miR-140-5p對炎癥模型支氣管上皮細胞炎癥因子分泌的影響
2021-06-08 07:37:52劉彩林
鄭州大學學報(醫學版) 2021年3期
張 俊,馬 冰,劉彩林
1)駐馬店市中心醫院檢驗科 河南駐馬店 463000 2)河南省人民醫院檢驗科 鄭州 450000 3)鄭州大學第一附屬醫院檢驗科 鄭州 450052
miRNA是一種單鏈RNA分子,在多種生物體中廣泛存在,其不僅參與調控細胞分化、衰老、免疫反應等正常生理過程,還與人類疾病的發生關系密切[1]。炎癥與多種miRNA的異常表達有關,這些miRNA通過調控下游靶基因的表達影響靶細胞炎癥因子的合成和分泌[2]。miR-140-5p是近些年來發現的具有多種生物學作用的miRNA,在腫瘤、關節炎等疾病的發生發展過程中發揮作用[3-4]。以前的研究[5]發現,miR-140-5p在支氣管哮喘模型小鼠肺組織中表達下調。呼吸道合胞病毒(respiratory syncytial virus,RSV)是一種常見的病原體,其可以誘導肺炎、毛細支氣管炎發生,RSV刺激可促使支氣管上皮細胞釋放大量炎癥因子,這也是RSV加重哮喘的重要原因[6-7]。本實驗以RSV感染人支氣管上皮細胞16-HBE,構建支氣管上皮細胞炎癥模型,探討miR-140-5p在支氣管上皮細胞炎癥反應中的作用和機制。
1 材料與方法
1.1細胞、主要試劑和儀器16-HBE購自上海康朗生物科技有限公司。RSV病毒Long株購自美國ATCC。Lipofectamine2000購自美國Invitrogen公司,miR-140-5p陰性對照(miR-NC)及其mimic購自百奧邁科生物技術有限公司,Toll樣受體4(Toll-like receptor 4,TLR4)過表達載體及其陰性對照載體由北京信諾金達生物科技有限公司構建。雙熒光素酶報告實驗試劑盒購自上海碧云天生物技術研究所。Trizol試劑、反轉錄試劑盒、Real-time PCR 試劑盒購自賽默飛世爾科技有限公司。IL-6、IL-8含量檢測試劑盒購自北京百奧萊博科技有限公司,IL-1β含量檢測試劑盒購自武漢艾美捷科技有限公司,NO含量檢測試劑盒購自南京碧云天生物技術研究所,TNF-α含量檢測試劑盒購自南京森貝伽生物科技有限公司。iNOS、TLR4抗體購自美國Abgent公司。酶標儀和熒光定量PCR儀購自美國Thermo公司;凝膠成像系統購自英國Syngene公司;全自動激光共聚焦顯微鏡購自日本Olympus公司。
1.2miR-140-5p與TLR4靶向關系的預測和鑒定利用生物信息學軟件TargetScan分析miR-140-5p的靶基因,發現miR-140-5p和TLR4的3’UTR有互補結合位點,利用雙熒光素酶報告實驗鑒定二者的靶向關系。將含有野生型(WT)或突變型(MUT)TLR4 3’UTR結合位點的熒光素酶報告載體分別與miR-NC、miR-140-5p mimic共轉染到16-HBE細胞中,培養24 h以后,測定熒光素酶活性。
1.3上調miR-140-5p表達對炎癥模型16-HBE細胞炎癥因子分泌和NO合成的影響
1.3.1 實驗分組 將16-HBE細胞分成空白對照組、RSV組、miR-NC+RSV組、miR-140-5p+RSV組,RSV組、miR-NC+RSV組、miR-140-5p+RSV組細胞感染RSV(MOI=1)24 h,以制備炎癥模型。然后miR-NC+RSV組、miR-140-5p+RSV組分別轉染miR-NC、miR-140-5p mimic,按照Lipofectamine2000說明操作。轉染24 h后進行指標檢測。實驗均重復3次。
