基于SNP標記的小麥籽粒性狀全基因組關聯分析
2021-05-31 07:44:34張芳任毅曹俊梅李法計夏先春耿洪偉
中國農業科學 2021年10期
張芳,任毅,曹俊梅,李法計,夏先春,耿洪偉?
1新疆農業大學農學院/農業生物技術重點實驗室,烏魯木齊 830052;2新疆農業科學院糧食作物研究所,烏魯木齊 830091;3山東省農業科學院作物研究所/小麥玉米國家工程實驗室/農業部黃淮北部小麥生物學與遺傳育種重點實驗室,濟南 250100;4中國農業科學院作物科學研究所/國家小麥改良中心,北京 100081
0 引言
【研究意義】小麥(TriticumaestivumL.)是全球播種面積最廣的糧食作物之一,也是中國重要的糧食作物之一,在糧食安全中具有舉足輕重的地位[1]。目前,小麥的需求增長速度逐漸快于產量的提高速度,使得糧食安全面臨嚴峻考驗。為此,在耕地面積有限的現狀下,提高小麥單產至關重要[2]。小麥籽粒大小決定粒重高低,其中籽粒的長、寬、面積、周長、長寬比等參數是構成小麥產量的重要指標[3]。基于此,研究小麥籽粒性狀的遺傳特性對提高小麥產量及品質至關重要。【前人研究進展】高產一直是育種家最為關注的育種目標之一,而籽粒性狀的遺傳改良對產量潛力的提升具有顯著影響[4-6]。隨著分子標記技術的快速發展,關聯分析被應用于數量性狀的遺傳研究,作為基因發掘的重要手段,為分子標記輔助選擇提供了重要依據[7]。SNP標記具有遺傳穩定性高、位點豐富且分布廣等特點,其密度遠遠高于傳統的分子標記,對于數量性狀定位更為高效和便捷[8-10]。目前,小麥相繼開發出9K、90K、660K、15K和50K等滿足不同育種需求的SNP芯片[11-13]。小麥籽粒性狀遺傳結構復雜,受微效多基因控制,國內外已有大量學者對小麥籽粒相關性狀進行了遺傳研究,并對調控小麥籽粒大小性狀的QTL區段進行了定位和遺傳解析。小麥籽粒性狀已挖掘到QTL位點主要位于1A、1B、1D、2A、2B、2D、3B、4B、5A、6B、7B和7D等染色體[14-16]。LI等[17]利用 W7984×Opata85重組自交系進行連鎖分析,在2個環境下均發現7D染色上控制粒長的主效 QTL,可解釋 13.5%—13.9%的表型變異。全基因組關聯分析(genome-wide association study,GWAS)作為分析復雜性狀遺傳結構的有力工具,在水稻、玉米和擬南芥中得到了廣泛的應用[18-21]。