穿龍薯蕷種子離體再生體系構建及其組培苗解剖結構特征
2021-05-25 02:01:24黨裳霓高潤梅石曉東張雨晴李紅月
西北植物學報 2021年4期
黨裳霓,高潤梅,石曉東,張雨晴,李紅月
(山西農業大學 林學院,山西太谷 030801)
穿龍薯蕷(Dioscoreanipponica)屬薯蕷科(Dioscoreaceae)薯蕷屬(Dioscorea)多年生草質藤本,廣泛分布于中國東北、華北、西北(除新疆)及河南、山東、安徽、浙江、江西 (廬山)、陜西 (秦嶺以北)、四川等地[1]。穿龍薯蕷的根狀莖俗稱穿山龍,主要活性成分是皂苷類化合物,是提取藥用成分薯蕷皂甙的重要原料[2-3]。近年來隨著對藥源材料的需求持續增加,連年的過度采挖,導致該種的野生資源急劇減少。穿龍薯蕷產地不同,根狀莖的薯蕷皂苷元含量差異較大,品質參差不齊[4-5],因此傳統的繁殖方式存在許多不足。組織培養技術可滿足大規模育苗的需要以及加快優良品種的推廣[6]。穿龍薯蕷木質化程度較高,愈傷組織產生較困難,以營養器官(幼根、莖段、葉片)為外植體產生愈傷時褐化嚴重,出愈率不高,有些器官根本不產生愈傷組織[7]。根狀莖常年生于地下,表面積累大量外源菌,消毒困難,接種后期還多會出現內源菌污染,造成離體培養失敗;而以莖段與葉片為外植體誘導愈傷組織會存在不同程度褐化,嚴重影響了愈傷組織的生長。因此,本研究以穿龍薯蕷的種子為試材,分析消毒時間、培養基激素配比對愈傷組織、腋芽誘導和生根培養等的影響,以期初步建立穩定和高效的穿龍薯蕷離體再生體系。同時,本研究進行了實生苗和組培苗解剖結構的比較,以期探討穿龍薯蕷在不同培養環境下的形態可塑性,衡量該種對環境的適應能力[8-9],旨在為后期的煉苗移栽及工廠化育苗提供科學理論依據。
1 材料和方法
1.1 植物材料
穿龍薯蕷種子采集于關帝山龐泉溝自然保護區。2018年秋季采集穿龍薯蕷成熟種子,風干后冰箱4 ℃冷藏。2019年6月選取飽滿均勻的種子,蒸餾水浸泡3 d后轉入200 mg/L的赤霉素溶液中浸泡 24 h,然后用鑷子去掉邊緣的種翅,置于含飽和洗衣粉溶液的燒杯中,流水持續沖洗30 min,再用蒸餾水沖洗3遍。將供試種子材料,放入超凈工作臺進行消毒[10]。
1.2 外植體消毒
將作為外植體的種子經75%酒精表面消毒1 min后,以2% NaClO進行消毒處理,分別設置12、15、18和21 min的處理時間,期間不斷振蕩;消毒結束后,用無菌水沖洗3次后平鋪在不添加激素的MS培養基表面,25 ℃暗培養。比較不同時間處理的消毒效果,接種15 d后,統計種子的死亡率、污染率和萌芽率。其中外植體無污染、未褐變視之為成活。
1.3 外植體離體培養
