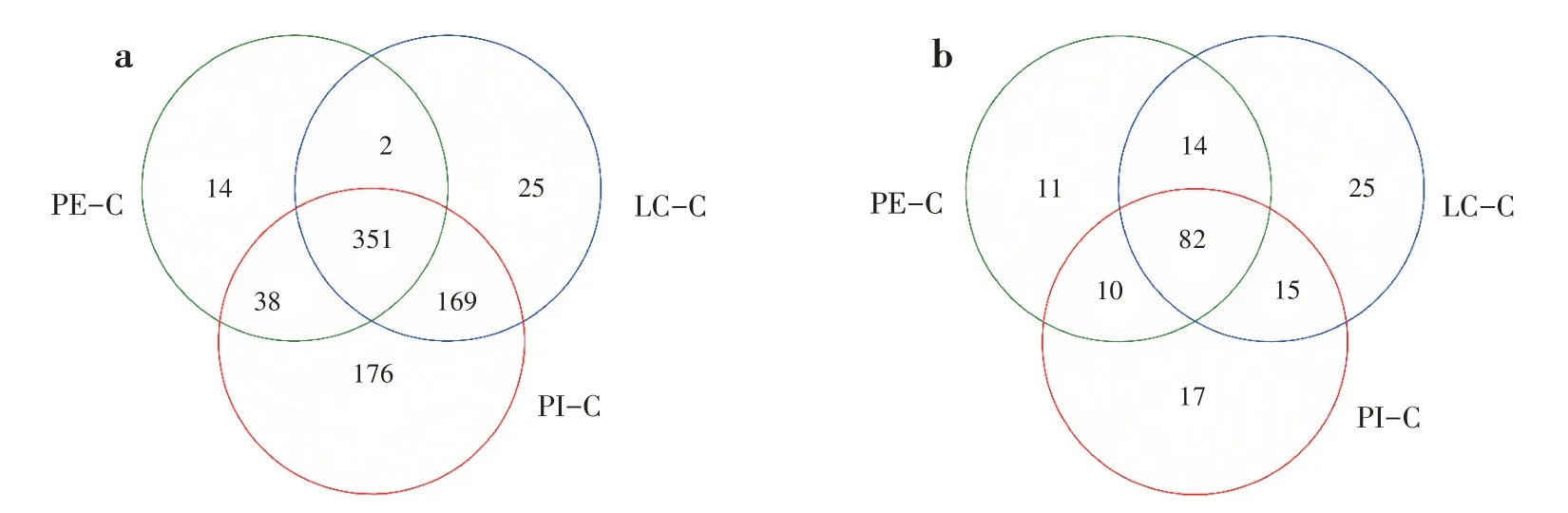
圖3 不同套種模式土壤微生物屬分類水平上Venn圖(a:細菌;b:真菌)Fig.3 Venn diagram of soil microbe of different interplanting models at the genus level(a:bacteria;b:fungi)
2.3.3 不同套種模式土壤微生物與環境因子關聯分析
冗余分析(RDA)是環境因子約束化的PCA 分析,反映樣本分布和環境因子間的關系[26]。細菌群落多樣性與pH、有機質呈顯著正相關(P<0.05),與全P 含量呈極顯著正相關關系(P<0.01),說明pH、有機質和全P 含量共同影響土壤細菌多樣性(圖4a);真菌群落多樣性與毛管持水量呈顯著正相關(P< 0.05),說明毛管持水量影響土壤真菌多樣性(圖4b)。
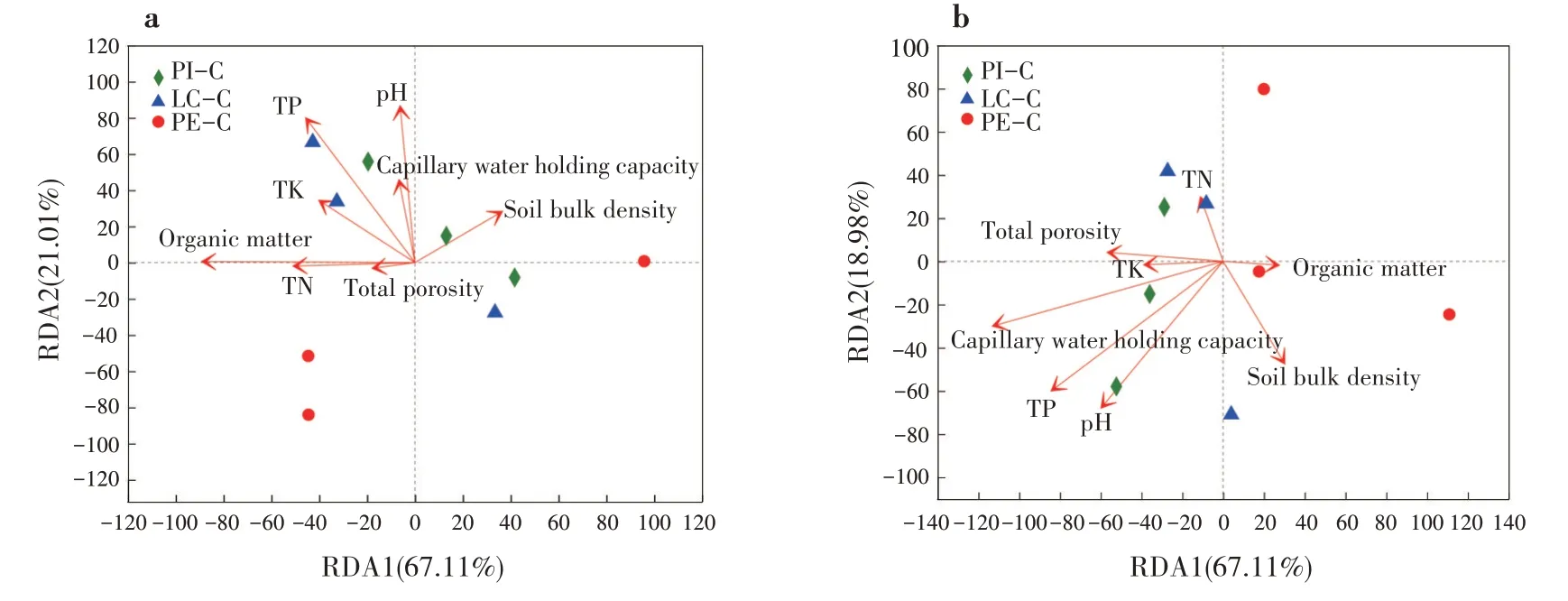
圖4 不同套種模式土壤微生物群落結構與環境因子的RDA分析(a:細菌;b:真菌)Fig.4 RDA analysis of soil microbial community structure and environmental factors of different interplanting models(a:bacteria;b:fungi)
通過計算環境因子和菌群之前的Spearman 等級相關系數發現,細菌的pH 與赭黃嗜鹽囊菌屬(Haliangium)、Gaiella、硝化螺旋菌屬(Nitrospira)、unclassified_o_Acidobacteriales、芽孢桿菌屬(Bacil?lus)、Luedemannella、土微菌屬(Pedomicrobium)呈現極顯著極強正相關(P<0.01;P<0.001;R ≥0.8);全P 含量與赭黃嗜鹽囊菌屬、Gaiella、硝化螺旋菌屬、unclassified_o_Acidobacteriales、芽孢桿菌屬、鏈霉菌屬(Streptomyces)呈極顯著極強正相關(P<0.01;P<0.001;R ≥0.8),與norank_p_WPS-2 呈極顯著極強負相關(P<0.01;R ≤-0.8);全K 含量與芽孢桿菌屬呈極顯著極強正相關(P<0.01;R ≥0.8);有機質與norank_p_GAL15 呈極顯著極強負相關(P<0.01;R ≤-0.8)(圖5a)。
真菌的容重與norank_f_Stenostomidae、有機質含量與Nuclearia均呈極顯著極強負相關(P<0.01;R ≥0.8);全鉀含量與Plectosphaerella和枝孢屬(Cladosporium)呈極顯著極強正相關(P<0.01;R ≥0.8),與乳菇屬(Lactarius)呈極顯著極強負相關(P<0.01;R ≥0.8);毛管持水量與unclassified_o_Peziza?les、unclassified_o_Eurotiales 呈極顯著極強正相關(P<0.01;R ≤-0.8),與Saitozyma、unclassified_p_As?comycota 和雙擔菌屬(Geminibasidium)呈極顯著極強負相關(P<0.01;R ≤-0.8);pH 與Arthrobotryss、全P含量與unclassified_o_Pezizales 均呈極顯著極強正相關(P<0.01;R ≤-0.8)(圖5b)。
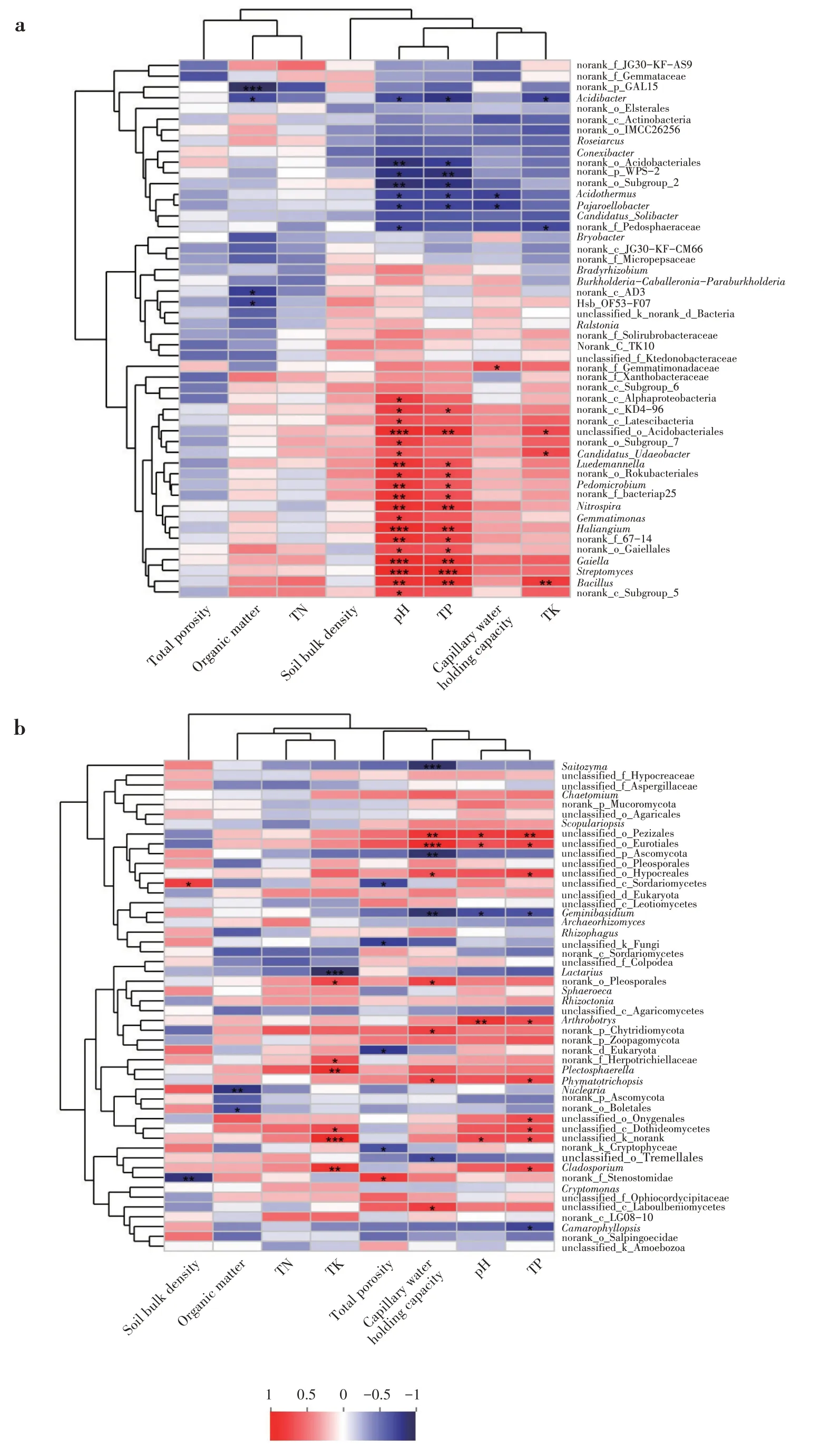