BHATTA等[22]以143份面包小麥品種為材料,采用GWAS關聯分析,在1B、1D、2A、2B、3A、3B、4A、4B、4D、5A、5B、5D、6D、7A、7B染色體上共檢測到 38個籽粒性狀相關位點,其貢獻率為0.3%—19.6%。SUN等[23]利用90K SNP芯片對163份小麥主載品種(系)進行關聯分析,在1A、3A、3D、5B、5D、6A、6B和7A染色體上檢測到多個千粒重相關位點,貢獻率為15.7%—31.6%。DABA等[24]利用200份冬小麥群體在2個環境下對粒重和粒長進行關聯分析,檢測到26個粒重和27個粒長顯著關聯的標記。LI等[25]采用SNP-GWAS方法對166份黃淮麥區小麥品種進行產量相關性狀分析,分別挖掘出 20個粒長和 31個粒寬顯著關聯的標記,貢獻率分別為7.1%—14.2%和6.8%—15.0%,其中位于 2A染色體的IWB32119標記與粒重基因TaFlo2-A1[26]的位點一致。【本研究切入點】新疆作為中國主要的小麥產區之一,對產量的提升至關重要。然而,該地區小麥生長關鍵階段常面臨干旱少雨和干熱風等不利環境的影響,常規育種手段依舊有效,但靠其進行單產的提升,跟不上生產需求,需對分子標記等新技術進行充分使用,但基于高密度芯片技術對新疆小麥籽粒性狀的全基因組關聯分析鮮有報道。【擬解決的關鍵問題】本研究以121份新疆種植的小麥品種(系)為材料,結合50K SNP芯片對千粒重、粒長、粒寬、籽粒長寬比、籽粒面積和籽粒周長等6個籽粒性狀進行全基因組關聯分析,以期獲得新的與籽粒性狀關聯的SNP位點,為小麥聚合育種及復雜數量性狀位點的研究提供參考依據。
1 材料與方法
1.1 試驗材料及田間種植
選用121份新疆種植小麥品種(系),其中引進品種(系)36份,分別來自前蘇聯、烏克蘭、羅馬尼亞、北京和河北;自育品種(系)68份;地方品種(系)17份。上述材料由新疆農業大學農學院小麥遺傳育種課題組提供。
參試材料于 2016—2017、2017—2018和 2018—2019年度種植于新疆農業科學院瑪納斯農業試驗站,隨機區組設計,三行區,行長2 m,行距25 cm,3次重復。田間管理遵循當地基本栽培管理方式。待小麥生理成熟期后,按每品種取10株收獲脫粒,在自然曬干后均通過杭州萬深 SC-G拷種儀器檢測千粒重(thousand kernel weight,TKW)、粒長(grain Length,GL)、粒寬(grain width,GW)、粒長寬比(length width ratio,LWR)、籽粒面積(grain area,GA)、籽粒周長(grain perimeter,GC)。