顯性效應對蘇淮豬肉色性狀遺傳評估和基因組選擇的影響
2021-05-28 04:14:00劉航侯黎明王彬彬劉晨曦陶偉張總平牛培培李強李平華黃瑞華
畜牧與獸醫 2021年6期
劉航,侯黎明,王彬彬,劉晨曦,陶偉,張總平,牛培培,李強,李平華,4,5*,黃瑞華,4*
(1. 南京農業大學養豬研究所,江蘇 南京 210095;2. 南京農業大學淮安研究院,江蘇 淮安 225001;3. 淮安市淮陰新淮種豬場,江蘇 淮安 223322;4. 江蘇現代農業(生豬)產業體系集成創新中心,江蘇 南京 210095;5. 淮安沐林新農村發展研究有限公司,江蘇 淮安 225001)
從傳統的表型選育,到最佳線性無偏預測(best linear unbiased prediction, BLUP),再到標記輔助選擇(marker assisted selection, MAS),育種技術得到了快速的發展,極大地促進了生豬品種的遺傳改良[1-2]。隨著Meuwiseen等[3]提出基因組選擇(genomic selection, GS),遺傳育種技術得到進一步的發展。基因組選擇將基因組信息考慮進估計育種值(estimated breeding values, EBVs)的BLUP方法中,即基因組最佳線性無偏預測(genomic best linear unbiased prediction, GBLUP)[4]。目前許多育種跨國企業如種豬改良公司(pig improvement company, PIC)、丹育、海波爾育種有限公司(Hypor)等已將GS應用于豬的育種中,國內以溫氏為代表的大型育種公司也紛紛開啟豬GS育種工作[5]。近年來肉質性狀開始被各大育種公司納入育種目標。肉色性狀是肉質性狀中最影響消費者購買欲的指標。肉色性狀作為微效多基因控制的數量性狀,其遺傳力屬于中或低等,且表型測定需要通過屠宰才準確,因此基于系譜和常規選擇方法遺傳參數預測的準確性仍達不到理想程度,需要采用基因組選擇方法進行遺傳參數估計和高效育種[6]。
決定數量性狀的多基因效應分為加性效應、顯性效應和上位效應,其中加性效應值為估計育種值,反映了目標動物在育種上貢獻的大小。復雜性狀的遺傳結構中主要包括基因的數量和效應大小、基因間的互作、基因與環境間的互作等。以肉色為代表的肉質性狀的遺傳結構復雜,然而目前常用的GS方法只將加性效應加入模型中。顯性效應為非加性效應,是同一位點上2個等位基因之間的相互作用[7]。Su等[7]利用4種不同遺傳效應組合的模型估計杜洛克豬平均日增重(30~100 kg)的遺傳力為0.357~0.397,該性狀的顯性遺傳方差占總表型方差的5.6%。研究發現當顯性遺傳方差和總表型方差的比率達到0.2時,估計模型中加入顯性效應時,其準確性會明顯提高[8]。王延暉等[9]利用模擬數據比較了不同模型下基因組估計育種值(genomic estimated breeding value, GEBV)估計的準確性,結果表明當顯性遺傳方差在遺傳方差中的比重越高,顯性效應對GEBV估計的準確性影響越大。