豬偽狂犬病病毒山西株的分離鑒定及遺傳變異分析
2021-05-24 01:53:10牛志紅任建樂王喜珍張衛(wèi)兵趙宇軍寧官保田文霞
山西農業(yè)科學 2021年5期
關鍵詞:檢測
牛志紅,任建樂,王喜珍,張衛(wèi)兵,張 鼎,趙宇軍,寧官保,田文霞
(山西農業(yè)大學動物醫(yī)學學院,山西太谷030801)
偽狂犬病(Pseudorabies,PR)是由偽狂犬病病毒(Pseudorabies virus,PRV)引起的多種家畜及野生動物共患的以奇癢、化膿性腦炎等神經(jīng)癥狀為主的急性接觸性傳染病[1]。PRV 為皰疹病毒科、α- 皰疹病毒亞科、水痘病毒屬的病毒成員[2-3]。該病毒可引起母豬流產(chǎn)、產(chǎn)木乃伊胎、仔豬死亡等臨床癥狀,給養(yǎng)豬業(yè)造成了嚴重的經(jīng)濟損失[4]。
自20 世紀70 年代末,我國引進匈牙利Bartha-K61 疫苗,并開始在豬場進行免疫,利用通過使用標記疫苗和各自的配套診斷性檢測區(qū)分感染與疫苗接種動物DIVA 策略,使我國的豬偽狂犬病疫情逐步得到控制[5-6],但從2011 年開始多個免疫Bartha-K61 疫苗的豬群相繼暴發(fā)由PRV 變異株引起的PR,隨后從河南、河北、山東等地蔓延至全國[3],并且PRV 變異株可引起人的呼吸功能障礙和腦炎等癥狀,成為潛在的人畜共患病原[7]。有研究表明,PRV 變異株基因組呈現(xiàn)較大的變異,對不同日齡的豬均有較強的致病性,其中,gE的變異在一定程度上可導致PRV 變異株致病力增強[8];變異株gC的變異是引起B(yǎng)artha-K61 血清中和能力降低的關鍵因子,并且gB、gC和gD變異均可導致Bartha-K61為宿主提供免疫保護能力下降[9-10]。另外,自2019 年以來多份最新研究發(fā)現(xiàn),PRV 發(fā)生了新的重組變異,PRV 變異株可與Bartha-K61 的gB和gC基因產(chǎn)生重組,形成新的重組變異株,可致病毒的致病性發(fā)生相應的改變,并且新的重組變異將可能加速PRV 在不同物種間的傳播[11-13]。
為調查山西省豬偽狂犬病病毒流行及其變異情況,本研究對2019 年采集的山西省部分規(guī)模豬場的4 346 份豬血清及964 份豬疑似病料進行血清學gB及gE抗體及病原學檢測分析,并對PRV分離株的gC和gE基因進行遺傳變異分析,以期為山西省PRV 的防控和凈化提供相關理論依據(jù)。
1 材料和方法
1.1 材料
非洲綠猴腎細胞(Vero-E6)由中國科學院微生物研究所饋贈;SX1801 和SX802 毒株均為山西農業(yè)大學生物制品實驗室保存。
1.2 試劑與儀器
gB-ELISA 檢測試劑盒、gE-ELISA 檢測試劑盒,均購自美國愛德士(IDEXX)公司;病毒DNA/RNA提取試劑盒,購自天根生化有限科技公司;豬偽狂犬病病毒實時熒光PCR 試劑盒,購自湖南國測生物科技有限公司;DMEM、FBS,購自賽默飛世爾科技有限公司;FITC 標記的羊抗鼠IgG、胰蛋白酶-EDTA 消化液、青霉素- 鏈霉素- 兩性霉素混合液、去離子水,均購自索萊寶科技有限公司;PreMix Taq 酶,購自大連寶日生物技術有限公司。