利用 IciMapping的ANOVA功能進行田間數據分析及遺傳力的估算。廣義遺傳力(h2),計算公式:h2=σg2/(σg2+σe2),σg2為遺傳方差,σe2為環境方差。
1.2 SNP標記檢測
50K標記利用Illumina SNP Genotyping技術測試平臺(北京博奧生物有限公司),使用微珠芯片技術(Bead Array)進行檢測,利用Genomestudio v1.0軟件進行多態性分析。獲得的基因型數據進行質量控制,刪除缺失率>20%的標記,過濾掉最小等位基因頻率(minor allele frequency,MAF)<5%的標記。
1.3 群體結構分析
采用PowerMarker V3.25軟件對參試材料進行遺傳多樣性和多態性信息含量分析。其中,PIC=1-∑(Pij)2,Pij表示i位點的第j個等位變異出現的頻率。
選取在小麥21條染色體上均勻分布的2 000個SNP標記,利用Structure 2.3.4軟件進行群體結構分析。參數設置:Burn-in period為10 000,MCMC為100 000,選擇Admixed模型,設K=2—12,每個K重復5次,結果上傳Structure Harvester,獲得最佳K值作為亞群數目。
1.4 關聯分析
應用Tassel V5.0中的混合線性模型(mixed linear model,MLM)對SNP標記與性狀進行關聯分析,并利用Structure 2.3.4軟件計算Q值,結合Tassel V5.0軟件計算Kinship值。在P<0.001水平下認為標記與性狀存在關聯,以第95百分位的r2值作為閾值估測LD衰減距離。
2 結果
2.1 表型分析
通過對千粒重、粒長、粒寬、籽粒長寬比、籽粒面積和籽粒周長等6個籽粒相關性狀的表型進行統計分析(表 1),結果表明,不同環境間各性狀均表現較大差異,其中,千粒重變異系數最大。各指標的峰度和偏度均接近1,說明均符合正態分布,是多基因控制的數量性狀。方差分析(表2)表明,各性狀在基因型、環境及基因型與環境互作間均呈現出極顯著差異(P<0.001)。6個籽粒相關性狀的廣義遺傳力(h2)為0.85—0.90,其中,長寬比和粒寬的遺傳力較高,分別達到0.90和0.89,表明小麥籽粒性狀主要受遺傳變異影響。相關性分析(表3)表明,粒長與籽粒周長相關性最大(r=0.94),其次為粒寬與千粒重相關性最好(r=0.93);長寬比與千粒重、粒寬和籽粒面積呈極顯著負相關(P<0.001),進一步說明小麥籽粒性狀間存在協同變化關系。