圖5 屬分類水平上土壤微生物相對豐度與環境因子相關性(a:細菌;b:真菌)Fig.5 Correlation between relative abundance of soil microbes and environmental factors at genus level(a:bacteria;b:fungi)
3 討論與結論
3.1 不同套種模式枯落物對土壤微生物多樣性的影響
不同植物的凋落物理化性質不同,分解時釋放的養分也具有差異性,對土壤微生物生長具有選擇性刺激作用,進而影響微生物群落結構和功能及其多樣性[27]。本研究表明印度紫檀×金花茶林、荔枝×金花茶林土壤細菌豐富度和多樣性明顯優于濕地松× 金花茶林,這與Klimek 等[28]研究結果相似,闊葉樹種較針葉樹種具有更高的土壤微生物活性和功能多樣性,亦與張信思[3]調查發現的野生金花茶原生生境的伴生植物多為闊葉林的特性相一致。
土壤細菌真菌比可反映細菌和真菌的相對豐富度[29]。在本研究中,印度紫檀×金花茶林的細菌真菌比明顯高于荔枝× 金花茶林和濕地松× 金花茶林,可能是因為印度紫檀×金花茶林枯落物的全N 和全P年變化量均明顯高于其他兩種林地,有研究表明枯落物中的N、P 元素釋放量越高,越有利于細菌的生長[30]。在后續研究中,還需進行枯落物礦化速率試驗進一步闡明不同金花茶套種模式的枯落物對微生物群落結構和多樣性的影響。
3.2 不同套種模式土壤理化性質對微生物多樣性的影響
土壤微生物對環境極為敏感,森林類型和土壤理化性質是影響其多樣性的重要因素[31-32]。RDA 結果表明,pH、有機質和全P 含量對土壤細菌多樣性的影響較大,其中,全P 含量的影響最大,呈極顯著相關;毛管持水量則影響土壤真菌多樣性。