試驗所用的培養基中均添加蔗糖30 g·L-1,瓊脂7 g·L-1,pH調至6.0,121 ℃高壓滅菌30 min。所有培養物均于(25±1)℃的恒溫培養箱中培養,光照強度1 500 Lx,光、暗時間各12 h。
1.3.1 愈傷組織誘導愈傷組織選用激素6-BA+NAA(2,4-D)組合來誘導,采用單因素區組試驗,6-BA濃度為1.0 mg·L-1,NAA、2,4-D各設置0.2、0.5、1.0 和2.0 mg·L-1等4個水平,共組成8個處理組合。每種處理培養基接種10瓶,每瓶接種3個外植體,接種30 d后統計愈傷組織誘導率,出愈率(%)=出愈外植體數/接種外植體數×100%。
1.3.2 不定芽誘導不定芽的誘導采用激素6-BA與NAA組合來進行,各設置3個水平,6-BA分別為1.0、2.0 和3.0 mg·L-1,NAA分別為0.2、1.0 和2.0 mg·L-1,采用雙因素完全區組試驗,共組成9個配比處理組合。每種處理培養基接種30瓶,進行差異顯著性分析。
1.3.3 生根培養苗高3 cm以上時,進行生根誘導以獲得完整再生植株,結合前人研究結果,探究生長素NAA對生根誘導的影響[6-7,11-12]。以1/2MS為基本培養基添加不同濃度的激素,進行生根培養。其中,NAA選取0.1、0.2、0.5和1.0 mg·L-1等4個濃度水平,6-BA濃度為0.5 mg·L-1,IBA濃度為0.5 mg·L-1,設置單一NAA處理(0.1、0.5 mg·L-1)及其與6-BA、IBA組合處理(0.5 mg·L-1NAA+ 0.5 mg·L-16-BA;1.0 mg·L-1NAA+0.5 mg·L-16-BA;0.2 mg·L-1NAA + 0.5 mg·L-1IBA)。每種處理培養基接種15瓶,每瓶接種2株,60 d后觀察不同處理的幼苗生長狀況及生根率、根數等情況。生根率(%)=生根苗數/接種苗數×100%。
1.4 實生苗和組培苗解剖結構觀察
供試材料為1/2MS+0.5 mg·L-1NAA生根培養基培養60 d的組培苗,選取頂端以下的第3片葉、莖和根狀莖作為試驗材料,同時以室外培養60 d的實生苗為對照,比較二者在結構上的差異。其中,葉片剪取中部含主脈0.5 cm×0.5 cm的方塊,放入50% FAA固定液中固定24 h;分別剪取纏繞莖與根狀莖長0.5 cm的小段,放入70% FAA固定液中固定24 h。固定材料采用常規石蠟切片,切片厚度12 μm,番紅-固綠對染,中性樹膠封片[13]。在NikonEclipse50i光學顯微鏡下觀察葉片、莖和根狀莖切片解剖結構并拍照,采用Image-ProPlu5.0測量統計。
1.5 數據分析
試驗數據整理和圖表繪制采用Excel軟件完成。采用SPSS軟件對試驗數據進行統計分析,并用最小顯著性差異法(Duncan)對其處理后的數據進行顯著性差異多重比較,顯著水平為0.05。
