陸地棉轉錄因子GhAHL4 基因的克隆及表達分析
2021-05-24 01:52:44劉萌萌韓立軍劉寶玲薛金愛李潤植
山西農業科學 2021年5期
劉萌萌,韓立軍,劉寶玲,薛金愛,李潤植
(山西農業大學分子農業與生物能源研究所,山西太谷030801)
轉錄因子(Transcription factors)通過與真核生物基因啟動子區順式作用元件相互作用激活或者抑制相關基因的表達,從而調控植物生長發育。其本質上是一種蛋白質,由4 個部分組成,包括DNA結合區、轉錄調控區、核定位信號區以及寡聚化位點[1]。含有AT-hook 基序的AHL(AT-Hook Motif Nuclear Localized)轉錄因子最初是在哺乳動物非組蛋白的染色體高移動群蛋白HMG-I/Y 中發現的[2],現已在多種植物中的DNA 結合蛋白中鑒定到同源序列。植物AHL 蛋白家族成員含有1 個或2 個AT-hook 基序及PPC/DUF296 保守結構域,能改變染色質結構并調節基因表達[3]。其中,AT-hook 基序以精氨酸- 甘氨酸- 精氨酸- 脯氨酸(RGRP)為保守殘基[4],是一種新的DNA 結合蛋白基序與DNA發生特異性識別[5-6],而PPC 結構域在AHL 蛋白的細胞核定位中發揮重要作用[7]。AHL 蛋白既可以彼此之間,也可以與其他轉錄因子相互作用共同調節植物新陳代謝[8]。
有研究發現,擬南芥和水稻2 個模式植物中AHL基因家族主要參與調控植物開花、根系活力及其生長發育以及下胚軸的伸長生長等形態建成過程,如擬南芥中超表達AHL27基因[9],可抑制開花基因FT的表達,同時促進FLC 的表達,進而延遲擬南芥在長日和短日條件下的開花時間;在種子萌發過程中,AtAHL4基因表達抑制編碼TAG 脂酶和β- 氧化相關基因SDP1、DALL5、KAT5的表達,從而調控植物體內TAG 的降解[10];AtAHL18通過調節根尖分生組織的活動、側根的起始和出苗來參與根系結構的調節[11];AtAHL27/29基因能夠抑制光照下幼苗的下胚軸伸長[12]。此外,AHL 在植物非生物脅迫反應中發揮著重要的作用。例如,水稻OsAHL1基因的過表達可顯著提高水稻的抗旱性[13];玉米全基因組鑒定表明,ZmAHLs主要調節植物的花粉發育、干旱響應、衰老和創傷響應過程[14]。含有AT-hook motif 基序的GhAT1 蛋白,能夠特異性結合棉花FSltp4基因的啟動子,抑制FSltp4基因的表達,從而使棉纖維的生長受到嚴重影響[15]。由此可見,不同植物的AHL 及其成員所行使的生物學功能不同,目前僅有少數AHL 蛋白被發現,這個家族的其他成員仍有待進一步研究。
棉花(Gossypium hirsutum)種植規模較大,棉籽油作為棉花生產的副產品,是植物油的來源之一,同時也是制備生物柴油的天然原料。然而,在農業生產中,環境脅迫通常會導致細胞損傷,從而使得棉花的生長和產量受到嚴重限制[16]。為了適應環境因素的變化,植物自身能夠形成一些機制抵御逆境脅迫。在逆境條件下,植物可感知非生物脅迫信號產生防御調控機制以應對外界不良環境。
為進一步解析棉花抵御非生物脅迫的AHL 表達模式,本研究以冀豐1271 棉花品種為材料,克隆陸地棉GhAHL4基因,采用生物信息學工具分析其編碼蛋白理化性質,用RT-qPCR 分析該基因在鹽脅迫處理下的表達模式,旨在為深入解析棉花GhAHL4基因的生物學功能提供科學參考,并為培育抗非生物脅迫能力強的棉花新品種提供理論依據。
1 材料和方法
1.1 試驗材料
陸地棉冀豐1271 種子、大腸桿菌DH5α 菌株、pMD18-T 載體均由山西農業大學分子農業與生物能源研究所提供。供試陸地棉材料于2020 年春種植于山西農業大學農作站試驗田。
1.2 陸地棉AHL4 基因克隆