QuantStudio6 實時熒光定量PCR 儀,購自美國ABI 公司;徠卡倒置顯微鏡,購自德國徠卡公司;PCR 擴增儀,購自德國Eppendorf 公司;DYY-8C 型電泳儀,購自北京市六一儀器廠;SYNGENE 全自動凝膠成像分析儀,購自美國SYNGENE 公司。
1.3 方法
1.3.1 樣本的采集與處理 2019 年1—12 月在山西省6 個gE基因缺失苗免疫豬場,采集不同生長階段的健康豬新鮮血清,共4 346 份,置于-20 ℃?zhèn)溆谩2杉i場疑似病豬的全血、臍帶、淋巴結、腦及胎兒胎衣等組織,共964 份,一部分加入液氮碾磨成組織粉末用于qPCR 檢測,剩余部分置于-80 ℃保存。
1.3.2 血清及病原檢測 利用PRVgE-ELISA、PRVgB-ELISA 抗體試劑盒對采集的血清樣本進行gE、gB抗體檢測,具體操作和判定根據(jù)說明書進行。采用病毒DNA/RNA 提取試劑盒對組織樣品進行PRV 基因組提取,然后使用豬偽狂犬病毒實時熒光PCR 試劑盒進行qPCR,具體操作及結果判定根據(jù)說明書進行。
1.3.3 引物的設計與合成 參考Ea 株(GenBank登錄號KU315430.1)基因序列,利用Primer 5.0 軟件設計特異性并合成針對gC和gE的特異性引物(表1)。引物由武漢天一輝遠生物科技有限公司合成。

表1 PRV 引物序列
1.3.4 病毒基因組的提取及PCR 鑒定 取50 mg陽性病料反復凍融3 次,置于組織碾磨器中碾成勻漿液,取200 μL 勻漿液按照病毒基因組DNA/RNA提取試劑盒說明書提取DNA。利用gE檢測引物進行PCR 擴增,1%瓊脂糖凝膠電泳檢測PCR 產(chǎn)物,陽性PCR 產(chǎn)物送至武漢天一輝遠生物科技有限公司進行測序鑒定。
1.3.5 病毒分離培養(yǎng) 將PCR 鑒定為陽性的組織勻漿液離心取上清,0.22 μL 濾器過濾除菌后,取500 μL 接種于單層Vero 細胞,同時設置正常細胞對照,置于37 ℃、5%CO2培養(yǎng)箱吸附1 h 后,棄上清加入含2%胎牛血清的DMEM細胞維持液,每日觀察細胞,待細胞出現(xiàn)明顯的細胞病變效應(CPE),且細胞病變達到80%時,收取病液,若無病變則繼續(xù)盲傳,持續(xù)盲傳3 代無病變則視為陰性。
1.3.6 病毒IFA 鑒定 將病毒分離株接種單層Vero 細胞,同時設置陽性對照(SX1801 與SX1802)及正常細胞對照,感染36 h 出現(xiàn)明顯的CPE 時,棄上清,PBS 洗滌3 次,以預冷的無水乙醇4 ℃固定30 min,棄上清,PBS 洗滌3 次,以抗PRV 的gB鼠源單克隆抗體(1∶2 000)為一抗,37 ℃孵育1 h,PBS 洗滌3 次;以FITC 標記的羊抗鼠IgG(1∶200)為二抗,37 ℃孵育1 h,PBS 洗滌3 次,于倒置熒光顯微鏡下觀察結果。