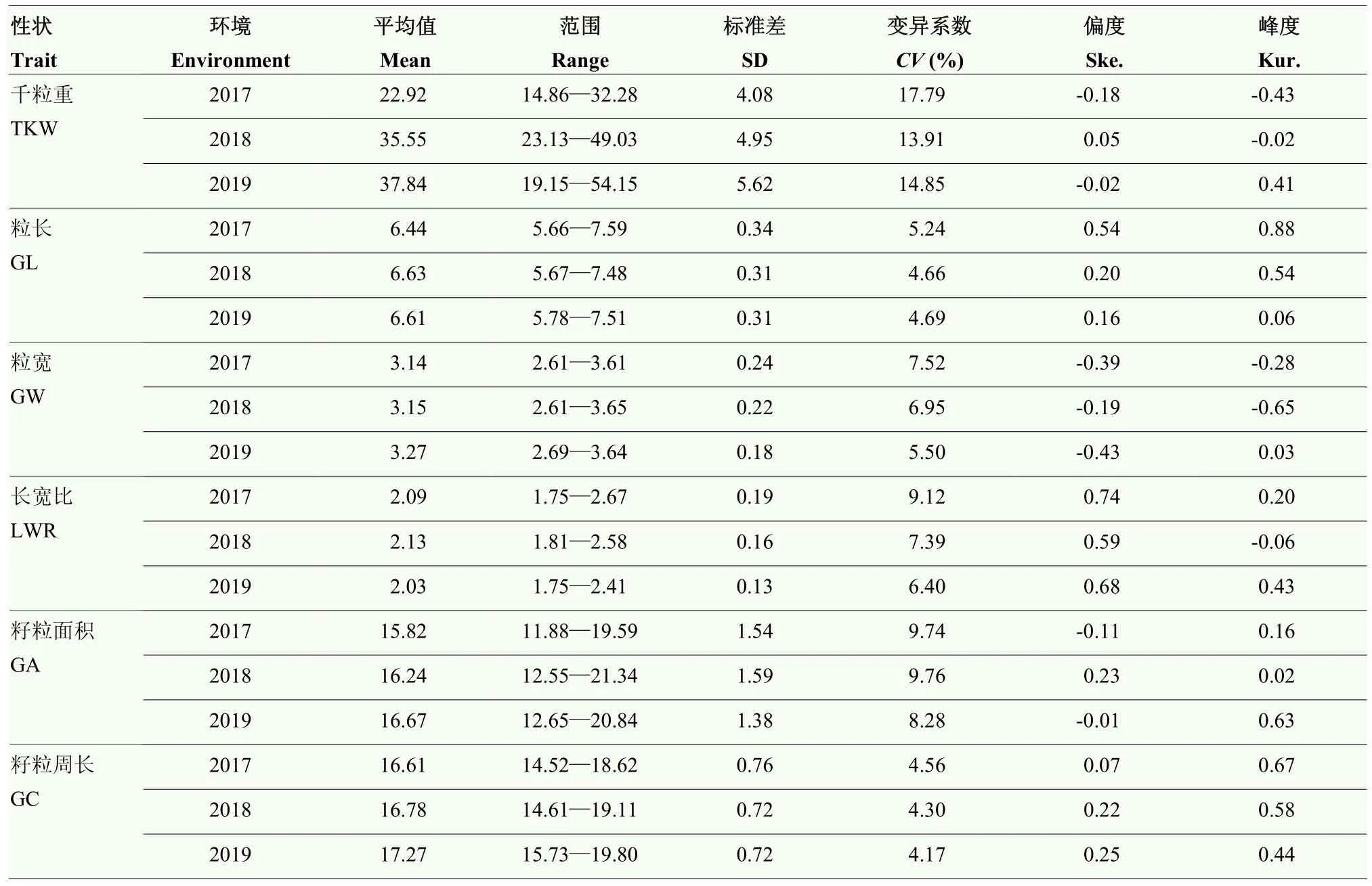
表1 小麥自然群體籽粒性狀基本統計信息Table 1 Phenotypic data analysis of grain size related traits in the wheat nature population
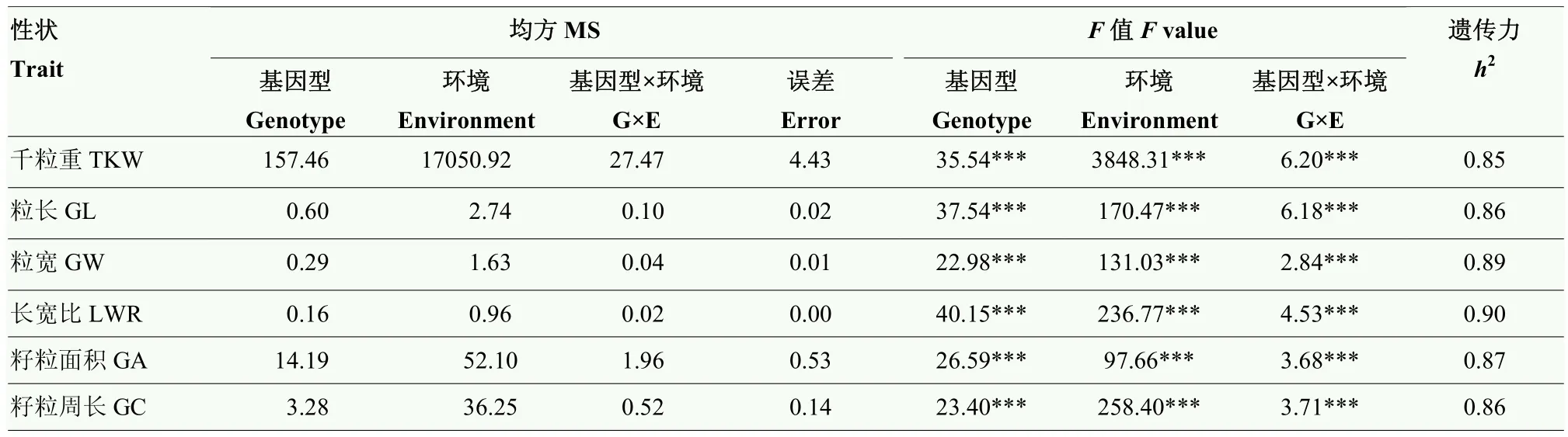
表2 小麥籽粒性狀方差分析Table 2 Analysis of variance of grain size related traits in the wheat
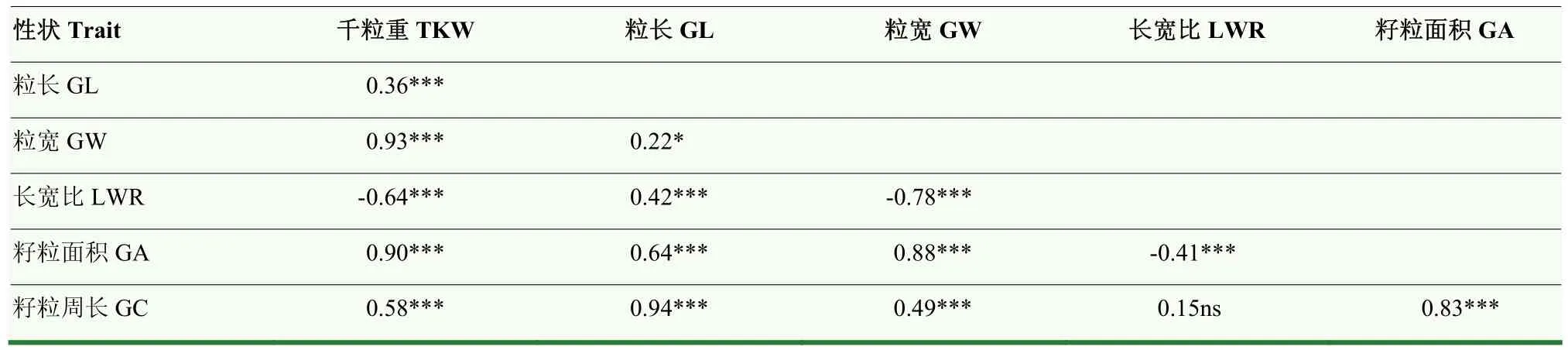
表3 小麥籽粒性狀間相關性分析Table 3 Correlation analysis of grain size related traits in the wheat
2.2 標記遺傳多樣性及群體結構
利用50K SNP芯片對121份新疆種植的小麥品種(系)進行檢測,經過質量控制最終篩選出36 873個SNP標記進行GWAS分析,其中,A基因組、B基因組和D基因組染色體平均包含16 514、11 490和8 869個SNP標記,每個標記間的平均距離為0.39 Mb(電子附表1)。A與B基因組的染色體組所用標記密度、標記數目、遺傳多樣性和多態信息含量均高于D染色體組。SNP標記的PIC值為0.09—0.38,平均值為0.32。
利用Structure 2.3.4軟件分析,獲得121份新疆種植的小麥自然群體結構Q值與極大似然值LnP(D),進一步確定最佳K值。當K=4時(圖1-A),ΔK值最大,由圖1-B可知,參試材料被劃分為4個亞群(subpopulation),其中,亞群Ⅰ(SubgroupⅠ)包含46份品種(系),主要來自自育和引進品種(系),分別占比47.83%和41.30%,引進品種多為前蘇聯品種(系);亞群Ⅱ(SubgroupⅡ)包含29份品種(系),主要來自自育品種(系),占比79.31%;亞群Ⅲ(SubgroupⅢ)包含41份品種(系),包含56.10%的自育品種(系)、26.83%的引進品種(系)和17.07%的地方品種(系);亞群Ⅳ(SubgroupⅣ)包含5份品種(系),均來自地方品種(系);4個亞群所含品種數分別占總材料份數的38.02%、23.97%、33.88%和4.13%。