2 結果與分析
2.1 消毒時間對外植體萌發的影響
由表1可知,2% NaClO消毒時間對外植體種子萌發指標(死亡率、污染率和萌芽率)有顯著影響(P<0.05)。其中,隨著消毒時間的增加,外植體的污染率逐漸降低,消毒15~21 min 處理均顯著低于消毒12 min處理,而其余處理間無顯著差異;外植體的萌芽率隨消毒時間延長而先升后降,并在消毒15 min和18 min分別為63.33%和70.00%,顯著高于其余處理。同時,隨消毒時間延長,外植體死亡現象逐漸加重,成活率顯著下降,死亡率在消毒21 min時達43.33%,顯著高于其余處理,而其余處理間則無顯著差異。可見,采用2% NaClO作為種子消毒劑的消毒時間以15~18 min為宜,萌芽率高,污染率和死亡率低。
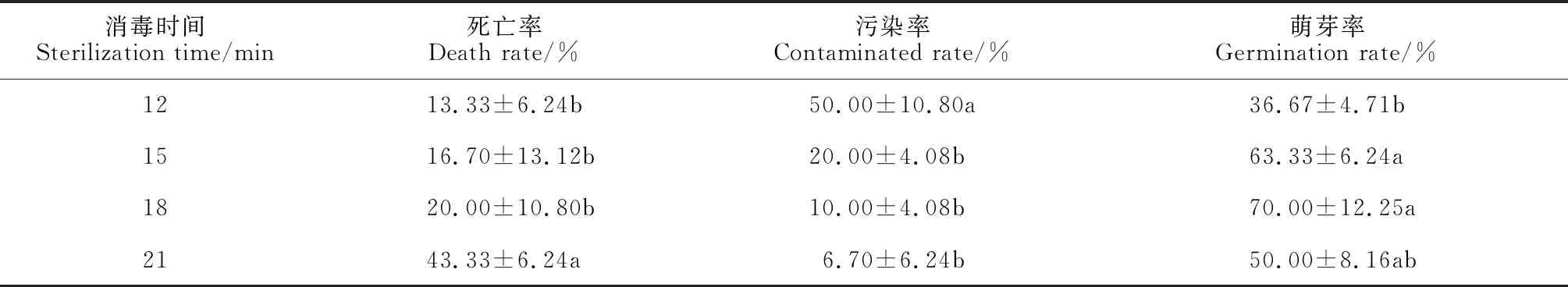
表1 不同消毒時間對外植體的處理結果Table 1 Treatment results of external implants at different disinfection times
2.2 激素NAA、2,4-D濃度對愈傷組織誘導的影響
待種子發芽后,接著轉至含不同激素種類和濃度的培養基中。一周左右胚軸的基部膨大,逐漸有愈傷組織形成,前期出現大量帶芽的愈傷組織。30 d后出愈率調查結果(表2)顯示。2,4-D對愈傷組織的誘導作用明顯優于NAA。在1.0 mg·L-16-BA基礎上,同濃度2,4-D的愈傷組織誘導率顯著高于NAA,并且愈傷組織的生長質量差別明顯,具體表現為添加NAA的愈傷組織生長緩慢,致密較硬(圖版Ⅰ,A);添加2,4-D的愈傷組織生長較快,松散柔軟(圖版Ⅰ,B、C)。另外,隨2,4-D濃度升高,外植體出愈數和出愈率均逐漸增加。其中,在1.0 mg·L-16-BA基礎上,當2,4-D濃度為2.0 mg·L-1時,外植體出愈率最高達到83.33%。可見,2,4-D處理的愈傷組織誘導率和生長質量均優于NAA,且適當提高2,4-D濃度有助于愈傷組織的形成。
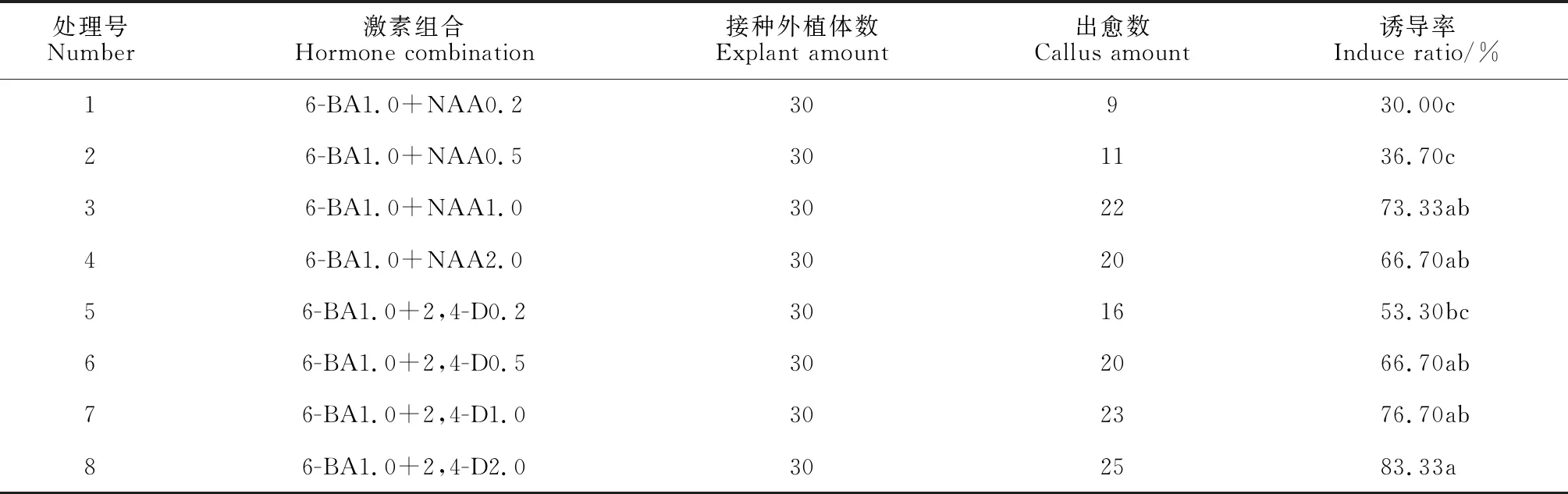
表2 不同濃度NAA、2,4-D誘導下外植體種子出愈率Table 2 The seed healing rate of explanst under different concentrations of NAA and 2,4-D
2.3 激素6-BA/NAA配比對不定芽誘導率的影響