1.3.7 PRV 分離株gC、gE基因遺傳變異分析 提取病毒DNA,利用表1 中的gC、gE基因克隆引物進行PCR 擴增,將PCR 產(chǎn)物送至武漢天一輝遠生物科技有限公司進行測序,利用生物學分析軟件DNAStar 7.1、MEGA 5.0 將分離株與近年來山西農業(yè)大學動物醫(yī)學學院生物制品實驗室分離鑒定(SX1801 與SX1802) 的及GenBank 中已登錄的PRV 參考毒株gC和gE核苷酸和氨基酸序列進行比對,分析分離株的序列特征。
2 結果與分析
2.1 血清學及病原學檢測

表2 血清學檢測結果
將2019 年收集的山西省6 個不同豬場免疫豬群的血清,進行偽狂犬gE、gB抗體檢測。檢測結果表明(表2),臨汾市A 豬場、朔州市A 豬場、朔州市B豬場、汾陽市A 豬場、呂梁市A 豬場、呂梁市B 豬場gE陽性率分別為0、0、18.2%、6.3%、3.5%、39.5%;gB陽性率分別為100%、95.6%、98.5%、95.5%、98.5%、97.6%。此次調查共檢測血清樣品4 346 份,其中,PRVgE抗體陽性率為11.3%,gB陽性率平均為97.6%,有4 個豬場為PRVgE抗體陽性豬場。
2019 年隨機收集山西省4 個不同豬場的部分免疫豬群的全血、胎兒胎衣、臍帶、淋巴結、腦等器官組織,共964 份樣本,經(jīng)qPCR 擴增檢測顯示(表3),僅朔州市B 豬場發(fā)現(xiàn)2 例PRV 感染病例,豬場病原檢出率為1.79%。其中,汾陽市A 豬場、呂梁市A 豬場、呂梁市B 豬場陽性病例個數(shù)均為0。綜合血清學和病原學結果顯示,盡管個別豬場中的gE抗體陽性率較高,但豬場的病原檢出率較低,總體病原檢出率僅為0.21%。
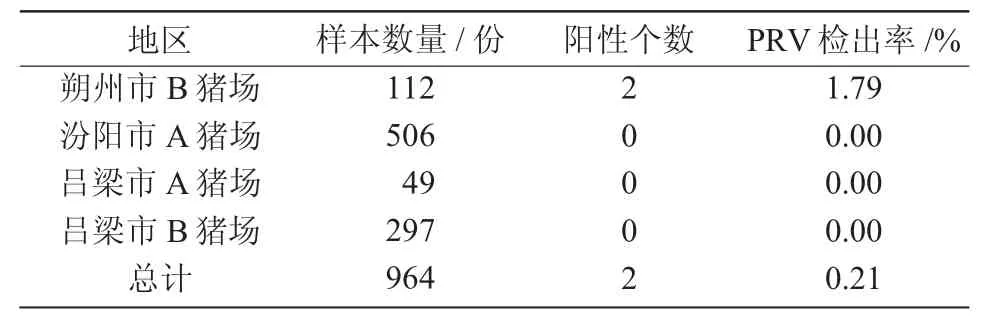
表3 qPCR 檢測結果
2.2 組織病料PCR 擴增
將qPCR 檢測為PRV 陽性的2 份組織病料研磨后,提取病毒基因組,利用gE檢測引物進行PCR擴增。由圖1 可知,PCR 擴增產(chǎn)物經(jīng)瓊脂糖凝膠電泳后條帶單一,大小為635 bp,與預期結果一致。

2.3 PRV 的分離及IFA 鑒定
將2 份PRV 陽性組織的勻漿液過濾除菌后,分別接種于單層Vero 細胞,48 h 后,其中1 份陽性樣品可致Vero 細胞產(chǎn)生固縮、變圓和脫落等疑似PRV的CPE 現(xiàn)象,并將該分離毒株命名為SX1911。以PRVgB單克隆抗體為一抗,F(xiàn)ITC 標記的羊抗鼠IgG 為二抗,對分離病毒株進行IFA 鑒定,結果顯示(圖2),分離株SX1911 與陽性對照組(2018 年分離株SX1801與SX1802)IFA 檢測均出現(xiàn)特異性綠色熒光。

2.4 PRV 分離株gC 基因遺傳變異分析
PCR 擴增分離株SX1911gE、gC全基因測序和拼接后分別得到1 740、1 464 bp 的完整的開放閱讀框。通過3 株山西PRV 分離株(SX1911、SX1801 和SX1802)的gC全長基因核苷酸及氨基酸序列與GenBank 登錄的17 株PRV 參考株同源性分析顯示(表4),3 株PRV 分離株與國外毒株(Kaplan、Becker 等毒株) 核苷酸和氨基酸相似性分別為94.9%~96.3%和90.4%~93.9%;與國內經(jīng)典株(SC、Ea 等)核苷酸與氨基酸相似性分別為96.8%~99.7%和94.3%~99.2%;與國內2011 年以后分離的毒株核苷酸與氨基酸相似性均為99.8%~100%;分離株間的核苷酸與氨基酸相似性分別為99.9%~100%和99.8%~100%。