根據已報道的擬南芥AtAHL4基因的蛋白序列,利用BlastP 搜索棉花基因組數據庫CottonFGD(https://cottonfgd.org/),篩選棉花AHL4候選同源基因,結合轉錄組和各種脅迫處理表達譜數據,最終鑒定獲得在非生物脅迫中上調表達的GhAHL4基因,并下載其基因序列。
使用Plant Total RNA Kit 提取棉花葉片的總RNA,使用ABM 公司的反轉錄試劑盒5×All-In-One MasterMix(with AccuRT Genomic DNA Removal Kit)反轉錄成cDNA,克隆陸地棉GhAHL4基因。以棉花葉片cDNA 為模板,采用天根公司的高保真酶Pfu DNA Polymerase 進行PCR 反應。20 μL 反應體系:10×Pfu Buffer 2 μL、dNTP Mix(2.5 mmol/L)1.6 μL、Pfu DNAPolymerase0.5 μL、上下游引物(表1)各0.5 μL、cDNA 模板2 μL、RNase-free H2O 12.9 μL。PCR 反應程序為:94 ℃預變性5 min;94 ℃變性30 s,58 ℃退火30 s,72 ℃延伸30 s,30 個循環;72 ℃終延伸10 min。PCR 產物經1%瓊脂糖凝膠電泳后,割膠回收目的條帶,然后連接到pMD18-T 載體上,轉化至大腸桿菌DH5α 感受態細胞,篩選陽性克隆后送上海生工有限公司測序。
表1 引物序列
1.3 陸地棉GhAHL4 基因的生物信息學分析
經ProParam(https://web.expasy.org/protparam/)分析陸地棉GhAHL4 編碼氨基酸的數目、蛋白的分子質量以及理論等電點、脂肪系數及穩定性等理化性質;利用TMHMM 2.0(http://www.cbs.dtu.dk/services/TMHMM/)和SignalP 3.0(http://www.cbs.dtu.dk/services/SignalP-3.0/)對氨基酸序列進行跨膜及信號肽分析;用在線分析工具LocTree 3(https://rostlab.org/services/loctree3/) 預測亞細胞定位。采用SPOMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=/NPSA/npsa_sopma.html)在線分析軟件預測蛋白質二級結構;通過SWISS-MODEL(https://swissmodel.expasy.org/interactive)對GhAHL4 蛋白的三級結構進行同源建模分析;運用在線分析軟件Plant CARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)對轉錄起始位點上游長度為2 000 bp 啟動子的序列進行分析,并對順式作用元件在GSDS 2.0 進行可視化體現;利用基因結構顯示系統GSDS 分析GhAHL4 基因外顯子、內含子數量,繪制基因結構圖;用ClustalX 和GeneDOC 軟件進行多序列比對,并分析其保守結構域;基于鄰接法(neighbor-joining,NJ)采用MEGA 7.0 構建無根系統發育樹。
1.4 陸地棉GhAHL4 基因表達分析
利用CottonFGD(https://cottonfgd.org/)下載陸地棉GhAHL4 的轉錄組數據,通過構建Heatmap 圖綜合分析該基因的表達情況。
