澳洲堅果蛋白磷酸酶基因MiSTPP1和MiSTPP4克隆與生物信息學分析
2021-05-31 00:41:00李季東楊祥燕蔡元保曾黎明黃思婕巫輔民林玉虹鄭文武黃錦媛
福建農業學報 2021年2期
李季東,楊祥燕,蔡元保,李 穆,曾黎明,黃思婕,巫輔民,林玉虹,鄭文武,黃錦媛
(廣西壯族自治區農業科學院/廣西壯族自治區亞熱帶作物研究所,廣西 南寧 530001)
0 引言
【研究意義】低溫、干旱、洪澇、鹽堿等逆境脅迫是嚴重影響植物生長發育的環境因子,可導致植物產量和品質下降甚至死亡。植物通過細胞功能調節來抵御外界環境的逆境脅迫,尤其是蛋白質可逆磷酸化在細胞的新陳代謝、生長發育、增殖與凋亡、信號轉導、基因調控等過程中發揮著重要的調控作用[1?2],而蛋白質可逆磷酸化主要由蛋白磷酸酶和蛋白激酶來平衡調控。因此,蛋白磷酸酶的功能研究對于了解蛋白質可逆磷酸化過程及植物耐逆機制研究具有重要意義。【前人研究進展】蛋白質磷酸酶分為絲氨酸/蘇氨酸磷酸酶、雙特異性蛋白磷酸酶、酪氨酸磷酸酶3類,其中絲氨酸/蘇氨酸磷酸酶(PP)又分為蛋白磷酸酶1(PP1)和蛋白磷酸酶2(PP2)[3]。目前,國內外對PP2家族蛋白的生物學功能比較清楚,如番茄(Solanum lycopersicum)LePP2Ac、水 稻(Oryza sativa)OsPp2a、擬 南 芥(Arabidopsis thaliana)AthPP2CA和玉米(Zea mays)ZmPP2C等[4?5];而對PP1家族蛋白的功能研究并不多,這個家族成員包括歐洲油菜(Brassica napus)BnPP1、野大豆(Glycine soja)GsPP1、豌豆(Pisum sativum)PsPP1和橙子(Citrus sinensis)CsPP1等[6]。PP1家族蛋白主要作用于磷酸酶激酶的β亞單位,其活性主要受內源熱穩定蛋白I-1和I-2的抑制[7]。雖然從模式植物擬南芥、水稻、煙草、苜蓿、豌豆等多種植物的基因組DNA或cDNA文庫中都分離到編碼PP1蛋白催化亞基的基因[6?8],但是對于這些基因的功能及其編碼蛋白的催化機制人們并不清楚。澳洲堅果又稱夏威夷果,是山龍眼科(Proteaceae)澳洲堅果屬(Macadamia)植物,原產于亞熱帶雨林,具有重要的營養價值和藥用價值,享有“干果皇后”的美譽。雖然澳洲堅果在澳大利亞、美國夏威夷、肯尼亞和中國等地廣泛種植,但是受其生長條件的限制,澳洲堅果無法進一步推廣種植[9?10]。【本研究切入點】目前,國內外仍未見澳洲堅果蛋白質磷酸酶的相關研究報道。在澳洲堅果抗寒轉錄組測序的基礎上,采用RT-PCR克隆澳洲堅果蛋白磷酸酶基因,并利用生物信息學分析這些蛋白磷酸酶的結構和功能。【擬解決的關鍵問題】基于前期的研究基礎[11],本研究從澳洲堅果光殼種中克隆編碼蛋白磷酸酶基因MiSTPP1和MiSTPP4,并對其進行生物信息學分析,以期為深入研究澳洲堅果PP1家族蛋白的結構和功能及闡明其耐逆的分子機制提供科學依據。
1 材料與方法
1.1 試驗材料