表4 核苷酸和氨基酸的相似性分析 %
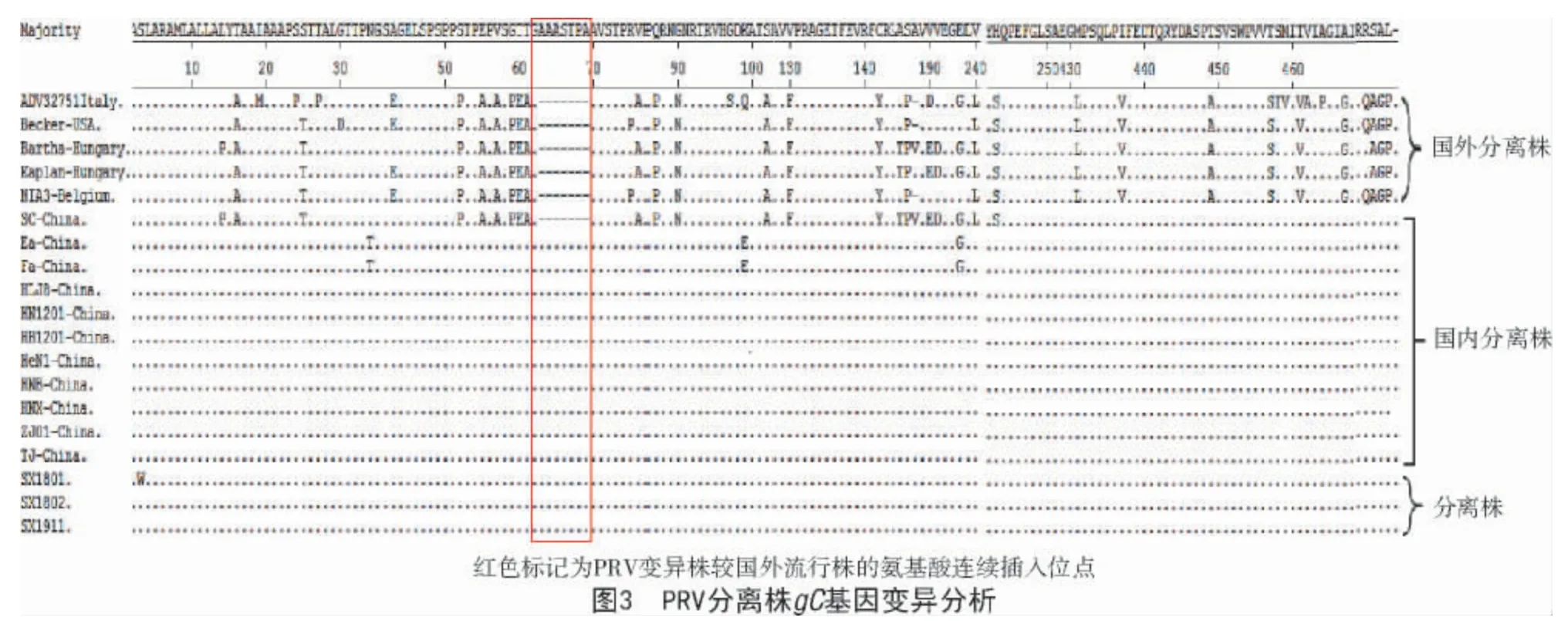
針對3 株PRV 變異株gC氨基酸序列進行分析表明,相較于國外流行株(Kaplan、Bartha 等),絕大部分PRV 變異株存在多個變異位點:在63—69 位氨基酸連續(xù)插入AASTPA7 個氨基酸,在43位點E→A(除Bartha 株外),在431 位點L→M,在130437 位點F→V,在449 位點V→A,在457 位點S→T,在461 位點V→T,在467 位點G→A,在142 位點Y→C,在190、191 位點均發(fā)生2 個氨基酸位點變異ED→VV(除Becker 和ADV32711Italy 株外),在243 位點S→H,在485—487 位點AGP→SAL。3 株山西PRV 分離株除SX1801 在3 位點發(fā)生一個氨基酸位點突變S→W 外,其他氨基酸變異位點均與國內早期分離的變異株相同(圖3)。
基于gC基因進行遺傳演化分析結果表明(圖4),國外毒株Becker、NAI3 等毒株屬于基因Ⅰ型,國內PRV 毒株為基因Ⅱ型。基因Ⅱ型病毒形成2 個獨立的亞分支,其中,國內變異株為一個單獨的亞分支,而國內經(jīng)典毒株分別形成不同獨立的亞分支。經(jīng)典毒株Fa、Ea 等毒株與變異株親緣關系近,而SC 株與變異株親緣關系較遠。本試驗研究的3 株PRV分離株與2011 年以后分離的PRV 國內變異株位于基因Ⅱ型的同一亞分支,且高度同源。

2.5 PRV 分離株gE 基因遺傳變異分析
針對SX1911、SX1801 和SX1802 株的gE全長基因核苷酸及氨基酸序列與GenBank 登錄的16 株PRV 參考株進行了同源性分析。結果顯示(表4),3 株山西PRV 分離株與國外毒株(Becker、Kaplan等毒株)核苷酸和氨基酸同源性分別為97.9%~98.2%和94.5%~95.0%;與國內經(jīng)典株(SC、Ea 等)核苷酸同源性均為99.7%,氨基酸同源性為98.4%~98.6%;與國內2011 年以后分離的毒株核苷酸與氨基酸同源性分別為99.8%~100%和98.4%~99.8%;分離株之間的核苷酸與氨基酸同源性分別為99.9%~100%和99.7%~99.8%。
基于gE蛋白氨基酸序列比對顯示,與國外毒株相比,國內PRV 分離株gE基因均存在17 個氨基酸突變位點。除此之外,與經(jīng)典株相比,國內PRV 變異株在55 氨基酸位點均存在突變位點D→G,在497 氨基酸位點插入一個天冬氨酸。與國內外經(jīng)典株相比,變異株(除TJ 外)均在449 氨基酸位點發(fā)生突變:V→I,與此同時,大多數(shù)變異株(除HLJB、TJ、SX1911 外)在511 氨基酸位點也發(fā)生突變:G→S(圖5)。