選取誘導獲得的優質愈傷組織轉接到各種誘導不定芽的培養基中,經過1周培養,淡綠色愈傷組織形成很多突起,近而形成多芽體,增殖培養過程中,不定芽基部有毛狀根產生(圖版Ⅰ,D、E)。9種濃度6-BA/NAA組合處理對腋芽誘導效果(表3)存在差異,愈傷組織不定芽誘導率存在顯著差異(P<0.05)。其中,培養基中添加2.0 mg·L-16-BA+0.2 mg·L-1NAA時,不定芽誘導率最高(90.00%±8.20%),芽苗長勢好,葉色濃綠(圖版Ⅰ,F);激素組合3.0 mg·L-16-BA+0.2 mg·L-1NAA處理效果次之,誘導率為83.33%±4.71%,芽苗生長較快,葉黃綠,莖粗壯且長,出現毛狀根(圖版Ⅰ,G);激素組合2.0 mg·L-16-BA+1.0 mg·L-1NAA和3.0 mg·L-16-BA+1.0 mg·L-1NAA處理下,不定芽誘導率處于63.33%~73.33%之間;在2.0 mg·L-16-BA+2.0 mg·L-1NAA和1.0 mg·L-16-BA+2.0 mg·L-1NAA處理下,不定芽的誘導率稍高于50%;其余3種激素組合處理的不定芽誘導率均低于50%。

表3 不同6-BA/NAA配比培養基中不定芽誘導與生長的情況Table 3 The adventitious bud induction and growth states in the medium with different 6-BA/NAA ratios
2.4 激素濃度及配比對組培苗生根的影響
將經繼代培養后高達3 cm的小植株,接種于添加不同激素的1/2MS生根培養基上。10 d左右開始出現白色根原基,20 d左右組培苗莖段底部逐漸出現白色根,1個月后小苗生根趨于穩定。統計結果(表4)表明,各激素組合處理組培苗的生根數和生根率差異顯著(P<0.05)。
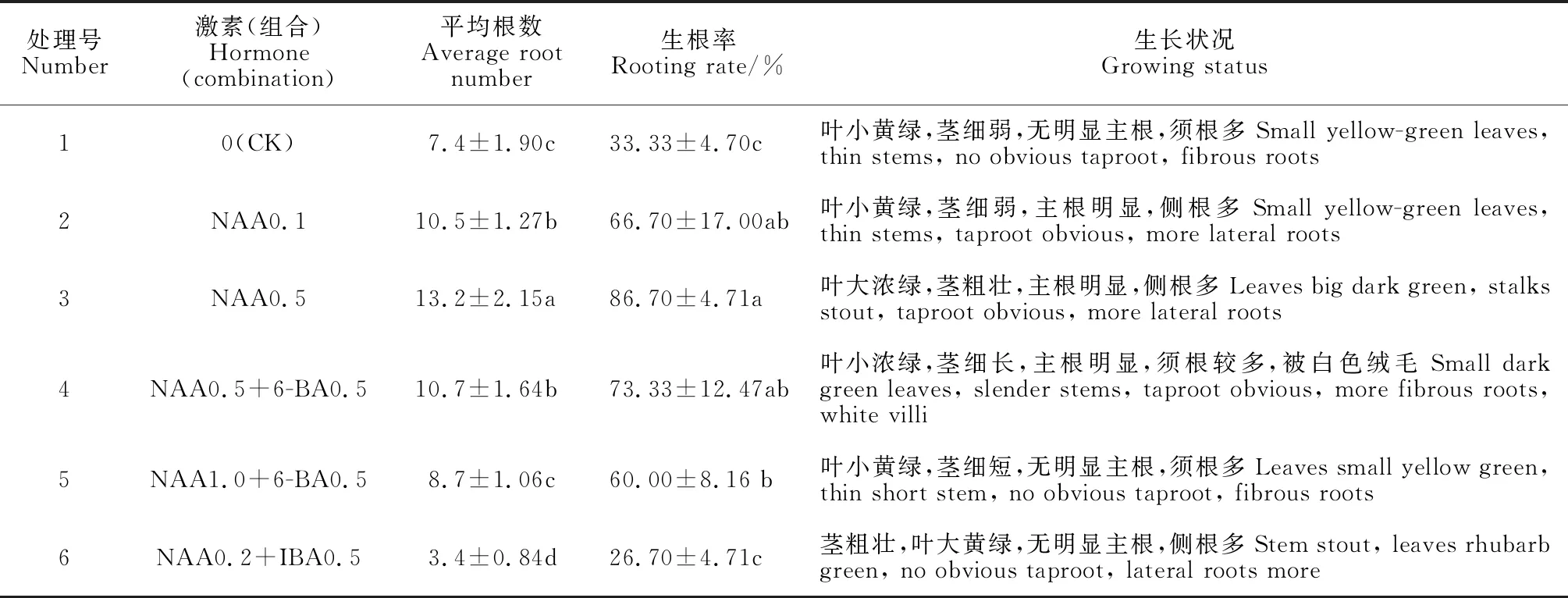
表4 不同激素及其組合培養基中組培苗生根情況Table 4 The rooting culture status of seedling in medium with different hormones and their combinations
其中,1/2MS生根培養基中單一添加0.5 mg·L-1NAA時,組培苗平均生根數為13.2條,生根率為86.70%,顯著高于不添加激素的對照,也明顯高于其余添加激素處理;培養基在添加0.5 mg·L-1NAA基礎上,再添加0.5 mg·L-16-BA時,組培苗的平均根數為10.7條,生根率為73.33%,與添加0.1 mg·L-1NAA處理的接近,但前者組培苗生長良好,后者組培苗莖較細弱,葉小黃綠(圖版Ⅰ,K);培養基添加0.5 mg·L-16-BA,同時加入1.0 mg·L-1NAA時,組培苗生根數大幅下降,與對照處于相同水平,其生根率也明顯下降,但仍顯著高于對照;培養基添加0.2 mg·L-1NAA+0.5 mg·L-1IBA時,組培苗生根數更少,僅有對照的45.95%,生根率大幅下降,稍低于對照(圖版Ⅰ,I)。可見,組培苗適宜生根培養基為1/2MS+0.5 mg·L-1NAA和1/2MS+0.5 mg·L-1NAA+0.5 mg·L-16-BA(圖版Ⅰ,H、J)。