試驗材料為廣西亞熱帶作物研究所澳洲堅果種質資源圃提供的澳洲堅果光殼種(Macadamia integrifolia)品 種Own Choice(O.C.)。對 接 穗 為4個月苗齡的嫁接苗進行4 ℃低溫處理0.5、1、3、6、12、24 h,采集這些低溫處理后的幼嫩葉片,液氮 速凍處理后?80 ℃保存備用。
1.2 總RNA提取及cDNA合成
參照蔡元保等[12]的CTAB改良法提取澳洲堅果葉片總RNA,利用核酸蛋白檢測儀分析所提取的總RNA純度和濃度,用1.5%瓊脂糖凝膠電泳檢測提取的總RNA完整性,按照美國Invitrogen公司反轉錄試劑盒(Reverse Transcription System)說明書合成澳洲 堅果cDNA。
1.3 澳洲堅果的MiSTPP1和MiSTPP4基因克隆
利用本課題組從澳洲堅果抗寒轉錄組中獲得2個蛋白磷酸酶基因,利用Primer Premier 5.0軟件在這2個基因開放閱讀框(ORF)以外的區域設計引物MiSTPP1-F和MiSTPP1-R,以及MiSTPP4-F和MiSTPP4-R(表1)。所用引物全部由大連寶生物工程有限公司合成。
PCR反應體系15 μL,包括澳洲堅果cDNA模板1.0 μL,2×Ex Taq PCR MasterMix 8.0 μL,正向和反向引物各0.5 μL,無菌ddH2O補足至15 μL。PCR擴增程序:95 ℃預變性2 min;94 ℃ 30 s,52.0 ℃或55.2 ℃(具 體 參 考 表1)30 s,72 ℃ 90 s,32個 循環;72 ℃延伸10 min。PCR擴增產物經1.0%瓊脂糖凝膠電泳檢測后,采用天根瓊脂糖凝膠回收試劑盒(DP209-02)進行回收與純化,將其連接到T載體pMD18上,轉化到大腸桿菌DH5α感受態細胞,并采用藍白斑篩選方法挑取白色陽性克隆,送至上海 生工生物工程有限公司進行序列測序。
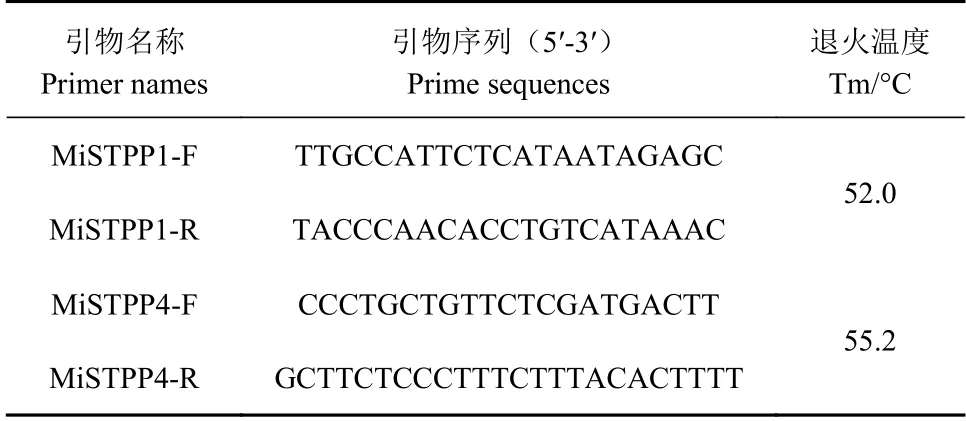
表 1 基因MiSTPP1和MiSTPP4克隆所用引物序列Table 1 Primer sequences used in cloning MiSTPP1 and MiSTPP4
1.4 生物信息學分析
利用NCBI網站的BLAST程序(https://blast.ncbi.nlm.nih.gov/Blast.cgi)進行蛋白質序列和功能結構域的同源檢索,用ORF Finder(http://www.ncbi.nlm.nih.gov/gorf/gorf.html)進行開放閱讀框ORF分析,用PROSITE(http://prosite.expasy.org/scanprosite/)和SMART(http://smart.embl-heidelberg.de/)軟件預測功能結構域,用ProtParam軟件(http://web.expasy.org/protparam/)分析目標蛋白質的理化性質,用在線程序NetPhos 3.1(http://www.cbs.dtu.dk/services/NetPhos/)預測磷酸化位點,用PSORT軟件(http://www.psort.org)分析蛋白質的亞細胞定位,用SignalP 4.1軟件(http://www.cbs.dtu.dk/services/SignalP/)預測蛋白信號肽,用TMpred軟件(http://www.ch.embnet.org/software/TMPRED_form.html)預測蛋白質的跨膜域,用SOPMA軟件(http://www.expasy.org)進行蛋白二級結構預測,用在線程序SWISS-MODEL(http://swissmodel.expasy.org/)預測蛋白質的三維結構。