gE基因遺傳演化與gC相一致,如圖6所示,國內PRV 毒株與國外毒株處于2 個不同的分支,國外PRV 毒株屬于基因Ⅰ型,國內PRV 毒株屬于基因Ⅱ型。國內分離株具有共同的起源,但Ea、Fa、SC 等經(jīng)典株毒株處于基因Ⅱ型的一個亞分支,2011 年以后的PRV 分離株位于基因Ⅱ型的另一個亞分支,說明國內流行的變異株親緣性較近。本試驗研究的3 株PRV 分離株與國內流行株HB1201等分離株處于同一亞分支,同源性較高。

3 結論與討論
自2011 年以來,PRV 變異株給我國的養(yǎng)豬業(yè)造成了巨大的經(jīng)濟損失,由于PRV 變異株的致病機制尚未完全明確,并且疫苗Bartha-K61 不能提供完全的免疫保護,但目前通過DIVA 策略仍是防控和凈化PRV 感染的重要措施。因此,本研究通過對山西省部分地區(qū)PRV 流行及變異情況分析發(fā)現(xiàn),山西部分規(guī)模豬場仍存在PRV 野毒感染,并且呈現(xiàn)不同程度的gE抗體陽性;成功分離到1 株PRV 變異株,命名為SX1911;遺傳變異分析表明,分離株SX1911 與實驗室已保存毒株均屬于變異毒株。該研究可為山西省的PRV 流行情況及疫病防控提供相關科學依據(jù)。
豬偽狂犬病在山西省部分地區(qū)仍廣泛流行。2019 年對山西省部分地區(qū)的免疫豬場所采血清樣品進行了血清學檢測。結果表明,PRV 感染豬場的陽性率高達66.67%,大部分豬場血清樣品gE抗體陽性率在3.5%~39.5%,但仍有部分gE抗體陰性豬場存在,推測規(guī)模化豬場之間的防控措施以及生產(chǎn)管理水平的不同可能是影響PRV 感染率差異的原因之一。寧慧波等[14]通過對2019 年山西地區(qū)豬偽狂犬病的gE抗體調查結果顯示,感染豬場陽性率達71.43%,血清樣品gE陽性率達到52.16%,略高于河北和內蒙古,但低于北京。與此同時,陳馳等[15]研究發(fā)現(xiàn),2018—2019 年江蘇、河南和江西3 個省豬場gE陽性率達到71.7%~88.24%,且樣品gE抗體陽性率分別為38.44%、63.28%和30.48%。由此表明,目前山西省部分地區(qū)豬場PRV 感染情況與全國水平大致相同,仍然廣泛流行。本研究在所采樣豬場中的血清學結果顯示,PRVgE抗體陽性率較高,但qPCR 僅檢出2 例陽性病例,其原因可能與組織樣本容量和種類較少,或病毒轉為潛伏感染有關。已有研究發(fā)現(xiàn),部分豬場雖然受到PRV 變異毒株的感染,但無明顯的臨床癥狀,表現(xiàn)為種豬群gE抗體轉陽或陽性率升高[16];同時PRV 野毒的檢出率跟采樣季節(jié)有關系,殷鑫歡等[17]研究發(fā)現(xiàn),PRV的檢出率在夏季(1.5%)和秋季(1.6%)明顯比春季(9.1%)和秋季(33.0%)低。這些數(shù)據(jù)表明,目前山西及國內其他省份仍然面臨著嚴峻的豬偽狂犬病防控壓力,因此,豬場應適度優(yōu)化免疫程序,降低豬偽狂犬病的臨床發(fā)病率,加強生物安全措施,防止病毒的傳入和傳播,逐步凈化豬偽狂犬病[18]。