2.3 籽粒相關性狀關聯分析
基于 LD衰減距離計算,將在物理圖譜上前后 3 Mb區間內的位點認定為一個候選位點。用36 873個分子標記對3個環境及平均環境下的表型數據進行關聯分析(圖2),6個小麥籽粒相關性狀共檢測到592個顯著SNP位點(P<0.001),單個關聯標記位點解釋的表型變異(R2)范圍為9.3%—28.6%。其中29個SNP至少在2個環境中被檢測到,分布在1A(5)、1B(2)、1D、2A(5)、3B、5A、5D、6B(4)、6D、7B和 7D(7)染色體上,可解釋 9.3%—22.7%的表型變異(表4)。檢測到6個與千粒重穩定關聯的標記,分布在1B、2A和6B染色體上,其中在6B染色體檢測到 4個千粒重位點(AX-109874065、AX-110446017、AX-110936500和AX-109493716),且效應值較大,介于9.7%—12.9%,說明 6B染色體對千粒重影響較大,可作為籽粒性狀重要的染色體。檢測到6個與粒長穩定關聯的標記,分布在1A、2A和7D染色體上,其中AX-111082947和AX-94497666在3個環境和均值下同時被檢測到,表型變異解釋范圍為9.9%—22.7%。檢測到2個與粒寬穩定關聯的標記,分布在 3B和 5D染色體上的AX-108948870與AX-179477405,表型變異解釋范圍為9.6%—12.2%。檢測到6個與長寬比穩定關聯的標記,分布在2A(2)、5A、7B和7D(2)染色體上,其中位于2A染色體的AX-111722425標記在全部環境下均被重復檢測到,可解釋10.1%—18.3%表型變異。檢測到3個與籽粒面積穩定關聯的標記,分布在1A、1B和1D染色體上,表型變異解釋率范圍為9.9%—18.2%,表明1號同源群對籽粒面積具有重要作用。檢測到6個與籽粒周長穩定關聯的標記,分布在1A(2)、2A、6D和7D(2)染色體上,其中,位于1A染色體的AX-111082947標記在多環境下同時被檢測到,且貢獻率最大,可解釋10.1%—22.6%表型變異。
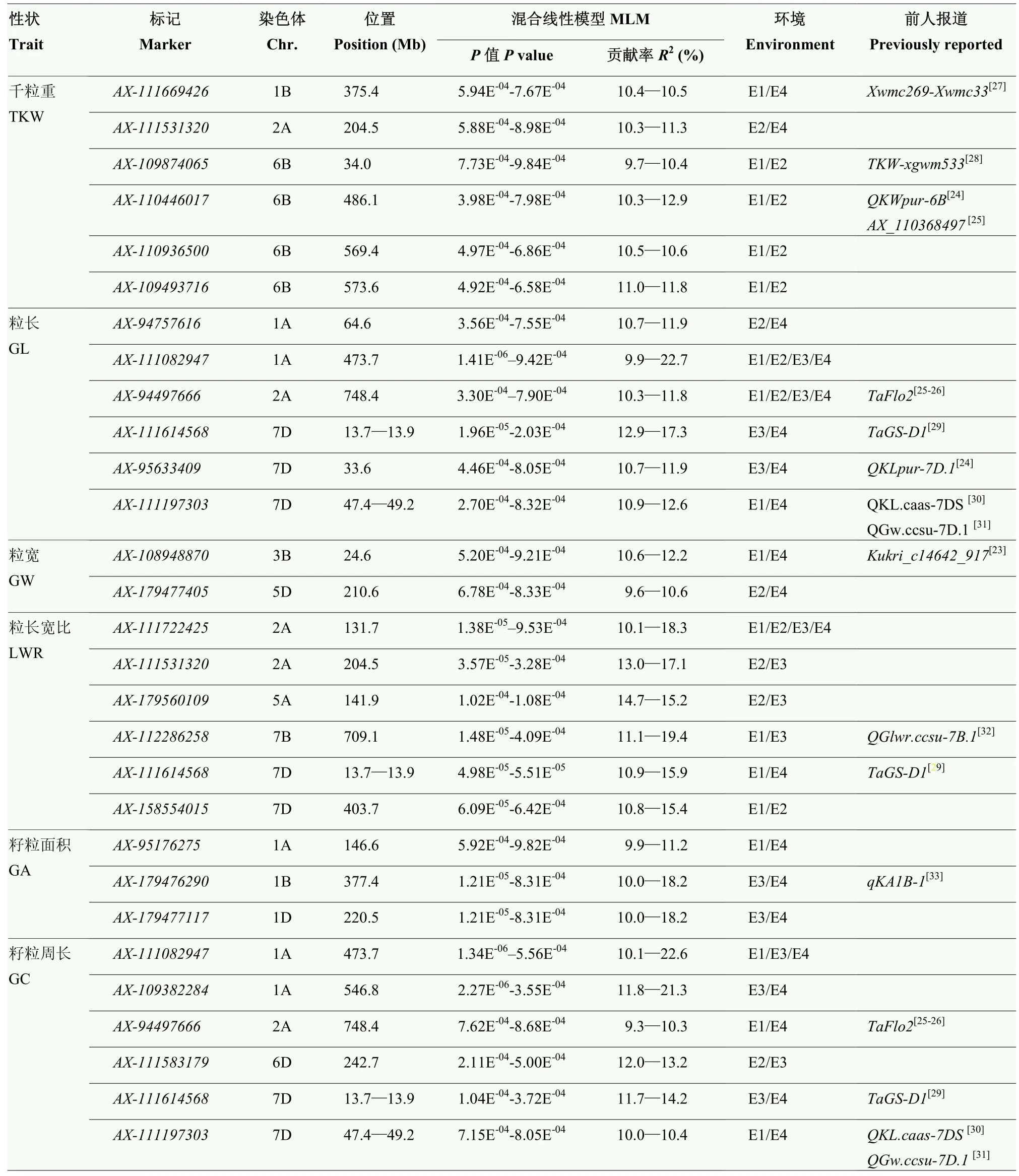