有研究表明顯性效應不僅能提升復雜性狀GEBV預測的準確性,同時可用于指導個體間的交配計劃來提高后代的總遺傳價值[10]。蘇淮豬含有75%大白豬和25%淮豬的血統,其作為國內優良的雜交品種豬,具有生長發育快、繁殖率高、肉質鮮美等特點[11]。目前尚未有團隊分析顯性效應對豬肉色性狀的影響,因此本研究以蘇淮豬為試驗動物,利用DMU軟件的單性狀模型和平均信息約束最大似然(average information restricted maximum likelihood, AI-REML)算法,比較包含和不包含顯性效應的模型下,GBLUP方法估計豬肉色性狀加性遺傳方差和顯性遺傳方差的變化,并評估GEBV預測的準確性。
1 材料與方法
1.1 試驗動物
本研究從江蘇省淮安市淮陰種豬場選取487頭蘇淮豬,其中291頭閹公豬和196頭母豬,分為4個批次在相同飼養條件下飼養后屠宰。屠宰前禁食24 h,自由飲水,于淮安市金源肉品中心進行屠宰。屠宰日齡為(239.4±2.2)d,胴體重為(61.25±0.46)kg,并依據屠宰時的氣候狀況分為4個季節(3、4、5月為春季,6、7、8月為夏季,9、10、11月為秋季,12、1、2月為冬季)。
1.2 樣品采集及表型測定
采集試驗動物的耳組織樣,用于基因組DNA提取[12]。采集胸腰結合處背最長肌組織樣品,用于測定肉色性狀。在屠宰后45 min,隨機選擇背最長肌橫截面的3塊區域,使用HunterLab MiniScan EZ (New York, USA)儀器測定肉色,通過國際照明委員會(International Commission on Illumination, CIELAB)色度坐標(亮度L*、紅度a*和黃度b*)參數表示,并記為L*45 min、a*45 min和b*45 min,求其平均值。置于4 ℃下保存24 h,再次測定背最長肌的3個隨機區域的肉色參數,并記為L*24 h、a*24 h和b*24 h,求其平均值。
1.3 基因組DNA提取、分型及質控
利用DNA提取試劑盒(天根生化科技有限公司,北京)提取耳組織樣DNA,通過NanoDrop 2000分光光度計測量DNA原液濃度,DNA原液純度指標(OD260 nm/OD280 nm)值在1.8~2.1之間。利用GGP Porcine 80K SNP 芯片(紐勤生物科技有限公司,中國上海)對合格的DNA原液進行SNP分型,共獲得68 516個SNP。
通過PLINK(V1.07)軟件對SNP分型結果進行質控,并去除位于性染色體和未知染色體上的SNP,質控條件如下:1)最小等位基因(minor allele frequency, MAF)大于 1%;2)SNP檢出率大于95%;個體檢出率大于90%。質控后,保留487個個體和55 035個SNPs,用于后續的分析。
1.4 統計模型及分析方法
本試驗先通過SAS 9.2軟件的一般線性模型(general linear model, GLM)分析每個肉色性狀的影響因素,將顯著影響肉色性狀的因素(P<0.05)考慮進后續分析對應模型中。一般線性模型如下:
y=μ+S+B+E+A+C+e。
模型中y代表肉色性狀表型值;μ代表群體均值;S代表性別;B代表屠宰批次;E代表屠宰季節;A代表日齡;C代表胴體重;e代表隨機殘差。
基于DMU軟件的單性狀模型和AI-REML算法,通過GBLUP方法估計蘇淮豬群體中肉色性狀的遺傳方差和GEBV。本試驗使用的遺傳力為狹義遺傳力,即加性遺傳方差與表型方差的比值。GBLUP統計模型如下:
GBLUP-A:y=Xb+Maqa+e,
GBLUP-AD:y=Xb+Maqa+Mdqd+e。
GBLUP-A為只考慮加性效應的統計模型,GBLUP-AD為同時考慮加性效應和顯性效應的統計模型。模型中y表示肉色性狀表型向量;b表示固定效應向量(L*值考慮屠宰批次、屠宰季節和胴體重;a*值和b*值考慮屠宰批次、屠宰季節和日齡),X表示與固定效應的關聯矩陣;qa和qd分別為加性效應向量和顯性效應向量,Ma和Md分別為加性向量和顯性向量的關聯矩陣;e為隨機殘差效應。
本試驗使用R3.6.3軟件循環進行5重交叉驗證用于檢測模型預測的準確性,共循環20次。5重交叉驗證先將試驗群體隨機等分為5份,依次將1份群體作為驗證群體,4份作為參考群體,將驗證群體預測的GEBV和矯正表型間的相關系數除以遺傳力的平方根作為預測準確性[13]。
數據以“平均值±標準誤”表示,P<0.01為差異極顯著,P<0.05為差異顯著。
2 結果與分析
2.1 蘇淮豬肉色性狀描述統計及影響因素
487頭蘇淮豬肉色性狀如表1所示,a*值的變異系數最大,L*值和b*值變異系數也較大。

表1 蘇淮豬肉色性狀的描述統計(n=487)
表2為GLM模型對肉色性狀影響因素顯著性檢驗的結果。在該蘇淮豬試驗群體內,屠宰批次和屠宰季節對肉色性狀均有顯著影響(P<0.05)。L*值在夏季時最高,春季時最低;而a*值和b*值在春季時最高,夏季時最低。隨著日齡增加,a*值極顯著上升(P<0.01),而b*值則顯著下降(P<0.05)。L*45 min和L*24 h隨著胴體重增加而顯著下降(P<0.05)。因此在估算肉色GEBV的GBLUP-A模型和GBLUP-AD模型中,屠宰批次和屠宰季節作為固定效應,日齡作為a*值和b*值的協變量,胴體重作為L*值的協變量。
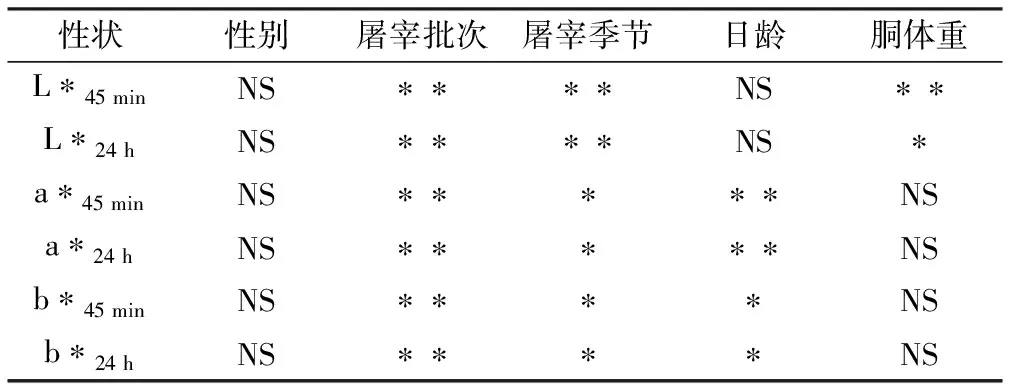
表2 肉色性狀影響因素的顯著性檢驗
2.2 不同模型估計的遺傳方差
由表3可見,a*45 min的遺傳力在GBLUP-A模型和GBLUP-AD模型中都最高,分別為0.32和0.27,而b*24 h在GBLUP-AD模型中獲得最低的遺傳力,為0.13。L*、a*和b* 3個肉色性狀受顯性效應影響程度不一,b*值在GBLUP-AD模型中顯性遺傳方差與加性遺傳方差的比率可以達到2.0,而L*24 h估計的顯性遺傳方差幾乎為0。顯性效應對于不同性狀加性效應估計的影響也不同,b*45 min的顯性遺傳方差與加性遺傳方差的比率最大,但其在GBLUP-A模型和GBLUP-AD模型的加性遺傳方差基本上未發生改變,因此遺傳力也未被影響。顯性效應對b*24 h具有較大影響,其遺傳力下降了0.06。a*值受顯性效應的影響也較大,a*45 min和a*24 h的顯性遺傳方差與加性遺傳方差的比率分別為0.53和1.40,其遺傳力都下降了0.05。而L*值受顯性效應的影響最小,L*24 h基本上不受影響,L*45 min的遺傳力只下降了0.01。