圖版Ⅰ 穿龍薯蕷組織培養中愈傷組織、不定芽、生根及組培苗和實生苗A.添加NAA形成的愈傷組織;B、C.添加2,4-D形成的愈傷組織;D.愈傷組織誘導出芽;E.毛狀根;F.2.0 mg·L-1 6-BA+0.2 mg·L-1 NAA不定芽誘導培養基;G.3.0 mg·L-16-BA+0.2 mg·L-1 NAA不定芽誘導培養基;H.1/2MS+0.5 mg·L-1 NAA組培苗根部;I.1/2MS+0.2 mg·L-1 NAA+0.5 mg·L-1IBA組培苗根部;J.1/2MS+0.5 mg·L-1 NAA生根培養基;K.1/2MS+0.1 mg·L-1 NAA生根培養基;L.實生苗Plate Ⅰ Callus,adventitious bud,rooting,tissue culture seedling and seedling of Dioscorea nipponicaFig.A.Callus formed by adding NAA;Fig.B,C.Callus formed by adding 2,4-D;Fig.D.Callus induced budding;Fig.E.Hairy root;Fig.F.2.0 mg·L-1 6-BA+0.2 mg·L-1 NAA adventitious bud induction medium;Fig.G.3.0 mg·L-1 6-BA+0.2 mg·L-1 NAA adventitious bud induction medium;Fig.H.1/2MS+0.5 mg·L-1 NAA the roots of tissue culture seedlings;Fig.I.1/2MS+0.2 mg·L-1 NAA+0.5 mg·L-1 IBA the roots of tissue culture seedlings;Fig.J.1/2MS+0.5 mg·L-1 NAA rooting medium;Fig.K.1/2MS+0.1 mg·L-1 NAA rooting medium;Fig.L.Seedling
2.5 組培苗和實生苗營養器官解剖結構比較
穿龍薯蕷組培苗的葉片、纏繞莖和根狀莖在解剖結構上與實生苗存在差異(表5)。首先,葉片厚度、上表皮厚度、主脈維管束面積都表現為實生苗顯著高于組培苗;柵欄組織厚度二者之間差異不明顯,但組培苗的海綿組織厚度極顯著高于實生苗,導致二者的柵欄組織厚度/海綿組織厚度(P/S)差異顯著;組培苗葉片主脈維管束面積較小(1 779.81 μm2),約為實生苗的1/2(圖版Ⅱ,A、B;表5)。