在NCBI網站GenBank中查找同源蛋白,并用DNAMAN 6.0軟件進行同源蛋白的系統進化樹構建。
2 結果與分析
2.1 澳洲堅果MiSTPP1和MiSTPP4基因的cDNA全長克隆
以澳洲堅果葉片cDNA為模板,采用特異性引物MiSTPP1-F和MiSTPP1-R,以及MiSTPP4-F和MiSTPP4-R分別擴增2個基因的開放閱讀框(ORF)(圖1)。測序結果表明,澳洲堅果MiSTPP1基因的cDNA全長為2 119 bp,ORF長度為1 053 bp,編碼350個氨基酸;MiSTPP4基因的cDNA全長為2 030 bp,ORF長度為981 bp,編碼326個氨基酸。利用NCBI網站的BLAST程序檢索同源蛋白的結果表明,這2個基因的編碼蛋白和其他物種的絲氨酸/蘇氨酸磷酸酶PP1具有很高的序列相似性。因此,將這2個基因分別命名為澳洲堅果的MiSTPP1和MiSTPP4基因,GenBank登錄號分別為MT374548和MT374551。
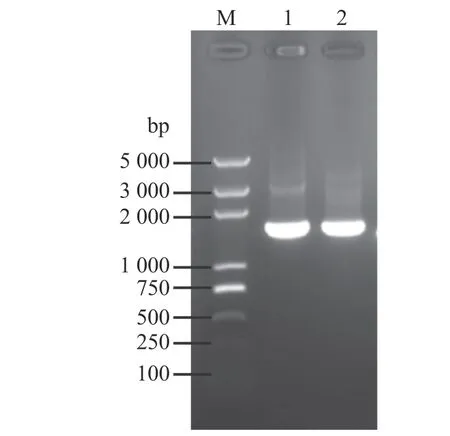
圖 1 澳洲堅果MiSTPP1和MiSTPP4基因的克隆Fig. 1 Isolations of MiSTPP1 and MiSTPP4 from M. integrifolia
2.2 MiSTPP1和MiSTPP4氨基酸序列的同源性分析
氨基酸多序列比對和功能結構域預測結果顯示,澳洲堅果MiSTPP1和MiSTPP4與其他植物來源的蛋白磷酸酶PP1家族成員的蛋白序列具有極高的相似度(圖2)。其中,MiSTPP1蛋白與水芙蓉NnPP1(XP_010259515)、歐洲油菜BnPP1(XP_013643938)、蕪菁BrPP1(XP_009151182)的氨基酸序列相似度分別為86.29%、83.72%和83.39%;MiSTPP4蛋白與雞血藤SsPP1(TKY61334)、鷹嘴豆CaPP1(XP_00449 5569)、蒺藜苜蓿MtPP1-1(AES61198)的氨基酸序列相似度分別為92.02%、91.72%和91.41%。這些蛋白都含有N端的絲氨酸/蘇氨酸特異性蛋白磷酸酶特征序列LRGNHE及蛋白磷酸酶PP1家族的典型結構域MPP_PP1_PPKL。
2.3 MiSTPP1和MiSTPP4的系統進化樹構建
絲氨酸/蘇氨酸磷酸酶(PP)家族的系統進化樹(圖3)顯示,PP家族分為PP1家族和PP2家族,其中,PP1家族成員又分成2個組,澳洲堅果MiSTPP1和MiSTPP4被分在不同的組內,其中MiSTPP1與水芙蓉NnPP1(XP_010259515)和罌粟PsPP1(XP_026450203)的親緣關系最近;MiSTPP4與橙子CsPP1(XP_006487724)和雞血藤SsPP1(TKY61334)的親緣關系最近,推測這些親緣關系近的PP1蛋白具有相同的進化起源及類似的結構和功能。