1.3.2 Real-time PCR法檢測細胞中miR-140-5p的表達 分別收集4組細胞,每107個細胞內添加1 mL Trizol試劑以提取細胞總RNA。RNA經紫外分光光度計測定,OD(260 nm)/OD(280 nm)比值介于1.8~2.0,可以用于后續實驗。取1 μg總RNA,添加1 μL的Oligo dT Primer,加RNase水補足12 μL,置于85 ℃結合5 min,冰上冷卻,然后添加1 μL的RNase抑制劑、1 μL的dNTP、4 μL的5×緩沖液、1 μL的RT,最后添加RNase水補足20 μL。反轉錄條件:30 ℃10 min,42 ℃50 min,85 ℃10 min。合成的cDNA保存在-20 ℃冰箱中。取100 ng的cDNA,加入0.4 μL的ROX reference Dye Ⅱ, 上下游引物各0.4 μL, 10 μL的SYBR Premix EX Taq,最后加ddH2O補足20 μL。PCR反應條件:預變性95 ℃ 30 s,變性95 ℃ 5 s,退火延伸60 ℃ 34 s,共40個循環。按照2-ΔΔCt法分析miR-140-5p表達水平。miR-140-5p上游引物序列為5’-GAGTGTCAGTG GTTTTACCCT-3’,下游為5’-GCAGGGTCCGAGG TATTC-3’;內參U6上游引物序列為5’-CTCGCT TCGGCAGCACATATACTA-3’,下游為5’-ACGAATTTGCGTGTCATCCTTGCG-3’。
1.3.3 ELISA法檢測細胞TNF-α、IL-1β、IL-6、IL-8分泌量 收集4組細胞培養上清,ELISA法測定TNF-α、IL-1β、IL-6、IL-8含量,按試劑盒說明操作。
1.3.4 Griess法檢測NO生成量 收集4組細胞培養上清,Griess法測定NO含量,按試劑盒說明操作。
1.3.5 Western blot法檢測細胞中iNOS和TLR4蛋白的表達 收集4組細胞, RIPA裂解,低溫高速離心10 min,吸取上清,以BCA法檢測蛋白濃度。將蛋白樣品和等體積的Loading Buffer充分混合后,按照每孔40 μg蛋白樣品上樣,SDS-PAGE(100 g/L分離膠、50 g/L濃縮膠)進行蛋白分離。將PVDF膜浸泡在轉移緩沖液中孵育5 min,在50 g/L脫脂奶粉溶液中室溫搖床孵育1 h,再分別置于iNOS和TLR4一抗稀釋液(均按1∶800稀釋)中于4 ℃冰箱中搖床孵育12 h, 然后置于二抗稀釋液(按1∶3 000稀釋)中室溫搖床孵育1 h,ECL發光,Image J分析。以目的蛋白和內參GAPDH條帶灰度值的比值表示目的蛋白表達水平。
1.4過表達TLR4對上調miR-140-5p表達的炎癥模型16-HBE細胞炎癥因子分泌和NO合成的影響將TLR4陰性對照載體和miR-140-5p mimic(miR-140-5p組)、TLR4過表達載體和miR-140-5p mimic(miR-140-5p+ TLR4)分別共轉染到16-HBE細胞中,培養24 h后,感染RSV(MOI=1)繼續孵育24 h。檢測培養上清中TNF-α、IL-1β、IL-6、IL-8、NO含量及細胞中iNOS、TLR4蛋白表達水平,操作同上,均重復3次。
1.5統計學處理實驗數據用SPSS 21.0分析。空白對照組、RSV組、miR-NC+RSV組、miR-140-5p+RSV組間miR-140-5p表達水平,TNF-α、IL-1β、IL-6、IL-8分泌量,NO生成量,iNOS和TLR4蛋白表達水平的比較均采用單因素方差分析,組間兩兩比較采用SNK-q檢驗;miR-140-5p組、miR-140-5p+ TLR4組間各指標的比較采用兩獨立樣本t檢驗;檢驗水準α=0.05。
2 結果
2.1miR-140-5p與TLR4靶向關系的驗證生物信息學分析發現miR-140-5p和TLR4的3’UTR有互補結合位點(圖1)。雙熒光素酶報告實驗結果顯示,miR-140-5p mimic和TLR4-3’UTR-WT共轉染的細胞熒光素酶活性降低(表1)。