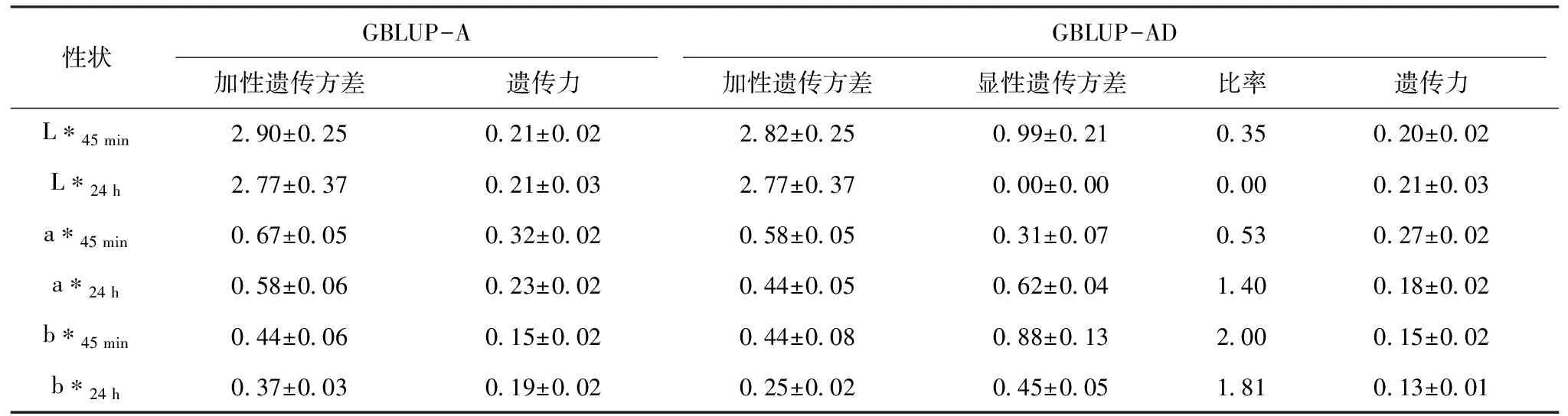
表3 不同模型下估計肉色的遺傳參數
2.3 GBLUP預測準確性比較
GBLUP預測不同性狀的GEBV時,其準確性不同。由表4可見,b*值的預測準確性最高,其中b*24 h的GEBV預測準確性在GBLUP-AD模型中達到0.790 6。a*45 min的GEBV預測準確性在GBLUP-A模型中最低,為0.448 7。比較了顯性效應對肉色性狀GEBV預測準確性的影響,除了L*24 h和b*45 min在GBLUP-A模型和GBLUP-AD模型中的GEBV預測準確性沒有明顯差異,L*45 min、a*45 min、a*24 h和b*24 h在GBLUP-AD模型中的GEBV預測準確性分別高于GBLUP-A模型。b*24 h在GBLUP-A模型和GBLUP-AD模型中的預測準確性都高于b*45 min,而a*24 h與a*45 min也呈現相同的趨勢。
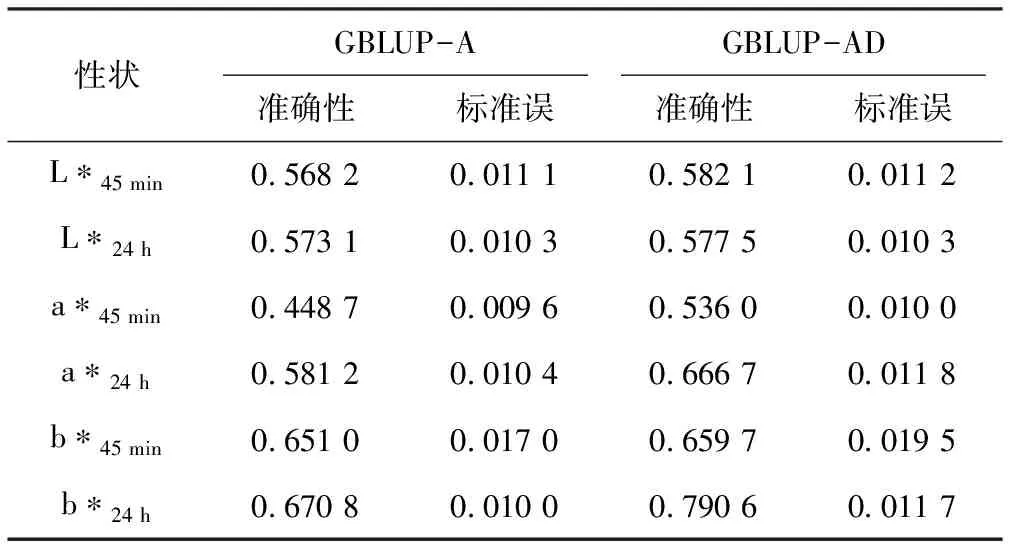
表4 不同模型預測肉色GBEV的準確性
3 討論
3.1 蘇淮豬肉色性狀的影響因素
蘇淮豬群體內肉色性狀存在較大的變異,其中a*45 min的變異系數達到32.22%,a*24 h的變異系數達29.33%,L*和b*也存在相對較大的變異,因此需對蘇淮豬肉色性狀開展選育工作。傳統的肉色性狀評定主要通過比色板進行,常用的有日本肉色比色板和美國肉色比色板,但這兩種測定方法主觀誤差較大[14]。本試驗采用CLELAB色度坐標(L*、a*和b*)對肉色性狀進行評定,相比于比色板更加科學客觀[15]。肉色參數L*值與肌纖維類型和肌肉中糖原含量相關,a*值與肌肉中肌紅蛋白含量相關,而b*值與脂肪含量相關[16]。因此影響3個肉色參數的主要因素并不相同。屠宰批次和屠宰季節對肉色具有顯著影響,表明肉色受環境影響較大。Simonetti等[17]比較了不同屠宰前環境溫度對肉品質的影響,發現當屠宰前環境溫度高時,豬肉在屠宰后24和48 h均呈現出明顯較低的pH值和a*值,明顯較高的滴水損失和L*值。因此在生產中應該控制屠宰環境的變化,降低對肉品質的損害。
3.2 遺傳結構對蘇淮豬肉色性狀遺傳力的影響