宋賢沖等[12]研究發現全P 含量與土壤微生物多樣性指數極顯著相關,有機質與土壤微生物多樣性表現為顯著相關;邵穎等[33]研究表明pH 值與土壤細菌群落多樣性有關,與本研究的發現一致。孔隙度、全N和全K 含量對土壤微生物多樣性貢獻不大,這與趙春梅等[34]的研究結果不一致,可能是收獲金花茶花朵和葉片時人為活動較多所致,且孔隙度、全N和全K并不能影響所有土壤微生物群落。根據相關性Heat?map 分析結果可知,pH、有機質、全P 和全K 含量可與部分細菌屬種類呈極顯著極強相關性,容重、毛管持水量、pH、全P含量和全K含量可與部分真菌屬呈極顯著極強相關性。
土壤生境不同,主要菌群的豐度和結構不同[35]。由于3 種套種模式土壤的孔隙度、pH、全P 和全K 含量均差異顯著,使土壤生境具有差異。3 種套種模式土壤中的優勢微生物均為變形菌門、放線菌門、酸桿菌門、綠彎菌門4個細菌菌群和子囊菌門、擔子菌門2 個真菌菌群,研究結果與張勝男等[35]和Han?son 等[36]的基本一致。有研究認為綠彎菌門和富營養化程度有關[37],酸桿菌門與貧瘠的土壤環境有關[38],子囊菌門能利用環境中多種養分供其生長[39],擔子菌門可作為土壤擾動強弱的指示真菌[40]。印度紫檀×金花茶林土壤微生物豐富度和多樣性最高,占比最多的是綠彎菌門和子囊菌門,特有的細菌屬種類也遠遠多于其他兩種模式的土壤,可能原因是印度紫檀×金花茶林枯落物代謝較快,更多養分流入土壤。荔枝× 金花茶林土壤中占比最多的是變形菌門和子囊菌門,濕地松×金花茶林土壤中占比最多的是放線菌門和擔子菌門。
本研究結果表明,枯落物蓄積量、全N 和全P年變化量以及土壤pH、有機質、全P 含量和毛管持水量能影響土壤微生物群落結構和多樣性。綜合考慮土壤理化性質和微生物多樣性,在林下套種金花茶栽培模式中,宜選擇落葉闊葉林作為上層林樹種。