冀豐1271 號種子脫絨后直接播種于滅菌蛭石中,待幼苗長至2 片真葉時,選取長勢一致的幼苗,將根部的蛭石經流水沖洗干凈,快速置于吸水紙上吸干,將棉苗的根部浸入200 mmol/L NaCl 溶液中,置于(25±2)℃培養箱,連續光照條件下生長,分別于0、1、3、6、12、24、48、72 h 采集真葉葉片,提取總RNA,反轉錄合成cDNA,以棉花histone 基因作為內參,使用TaKaRa 公司的TB GreenTMPremix Ex TaqTMⅡ熒光定量PCR 試劑盒進行RT-qPCR。反應總體系為10 μL:TB Green Premix Ex Taq II 5 μL、上下游引物(表1)各0.4 μL、cDNA 模板0.8 μL、RNasefree H2O 3.4 μL。反應程序為:95 ℃5 min;95 ℃5 s,58 ℃30 s,72 ℃30 s,40 個循環。用2-ΔΔCt法進行基因的相對表達量分析[17]。每個樣品設置3 個生物學重復。
2 結果與分析
2.1 陸地棉GhAHL4 基因的克隆及其編碼蛋白的理化性質分析
根據已報道的擬南芥AtAHL4基因的蛋白序列,利用BlastP 搜索棉花基因組數據庫CottonFGD,篩選棉花AHL4同源基因,結果發現,該基因在A亞組、D 亞組均有拷貝,結合轉錄組和各種脅迫處理表達譜數據,發現只有D 亞組GhAHL4差異表達顯著,最終鑒定獲得在非生物脅迫中上調表達的GhAHL4基因。以冀豐1271 陸地棉葉片的cDNA為模板,克隆到GhAHL4基因cDNA 全長為984 bp(圖1),與基因組所注釋的該基因編碼序列存在部分堿基差異,但蛋白序列一致。
理化性質分析表明,GhAHL4基因編碼327 個氨基酸,理論等電點為9.44,分子質量約33.74 ku,不穩定系數為50.1,脂肪系數為68.56,親水性系數為-0.353,據此推測,GhAHL4 蛋白為不穩定親水性蛋白質[18-19]。亞細胞定位預測結果顯示,其定位于細胞核中,預測可信度為84,無信號肽和跨膜結構,屬于一種非分泌型蛋白。
2.2 陸地棉GhAHL4 基因的結構分析
采用SOPMA 在線預測GhAHL4 的二級結構,結果顯示(圖2),GhAHL4 蛋白結構以無規則卷曲(Random coil)和延伸鏈(Extended strand)為主,分別占68.20%和23.85%。通過在線軟件SWISS-MODEL對GhAHL4 蛋白的三維結構進行預測分析與同源建模,以2dt4.1.A[8]蛋白(a plant-and prokaryote-conserved(PPC)protein inPyrococcus horikoshii)為模板,該蛋白與棉花GhAHL4 蛋白氨基酸序列相似性為29%,包含3 個GOL 配體(圖3)。
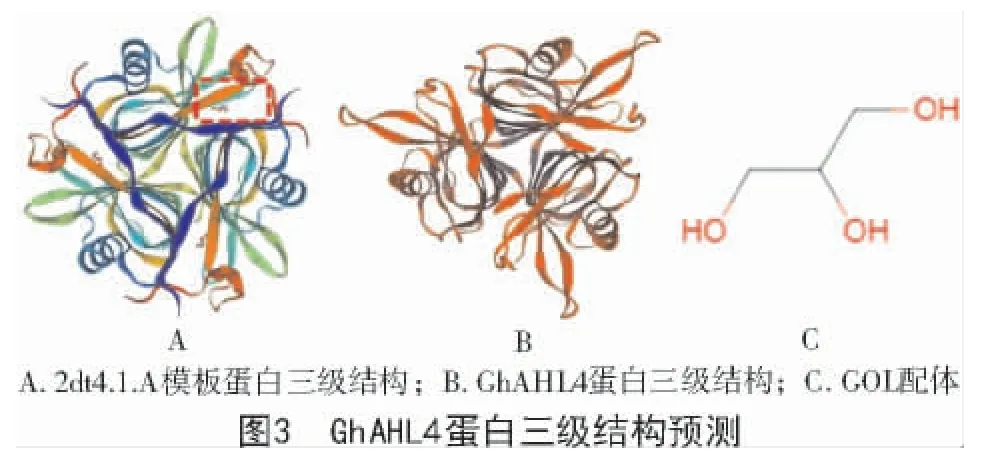
利用GSDS 2.0 在線檢測GhAHL4基因結構發現,該基因含有5 個外顯子、4 個內含子(圖4)。
為探究該基因是否可能受逆境脅迫調控,進一步分析GhAHL4基因上游啟動子序列,結果顯示(圖5),GhAHL4基因啟動子含有多個參與植物響應非生物脅迫的順式作用元件,響應脫落酸脅迫的ABRE 元件、響應光反應的G-box 及低溫響應元件LTR。

2.3 陸地棉GhAHL4 蛋白的序列比對和進化分析


多序列比對顯示,GhAHL4 蛋白與木槿HsAHL4(KAE8694315.1)、榴蓮DzAHL4(XP_022719610.1)、蓖麻RcAHL4(XP_002509630.1)、大豆GmAHL4(XP_003525779.1) 的同源蛋白的序列相似性分別為86.54%、84.89%、75.45%和73.09%(圖6)。為了明確陸地棉GhAHL4 與其他植物AHL 蛋白的系統發育的關系,利用MEGA 7.0 軟件對GhAHL4 和其他植物AHL 蛋白進行序列比對,并構建進化樹,進化分析結果表明(圖7),GhAHL4 除與亞洲棉GaAHL蛋白外,與木槿HsAHL4(KAE8694315.1)的親緣關系最近。
2.4 GhAHL4 在不同組織和非生物脅迫下的表達模式分析