在蘇淮豬群體中,肉色的遺傳力在0.13~0.32之間,屬于低至中等遺傳力,其中a*45 min在GBLUP-A模型中估計的遺傳力最高達到0.32,b*24 h在GBLUP-AD模型中估計的遺傳力最低,為0.13。不同群體間的肉色遺傳力略有差異,但都屬于低至中等遺傳力。Cabling等[18]計算690頭杜洛克豬的肉色遺傳力,L*、a*和b*的遺傳力分別為0.44、0.68和0.64。而Miar等[19]利用杜洛克和大白豬雜交產生的2 075頭后代測定的肉色遺傳力略低,L*、a*和b*的遺傳力分別為0.28、0.26和0.31。屠宰后45 min和24 h的分別代表著熱鮮肉和冷鮮肉,具有不同的經濟價值,其中a*值和b*值在2個時間點也體現出不同的遺傳力。本文發現蘇淮豬a*值在45 min的遺傳力明顯高于24 h的遺傳力,b*24 h則高于b*45 min。這可能是由于肌紅蛋白的氧化狀態容易受保存環境的影響,而脂肪則在4 ℃環境下趨于穩定狀態。
不同性狀的加性遺傳方差與非加性遺傳方差的比例不同,一般來說,低遺傳力的性狀相比于高遺傳力的性狀更容易受到非加性效應的影響。Tusell等[20]通過在純合群體和雜合群體中對豬的6類性狀進行遺傳貢獻的估計,發現不同性狀其遺傳表達模型不同,其中生長性狀受顯性效應影響較小,而肉質和行為性狀則受顯性效應影響較大。Liu等[21]對杜洛克豬和皮特蘭豬雜交的品系進行35個表型的數量性狀遺傳位點(quantitative trait locus, QTL)定位,共鑒定到71個QTL,其中40個QTL加性效應較強,31個顯性效應較強,值得注意的是與肉質性狀相關的11個QTL的顯性效應值與加性效應值的比率在0.3~3.0之間,這與本研究結果一致,表明肉質性狀受顯性效應影響較大。Stratz等[22]也對梅山豬、歐洲野豬和皮特蘭形成的雜交系進行與肉質相關QTL的鑒定,鑒定到10個同時具有加性效應和顯性效應的QTL,3個只具有顯性效應的QTL,5個只具有加性效應的QTL。本次試驗中,顯性遺傳方差與加性遺傳方差的比率在b*值上表現最高,在L*值上表現最低,表明b*值受顯性效應影響較大,而L*值則較小。b*45 min在GBLUP-A模型和GBLUP-AD模型中的遺傳力并未太大變化,而b*24 h在GBLUP-AD模型中的遺傳力下降了0.06,這可能是由于影響b*45 min和b*24 h的基因存在差異,b*24 h受環境影響更大。a*45 min和a*24 h受到顯性效應不同程度的影響,但其遺傳力都下降了0.05,且a*24 h的遺傳力更低,即顯性效應對a*24 h遺傳力估計有更大的影響,而顯性效應對于L*值的影響較小。肉色受到微效多基因控制,其遺傳結構復雜,在估計肉色遺傳力的模型中考慮不同的遺傳效應有助于提高估計的準確性。
3.3 不同遺傳結構對GEBV預測準確性的影響
隨著測序成本的下降,基因組數據可以加入育種值估計的模型中,這大幅度提升了模型的準確性。GBLUP的模型可分為直接法和間接法,直接法利用基因組數據構建的親緣關系矩陣代替了系譜構建的親緣關系矩陣考慮進動物模型中;間接法則通過參考群體來估計各個SNP的效應值,將驗證群體的各SNP效應值相加為GEBV[23]。