2.4 MiSTPP1和MiSTPP4蛋白的基本理化性質分析
ProtParam軟件分析MiSTPP1蛋白的結果表明,其相對分子質量為39.57 kDa,分子式C1 764H2 747N475O524S18,等電點(pI)為5.14,負電荷殘基數(Asp+Glu)為49個,正電荷殘基數(Arg+Lys)為36個,不穩定系數為50.03(為不穩定蛋白),脂肪族指數為90.54,親水性總平均值(GRAVY)為?0.274。因此,推測MiSTPP1蛋白是一個不穩定的親水性蛋白。

圖 2 MiSTPP1和MiSTPP4與其他植物PP1蛋白的同源性比對結果Fig. 2 Homology between MiSTPP1, MiSTPP4, and other PP1 proteins

圖 3 MiSTPP1和MiSTPP4與其他植物PP蛋白的系統進化關系Fig. 3 Relationship among phylogenetic trees of MiSTPP1, MiSTPP4, and other PP proteins
ProtParam軟件分析MiSTPP4蛋白的結果表明,其相對分子質量為36.74 kDa,分子式為C1 646H2 579N439O478S18,等電點(pI)為5.75,負電荷殘基數(Asp+Glu)為42個,正電荷殘基數(Arg+Lys)為37個,不穩定系數為36.77(為穩定蛋白),脂肪族指數為92.09,親水性總平均值(GRAVY)為?0.178。因此,推測M iSTPP4蛋白是一個穩定的親水性蛋白。
2.5 MiSTPP1和MiSTPP4蛋白的功能預測
在線程序NetPhos 3.1預測結果表明,MiSTPP1蛋白有23個磷酸化位點,其中絲氨酸(Serine)13個,蘇氨酸(Threonine)7個,酪氨酸(Tyrosine)3個;MiSTPP4蛋白有20個磷酸化位點,其中絲氨酸(Serine)8個,蘇氨酸(Threonine)9個,酪氨酸(Tyrosine)3個。PSORT軟件預測表明,MiSTPP1和MiSTPP4蛋白定位于細胞質可能性都為65%。SignalP和TMpred軟件預測顯示,MiSTPP1和MiSTPP4蛋白都不存在跨膜結構和信號肽,都為非分泌蛋白和非跨膜蛋白。
2.6 MiSTPP1和MiSTPP4蛋白的二級和三維結構預測
SOPMA軟件分析結果表明,澳洲堅果MiSTPP1多肽鏈中主要含有無規則卷曲(38.57%)和α-螺旋(36.57%),其次是折疊延伸鏈(18.29%)和β-轉角(6.57%)(圖4-A);MiSTPP4多肽鏈中主要含有α-螺旋(42.02%)和無規則卷曲(36.50%),其次是折疊延伸鏈(14.42%)和β-轉角(7.06%)(圖4-B)。
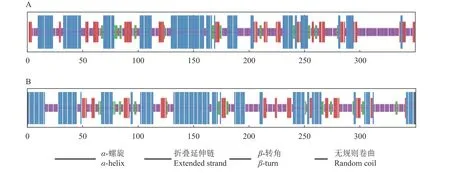
圖 4 MiSTPP1蛋白(A)和MiSTPP4蛋白(B)的二級結構分析Fig. 4 Secondary structure of MiSTPP1 (A) and MiSTPP4 (B)