其次,組培苗和實生苗在解剖結構的差異以纏繞莖尤為顯著。組培苗纏繞莖的內徑、導管面積和維管束數量均小于實生苗,但其維管束面積較大,但導管面積/維管束面積較小(0.06),為實生苗的1/2(表5)。實生苗纏繞莖皮層比例小,根據細胞形態可明顯區分為厚角組織、厚壁組織與薄壁組織,棱角處厚角組織發達(2~3層),環狀厚壁組織由2~3層細胞組成(圖版Ⅱ,C);組培苗皮層比例大,且無厚角組織,厚壁組織分化不甚完全,未形成明顯環帶,多數為排列疏松的薄壁細胞,細胞內富含淀粉粒(圖版Ⅱ,D)。實生苗纏繞莖維管束2輪,均為有限維管束,其中靠近皮層一側的維管束較小,為外韌維管束,排列疏松;靠近髓部的維管束較大,為雙韌維管束,排列緊密,與外側的外韌維管束相互間隔排列(圖版Ⅱ,E);組培苗纏繞莖僅有雙韌維管束,排列極為分散(圖版Ⅱ,F)。
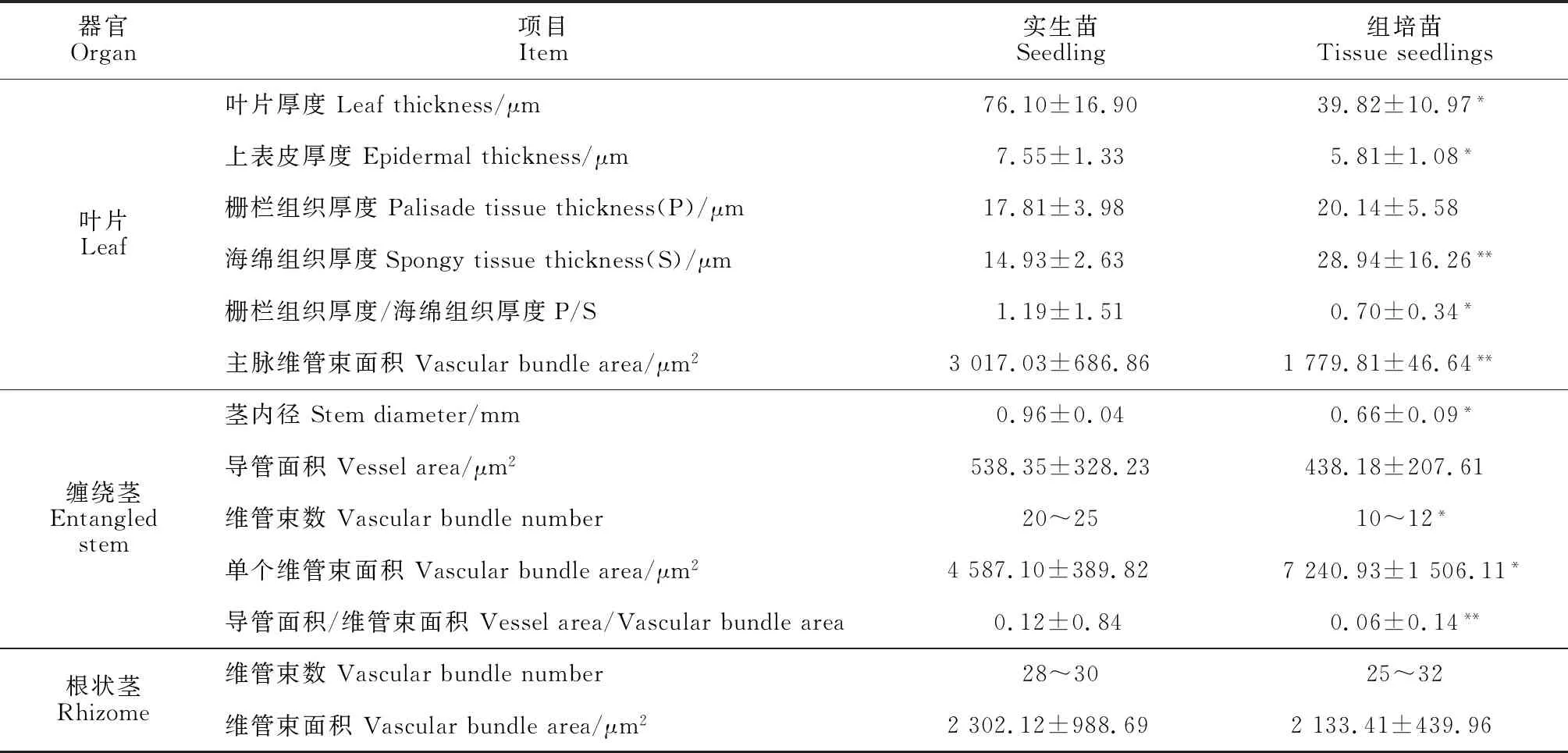
表5 組培苗和實生苗營養器官結構比較Table 5 Comparison of vegetative organ structure between tissue culture seedlings and seedlings
另外,根狀莖由周皮、基本組織和散生其中的外韌維管束組成(圖版Ⅱ,G、H)。組培苗和實生苗根狀莖解剖結構差別不大,維管束數和維管束面積在實生苗與組培苗之間差異均不顯著(表5)。

圖版Ⅱ 組培和實生苗營養器官解剖結構比較Xy.木質部;Ph.韌皮部;Vb.維管束;Co.厚角組織;Sc.厚壁組織;Pa.薄壁組織;Pc.薄壁細胞A.實生苗葉中脈;B.組培苗葉中脈;C.實生苗纏繞莖表皮;D.組培苗纏繞莖表皮;E.實生苗纏繞莖;F.組培苗纏莖;G.實生苗根狀莖橫切;H.組培苗根狀莖橫切Plate Ⅱ Anatomical structure comparison of vegetative organ between tissue culture and live seedlingsXy.Xylem;Ph.Phloem;Vb.Vascular bundle;Co.Collenchyma;Sc.Sclerenchyma;Pa.Parenchyma;Pc.Parenchymal cellFig.A.Midvein of seedling leaf;Fig.B.Midvein of tissue culture seedling leaf;Fig.C.Entangled stem epidermis of seedlings;Fig.D.Entangled stem