山西PRV 流行毒株仍為變異株。本研究針對新分離毒株SX1911 與山西農業(yè)大學生物制品實驗室已保存的SX1801 和SX1802 的gC和gE基因遺傳變異分析發(fā)現(xiàn),3 個毒株均與目前國內流行的毒株一致,位于基因Ⅱ型的同一個亞分支,屬于變異株。進一步序列比對發(fā)現(xiàn),相比于歐美毒株,3 個毒株的gC在第63 位點存在1 個AAASTPA 連續(xù)7 個氨基酸插入,這個位點的插入均與目前國內流行的毒株相一致。PRV 糖蛋白gC是能夠誘導機體產(chǎn)生中和抗體的重要免疫原,并且已有研究證明,gC變異是引起抗Bartha-K61 血清中和變異株的關鍵因素[10];進一步分析發(fā)現(xiàn),gC的變異主要存在于N 端的13-243 aa,其N 端存在介導PRV 與靶細胞的吸附的肝素結合域(heparin-bindingdomains,HBDs)[19],因此,gC的變異可能影響PRV 與靶細胞的吸附能力。最新研究表明,目前多株已分離的PRV 毒株基因發(fā)現(xiàn)重組變異,其中,變異株可與疫苗株Bartha-K61 的gC基因發(fā)生重組[11],但是在本研究中并未發(fā)現(xiàn)重組,SX1911、SX1801 和SX1802 這3 株病毒的其他基因是否能夠產(chǎn)生重組變異,需要全基因測序進一步分析。gE是決定PRV 的毒力的因子之一,且gE變異可部分增強變異株的致病性。本研究發(fā)現(xiàn),與國外株相比,所有國內分離株均具有17 個氨基酸突變位點,與孫穎等[20]的研究結果一致;所有變異株gE在48 和497 氨基酸位點均插入一個天冬氨酸;趙鴻遠等[21]研究認為,該突變可作為PRV 變異株診斷的分子特征。除此之外,大部分變異株還有個別氨基酸位點突變,這些變異可能是PRV 逐步進化形成獨立亞分支的原因。
本次流行病學調查反映了山西省部分地區(qū)PRV 流行情況,結果表明,PRV 仍然在山西省廣泛流行,流行毒株為變異株;血清學結果顯示,疫苗免疫后可產(chǎn)生高水平抗體,但仍可被PRV 野毒感染,提示應從當前PRV 流行變異株中選取合適的毒株研發(fā)疫苗,同時優(yōu)化管理程序為凈化PRV 提供高效防控措施。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
中學生數(shù)理化·七年級數(shù)學人教版(2020年12期)2021-01-18 06:57:46
中學生數(shù)理化·七年級數(shù)學人教版(2020年12期)2021-01-18 06:57:46
中學生數(shù)理化·七年級數(shù)學人教版(2019年9期)2019-11-25 07:34:36
中學生數(shù)理化·七年級數(shù)學人教版(2019年9期)2019-11-25 07:34:34
中學生數(shù)理化·七年級數(shù)學人教版(2019年12期)2019-05-21 02:53:50
中學生數(shù)理化·七年級數(shù)學人教版(2019年12期)2019-05-21 02:53:48