以絲氨酸/蘇氨酸磷酸酶PP1的晶體結構(PDB ID:1s70.1.A)為同源模板,利用Swiss-model軟件構建MiSTPP1蛋白的三維預測模型。結果表明,MiSTPP1蛋白的氨基酸序列與模板序列一致性為69.55%,GMQE值為0.78,其核心結構主要由無規卷曲和α-螺旋組成(圖5-A)。同樣,以絲氨酸/蘇氨酸磷酸酶PP1的晶體結構(PDB ID:6dcx.2.A)為同源模板,利用Swiss-model軟件構建MiSTPP4蛋白的三維預測模型。結果表明,MiSTPP4蛋白的氨基酸序列與模板序列一致性為76.42%,GMQE值為0.83,其核心結構主要由α-螺旋和無規卷曲組成(圖5-B)。MiSTPP1和MiSTPP4蛋白的三維模型組分與其二級結構的預測結果相符合。

圖 5 MiSTPP1蛋白(A)和MiSTPP4蛋白(B)的三維結構分析Fig. 5 3D configurations of MiSTPP1 (A) and MiSTPP4 (B)
絲氨酸/蘇氨酸磷酸酶PP1家族成員在細胞信號轉導、新陳代謝、生長與凋亡等多種細胞生理生化過程中廣泛表達,且其蛋白序列與結構在長期進化中也高度保守[13]。本研究利用本課題組澳洲堅果抗寒轉錄組數據信息,通過RT-PCR技術成功獲得了屬于PP1基因家族的澳洲堅果MiSTPP1和MiSTPP4基因cDNA全長。蛋白序列和系統進化樹分析表明,MiSTPP1和MiSTPP4蛋白與其他植物PP1蛋白具有極高的序列相似度,都屬于PP1進化分支,都含有PP1家族的典型結構域MPP_PP1_PPKL,尤其是N端都含有絲氨酸/蘇氨酸特異性蛋白磷酸酶特征序列LRGNHE。這些為MiSTPP1和MiSTPP4蛋白的結構與功能研究提供氨基酸序列基礎。
蛋白基本理化性質分析表明,MiSTPP1蛋白是一個不穩定的親水性蛋白,而MiSTPP4蛋白是一個穩定的親水性蛋白。蛋白磷酸化位點分析表明,MiSTPP1蛋白以絲氨酸磷酸化為主,而MiSTPP4蛋白以絲氨酸和蘇氨酸磷酸化為主,結合磷酸化修飾在蛋白功能中的作用[14?15],推測這兩類氨基酸進行磷酸化修飾時在各自蛋白行駛功能中發揮重要的作用。蛋白亞細胞定位、跨膜結構和信號肽預測表明,MiSTPP1和MiSTPP4蛋白極可能定位于細胞質,推測這兩個蛋白作為非分泌蛋白或非跨膜蛋白行駛其生物學功能。蛋白二級結構和三維結構分析表明,MiSTPP1和MiSTPP4蛋白的二級和三級結構主要由α-螺旋和無規卷曲組成,并夾雜著折疊延伸鏈,尤其是預測的三維結構模型與PP1家族已知的晶體結構具有極高的一致性(分別達到69.55%和76.42%)。說明PP1蛋白在長期的物種進化過程中,MiSTPP1和MiSTPP4蛋白同其他PP1家族成員一樣,其蛋白的空間結構高度保守,推測其有類似的生物學功能。
鑒于MiSTPP1和MiSTPP4蛋白與其他功能已知的PP1家族成員具有極高的親緣關系和空間結構相似性,可以初步推測MiSTPP1和MiSTPP4基因在響應逆境脅迫、信號轉導、生長發育等生理生化過程中發揮重要的作用。但是,其具體的功能還需要通過亞細胞定位、酵母自激活及遺傳轉化等技術進行深入研究,從而為闡明澳洲堅果PP1蛋白的分子調控機制提供科學依據。