表4 SNP-GWAS檢測到的產量性狀相關位點Table 4 Loci for grain size related traits in the diversity panel identified by SNP-GWAS
29個穩定關聯的位點中,位于 1A染色體上的AX-111082947和位于2A染色體上的AX-94497666標記均與粒長和籽粒周長顯著關聯,貢獻率分別為9.3%—22.7%和 9.3%—11.8%;位于 2A染色體上的AX-111531320標記均與千粒重和籽粒長寬比顯著關聯,貢獻率分別為 10.3%—17.1%;位于 7D染色體上的AX-111614568標記同時與粒長、籽粒周長和籽粒長寬比顯著關聯,貢獻率為10.9%—17.3%;位于7D染色體上的標記AX-111197303同時與粒長和籽粒周長顯著關聯,貢獻率為10.0%—12.6%,說明這5個位點是一因多效位點。
3 討論
3.1 小麥籽粒性狀表型
小麥籽粒性狀包括粒長、粒寬、長寬比、粒厚、籽粒面積、籽粒周長和千粒重等指標,其高低是衡量產量的重要依據之一,且性狀便于觀察,作為小麥育種選擇最直觀的性狀,往往受到研究者的高度關注[34]。DHOLAKIA 等[35]認為小麥籽粒性狀不但對產量具有影響,也決定小麥的商品價值。粒重受粒長、粒寬、粒厚以及籽粒周長等方面的直接影響[36],其中粒寬與粒重呈顯著正相關,相關性較大。陳佳慧等[37]、余曼麗等[38]發現千粒重與粒長、粒寬、籽粒面積、籽粒周長均呈極顯著正相關,與籽粒長寬比呈負相關,這與本研究結果基本一致。此外發現粒長與籽粒周長、粒寬與籽粒面積存在協同促進關系,說明各性狀在生長發育過程中由單個基因多效性引起或多個基因緊密連鎖共同控制某些性狀來作為性狀相關的遺傳基礎。不同類型新疆種植的小麥材料中,自育品種的大粒表現較高,地方品種次之,引進品種較低,這一結果與馬艷明等[39]基本一致,充分體現了新疆自育品種間籽粒性狀差異較豐富。究其原因一方面是自育品種經過育種家多年以大粒選擇為主有關,其重視度和選擇度較強;另一方面是結合當地環境的育種需求,大粒小麥品種對小麥粒重遺傳改良具有較大的提升空間,將更有利于中國小麥高產優質新品種培育。另外,本研究還發現籽粒長寬比地方品種顯著高于自育品種和引進品種,這可能由于地方品種一般為長而尖粒型,其粒長較粒寬發育較早,而育成品種在選育過程中,著重于粒長粒寬進行改良,呈現出粒寬的增加同時粒長未發生明顯變化[39]。這一結果充分體現出千粒重提高主要由粒寬的增加而影響,而小麥產量的提升主要由千粒重來貢獻。