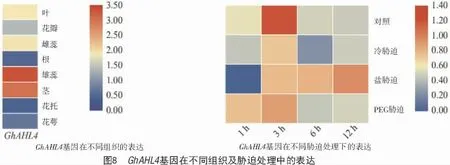
陸地棉GhAHL4在不同組織及不同脅迫中的表達情況如圖8 所示,該基因在根中表達量最低,在雄蕊中高表達;冷脅迫及干旱脅迫處理3 h 時GhAHL4基因表達量最高,之后隨著脅迫時間的延長表達量呈下降趨勢,而在鹽脅迫條件下該基因的表達量在12 h 內持續升高。
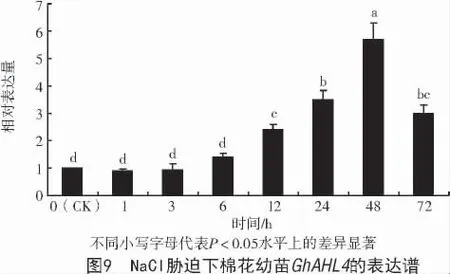
為進一步分析GhAHL4基因在鹽脅迫下的表達模式,用200 mmol/L NaCl 脅迫處理2 片真葉的棉花幼苗,探究脅迫處理0、1、3、6、12、24、48、72 h時的GhAHL4差異表達,結果顯示,處理0~3 h 棉花GhAHL4基因的表達量沒有發生顯著變化;在處理6~48 h 時間內,其表達量呈上升趨勢,48 h 時表達量最高,約為CK 的6 倍,且二者間差異達顯著水平(P<0.05);處理48 h 后,表達量開始下調(圖9)。表明棉花GhAHL4基因參與棉花幼苗對鹽脅迫的應答。
3 結論與討論
AHL 蛋白作為一類DNA 結合蛋白,在植物的生長發育過程中起著重要的調控作用。目前,在花生[20]、番茄[21]以及模式植物擬南芥[22-23]和水稻[24]中有關AHL 蛋白的研究較為深入。棉花作為我國重要的經濟作物之一,有關其AHL蛋白功能研究鮮見報道。本研究克隆了陸地棉GhAHL4基因,含有5 個外顯子和4 個內含子,而大豆GmAHL4基因卻不含內含子[25],由此可見,不同AHL基因的結構存在很大的差異。編碼蛋白定位于細胞核內,這與前人研究AHL 蛋白大多定位于細胞核的結果基本一致[15],且包含3 個GOL 配體(三羥基糖醇,作為碳水化合物和脂質代謝的中間產物),可能參與脂肪酸生物合成。多序列比對結果顯示,陸地棉GhAHL4蛋白存在AT-hook 結構域的保守殘基KKKRGRP,這與擬南芥AtAHLs 蛋白結構一致[4],可能通過與下游靶基因的DNA 結合從而參與其表達調控。系統發育分析表明,GhAHL4 蛋白與同科的亞洲棉及木槿聚為一支,因為陸地棉與亞洲棉及木槿同為錦葵科植物,這也與經典植物分類的結論一致。
GhAHL4基因上游啟動子包含了與光響應有關的元件G-box、赤霉素應答元件GAREAT、脫落酸應答相關元件ABRE 等,以及低溫響應元件LTR,這與毛果楊PtrAHL6、PtrAHL18、PtrAHL24啟動子中含有脫落酸響應及低溫響應等順式作用元件結構相似[26]。不同脅迫下基因表達模式進一步表明,GhAHL4可能參與逆境環境脅迫應答。RT-qPCR結果顯示,NaCl 脅迫下GhAHL4基因表達量上調,這與丁麗雪等[21]在番茄中發現的部分AHL基因受鹽脅迫誘導結果相似,而OsAHL1的過表達增強了對鹽脅迫的耐受性[13]。表達量在48 h 內持續升高預示著該基因參與棉花幼苗鹽脅迫應答,基因過表達可能增強了棉花對逆境的適應性,因此可作為棉花抗逆機制研究和棉花抗逆遺傳改良的候選目的基因。但GhAHL4基因具體如何介導棉花抵御鹽脅迫等逆境應答的調控網路還有待進一步論證。
本研究以擬南芥AtAHL4 蛋白序列為參考序列,鑒定得到了棉花GhAHL4基因,通過生物信息學技術分析了基因結構和編碼蛋白結構及理化特性,通過RT-qPCR 基因表達分析發現,GhAHL4基因的表達受鹽脅迫的誘導,結果為進一步闡明棉花GhAHL4基因的生物學功能以及基因工程育種研究奠定了基礎。
猜你喜歡
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
山東工業技術(2016年15期)2016-12-01 05:31:22
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