epidermis of tissue culture seedlings;Fig.E.Seedlings entangled stem;Fig.F.Tissue culture seedlings entangled stem;Fig.G.Seedling rhizome transverse;Fig.H.Rhizome of tissue culture seedlings are transverse
3 討 論
3.1 穿龍薯蕷再生體系建立
愈傷組織誘導是建立植物再生體系或進行遺傳轉化等的重要途徑,在快速繁殖、育種、轉基因、脫毒苗生產等方面均有極大的應用價值[14]。彭金環等[15]對穿龍薯蕷愈傷組織誘導研究中,以不同外植體為材料誘導愈傷組織,發現各種營養器官均能誘導出愈傷組織,但外植體存在不同程度的褐化,嚴重影響了愈傷組織的生長。利用種子誘導出的愈傷組織,生長速度快,出愈率比較高,而且因為沒有機械損傷,很大程度上抑制了褐化的發生。但在愈傷組織誘導過程中,種子須先平鋪于MS培養基中,待種子長出芽和胚軸再將其接種到誘導愈傷培養基中。如若直接接種至愈傷組織培養基中,出愈率很低,2,4-D會干擾生長素的分布模式,使整個胚胎中的生長素信號變強,對植物胚軸形成有抑制作用[16]。培養基中添加的激素類型及濃度對體外繁殖有重要影響。植物激素是影響薯蕷愈傷組織誘導和分化再生植株的重要因素,不同的外植體誘導率和生長差異很大,植物激素的種類和配比濃度也要隨之變化。張數鑫[17]在試驗中發現穿龍薯蕷不同外植體的誘導率和生長差異很大,并以莖尖誘導率最高,莖段次之。
陳鳳清[18]采用無菌帶芽莖段為外植體,通過多芽體誘導、微塊莖形成、根誘導和增殖成功建立了再生體系。同時提出,激素的協同作用影響芽、根的形成,不同濃度激素適當組合對組織培養更有利。在植物生長調節劑中,生長素和細胞分裂素是影響離體試管苗生芽和生根的主要決定因素。植物細胞培養中的器官發生(類型和范圍)取決于生長素與細胞分裂素的比例[19]。細胞分裂素,如KT和6-BA,已被證明能促進細胞分裂、芽增殖、芽形態發生和抑制根部形成;而生長素,如NAA和2,4-D通常用于刺激愈傷組織產生和細胞生長,啟動萌芽和生根,誘導體細胞胚胎發生,并刺激莖尖和莖段的生長[20-23]。本研究以種子為外植體誘導愈傷組織最適培養基為MS+1.0 mg·L-16-BA+2.0 mg·L-12,4-D,形成愈傷組織長勢好,分化能力強,在6-BA濃度相同的情況下,2,4-D誘導作用優于NAA,這與彭金環研究結果一致;愈傷組織分化的適宜培養基為MS+2.0 mg·L-16-BA+0.2 mg·L-1NAA,芽苗長勢好,葉色濃綠并且在誘導愈傷組織分化芽過程中產生了毛狀根,后期生根培養中產生的根更加健壯。有研究發現,毛狀根生長快,次生代謝產物含量高,遺傳穩定,是工業化生產所向往的,可以實現藥用植物次生代謝物的工廠化生產[17,24-25]。