3.2 小麥籽粒性狀GWAS分析
在多個環境下均能被檢測到的關聯位點可視為相對穩定的位點,本研究共檢測到29個籽粒性狀穩定關聯的SNP位點。粒長穩定位點分布于1A、2A和7D染色體上,LI等[25]通過166份黃淮麥區材料結合90K SNP芯片進行關聯分析,在2A染色體上發現一個穩定位點,并確定為粒重基因TaFlo2-A1的位點,這與本研究在2A染色體檢測到的標記AX-94497666所在位點相距僅約3.4 Mb,可能為相同位點,該位點控制的粒長效應可能來自于該基因的作用。同時位于 7D染色體上的標記AX-111614568檢測到一個粒長相關位點,這與粒重基因TaGS-D1位點一致,也為相同位點[29]。粒長標記AX-95633409與 DABA等[24]發現位點QKLpur-7D.1較近,相距0.5 Mb,LI等[30]與MIR等[31]分別也在7D染色體上檢測到標記QKL.caas-7DS和QGw.ccsu-7D.1與本研究粒長標記AX-111197303均位于短臂區域。本研究在 3B染色體上發現的控制粒寬的SNP標記AX-108948870與王瑞霞等[27]、SUN等[23]報道的粒寬QTL位置相似,其表型貢獻率較大,CABRAL等[40]也證實了在3B染色體上相近的位置上發現了一個QTL位點。位于7B染色體上發現籽粒長寬比標記AX-112286258貢獻率最大,KUMARI等[32]檢測到的 QTL位點相距 31.6 Mb。XIN 等[33]利用Dalibao×BYL8 RIL進行籽粒性狀 QTL定位,獲得qKA1B-1籽粒面積位點與本研究發現的標記AX-179476290位置相近。對于千粒重,在6B染色體上鑒定了 4個穩定顯著 SNP標記(AX-109874065、AX-110446017、AX-110936500和AX-109493716),其貢獻率為9.7%—12.9%。其中,AX-109874065標記與張坤普[28]報告的千粒重的位點TKW-xgwm533僅為1.3 Mb,因此,筆者認為上述屬于同一位點控制千粒重高低。DABA等[24]和LI等[25]分別檢測到QKWpur-6B和AX_110368497與本研究標記AX-110446017相距31.7和31.9 Mb,說明該染色體上存在粒重密切相關位點。MUHAMMAD等[41]也在6B染色體檢測到控制千粒重的位點,其貢獻率為12.0%—14.0%,后期將對該染色體的重要位點進行SNP標記開發,為分子標記輔助選擇提供有效的選擇標記。