為提高動物模型預測的準確性,多種模型被開發,如一步法(single-step, SSBLUP)、性狀特異關系矩陣(trait-specific relationship matrix, TABLUP)、嶺回歸最佳線性無偏預測法(ridge regression best liner unbiased prediction, RRBLUP)和貝葉斯方法(如BayesA、BayesB、BayesC、BayesCπ和BayesLASSO)等[24]。本試驗采用GBLUP的直接法,當性狀遺傳力低且受到多個微效QTL控制時,該方法預測準確性與貝葉斯方法、SSBLUP等模型相近[25]。在預測肉色性狀GEBV的模型中,性狀遺傳力越高,其模型預測準確性也就越高,但同時也存在多種因素影響,如性狀遺傳結構、相關QTL數量、芯片SNP的分布及數量、有效群體數量等[26]。
本次試驗中L*24 h的顯性遺傳方差接近為0,而其余5個性狀都受到顯性效應的影響,可能是由于環境對L*24 h產生較大的影響。結果表明L*24 h和b*45 min在GBLUP-A模型和GBLUP-AD模型中的準確性并未有明顯差異,其余性狀在GBLUP-AD模型中的準確性均有所提高,其中b*24 h的預測準確性在GBLUP-AD模型中比在GBLUP-A模型中約高了0.12。對于存在顯性效應影響的性狀,模型中考慮顯性效應會增加模型的預測準確性。Nishio等[27]通過模擬數據來評估顯性效應對于豬GEBV預測準確性的影響,結果表明當顯性效應方差較大時,模型中加入顯性效應,其準確性相比于常規模型提高了1.2%。Da等[28]基于1 654個荷斯坦奶牛的基因組數據,比較了GBLUP和GREML方法在不同模型下的預測準確性,發現同時包含加性效應和顯性效應的GBLUP和GREML相比于只含單一遺傳效應的模型具有高的預測準確性。因此在動物模型中考慮顯性效應,可提高復雜性狀GEBV預測的準確性,提升育種效率。
綜上所述,本研究通過研究顯性效應對肉色GBLUP預測準確性的影響,在肉色6個性狀中L*24 h受顯性效應影響小,而L*45 min、a*45 min、a*24 h、b*45 min和b*24 h受顯性效應影響大。考慮顯性效應的模型在預測L*45 min、a*45 min、a*24 h和b*24 h的GEBV時展現出更高的準確性,這為蘇淮豬肉質性狀遺傳改良提供了更加合適的模型。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
核科學與工程(2021年4期)2022-01-12 06:30:26
今日農業(2020年19期)2020-12-14 14:16:52
小學生必讀(中年級版)(2020年9期)2020-12-04 02:07:22
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
中學物理·高中(2016年12期)2017-04-22 11:53:03
光學精密工程(2016年6期)2016-11-07 09:07:19
核科學與工程(2015年4期)2015-09-26 11:59:03
小櫻桃·童年閱讀(2014年11期)2014-12-01 22:21:30