3.2 穿龍薯蕷組培苗和實生苗營養器官解剖結構的差異
組培苗生長的培養瓶中環境條件與室外差別很大,小苗幼嫩,一直處于可控的環境下生長,抗性較低。因此,必須經過煉苗來提高組培苗對外界環境的適應性,達到提高移栽成活率的目的,而煉苗成功與否關系到整個組培過程能否成功完成[26]。本研究通過比較穿龍薯蕷組培苗和實生苗葉片、纏繞莖和根狀莖的解剖結構發現,組培苗葉片形態、結構變異較大,葉片更薄,柵欄組織和海綿組織分化不完全,P/S值低,其原因可能是組培苗的培養光照較弱,但營養和水分充足,幼苗適應這樣的生長條件作出相應的形態與結構調整[27]。而P/S值常被用作衡量植物抗寒[28]、抗旱[29-30]能力和光合速率高低[31]的重要指標,弱光下的組培苗葉片較薄,P/S值隨之降低。因此在煉苗移栽初期要注意環境的通透性、保水性以及透光性。組培苗的纏繞莖解剖結構也與實生苗存在顯著差異。很多植物通過增加莖的表皮厚度來降低蒸騰作用,減少水分蒸發,從而抵抗干旱[32-33]。本研究中穿龍薯蕷實生苗纏繞莖的表皮厚于組培苗,皮層分化出厚角組織與厚壁組織,維管束數量多,排列較緊密,而組培苗皮層分化簡單,維管束數量少,排列分散,可能由于組培苗處于高濕的可控環境,水分營養充足,機械組織與輸導組織的結構隨之簡化。
移栽馴化過程是組培苗組織結構分化發育日趨完善、逐漸適應外界環境的過程。也是組培苗由半自養半異養向完全自養轉變的過程[34]。穿龍薯蕷組培苗和實生苗解剖結構表觀特征的變化是其適應不同生態環境的具體表現,因此,試管苗在向大田移栽過程中,要想獲得較高的成活率,使其能夠適應外在自然環境,解剖結構特征發生相應的環境適應性非常重要,在移栽馴化初期應重點保障移栽小環境的濕度與光照,以提高其成活率。
4 結 論
本試驗進行了穿龍薯蕷種子組培體系優化研究,并在此基礎上開展組培苗和實生苗營養器官解剖結構的對比研究,旨在為后期研究穿龍薯蕷有效藥用成分,以及實現穿龍薯蕷生產的產業化,促進穿龍薯蕷種植產業的健康發展提供技術支撐。結果表明:(1)2% NaClO作為種子誘導外植體消毒劑,消毒時間以15和18 min為佳。(2)MS+1.0 mg·L-16-BA+2.0 mg·L-12,4-D為愈傷組織誘導的最適培養基。(3)MS+2.0 mg·L-16-BA+0.2 mg·L-1NAA為愈傷組織分化芽的最適培養基。(4)1/2MS+0.5 mg·L-1NAA為最適生根培養基。(5)組培苗和實生苗解剖結構比較發現,組培苗煉苗移栽初期要注意環境的通透性、保水性以及透光性,使組培苗能夠適應外在自然環境,從而提高其成活率。