本研究在位于1A、1B、2A、5A、5D、6B和7D染色體上發現可能與籽粒相關性狀穩定遺傳的新位點。其中,在 2A染色體上發現控制千粒重的新位點SNP標記AX-111531320,同時也控制籽粒長寬比,其效應值大于10%且穩定。共位點區域可能存在著控制不同性狀的多個基因,或者是存在一個基因控制多個性狀[42]。位于1A染色體上標記AX-111082947被發現是新位點且貢獻率最大,為9.8%—22.7%,同時影響粒長和籽粒周長,有可能為控制小麥粒重主效位點,該基因可在小麥產量育種中起到重要的作用,可優選1A染色體關聯位點進行功能標記開發,為分子標記輔助育種提供有效信息。在 7D染色體上接觸到的位點AX-111197303標記,也可同時影響粒長和籽粒周長,表明這兩個位點控制粒長和籽粒周長的基因可能為同一基因且驗證了粒長與籽粒周長具有協同作用。后期研究可聚焦這些多效應位點且在多環境下均被檢測到的穩定位點,進一步開展精細定位和分子標記開發等研究。
4 結論
本研究材料遺傳多樣性豐富,在2個及以上環境中共檢測到29個與6個籽粒性狀穩定顯著關聯位點,其中標記AX-94497666和AX-111614568與粒長和籽粒周長顯著關聯,與前人發現的TaFlo2-A1和TaGS-D1相吻合。檢測到5個顯著位點為多效性位點,貢獻率為9.9%—22.7%。標記AX-111082947(1A)和AX-111197303(7D)與粒長和籽粒周長顯著關聯,貢獻率分別為9.8%—22.7%和10.0%—12.6%。
猜你喜歡
新世紀智能(數學備考)(2021年9期)2021-11-24 01:14:30
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
當代陜西(2021年17期)2021-11-06 03:21:36
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
當代陜西(2019年15期)2019-09-02 01:52:00
學苑創造·A版(2018年11期)2018-02-01 06:29:20
讀者(2017年5期)2017-02-15 18:04:18