圖1 miR-140-5p和TLR4結合位點示意圖

表1 熒光素酶活性(n=3)
2.2上調miR-140-5p表達后炎癥模型細胞中miR-140-5p和TRL4蛋白表達水平的變化空白對照組、RSV組、miR-NC+RSV組、miR-140-5p+RSV組細胞中miR-140-5p和TRL4蛋白表達檢測結果見圖2、表2。可以看出,miR-140-5p mimic轉染可有效提高miR-140-5p的表達;炎癥模型細胞中miR-140-5p表達水平降低;miR-140-5p可負調控0炎癥模型細胞中TRL4蛋白的表達。

1:空白對照組;2:RSV組;3:miR-NC+RSV組;4:miR-140-5p+RSV組

表2 4組細胞中miR-140-5p和TRL4蛋白的表達水平(n=3)
2.3上調miR-140-5p表達后炎癥模型細胞炎癥因子分泌和NO合成能力的變化空白對照組、RSV組、miR-NC+RSV組、miR-140-5p+RSV組細胞中炎癥因子分泌量、NO生成量和細胞中iNOS蛋白表達水平的比較見圖3、表3。可以看出,上調miR-140-5p表達可以降低炎癥模型細胞炎癥因子分泌和NO合成的能力。
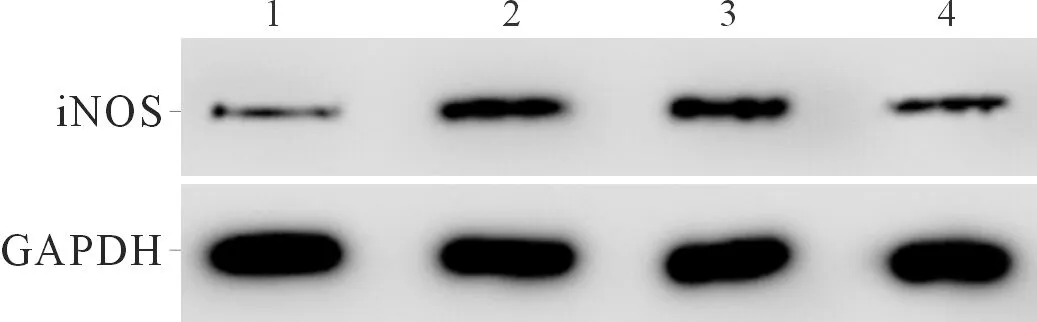
1:空白對照組;2:RSV組;3:miR-NC+RSV組;4:miR-140-5p+RSV組

表3 4組細胞TNF-α、IL-1β、IL-6、IL-8分泌量,NO生成量及細胞中iNOS蛋白表達水平的比較(n=3)
2.4過表達TLR4后miR-140-5p表達上調的炎癥模型細胞炎癥因子分泌和NO合成能力的變化miR-140-5p組、miR-140-5p+TLR4組各指標測定結果見圖4、表4。可以看出,TLR4可逆轉miR-140-5p誘導的炎癥模型細胞炎癥因子生成的作用。
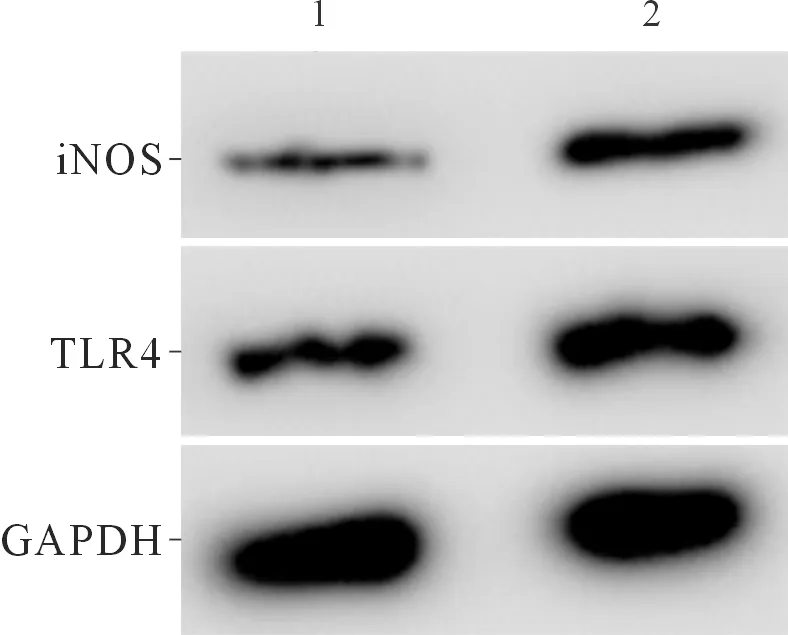
1:miR-140-5p組;2:miR-140-5p+TLR4
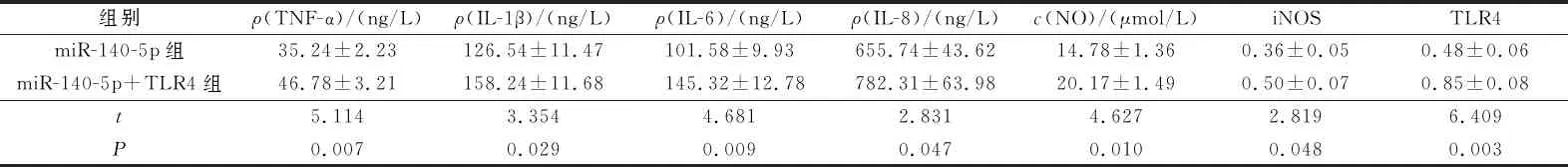
表4 2組細胞TNF-α、IL-1β、IL-6、IL-8分泌量,NO生成量及細胞中TLR4、iNOS蛋白表達水平的比較(n=3)
3 討論
RSV是一種常見的毛細支氣管炎病原體。研究[8]顯示,RSV刺激后支氣管上皮細胞分泌大量的炎癥介質,誘導炎癥發生。TNF-α、IL-1β、IL-6、IL-8是常見的促炎因子,其表達升高與支氣管哮喘關系密切[9]。NO是一種重要的信號傳遞分子,也是一種重要的炎癥介質,與免疫反應、神經傳導等有關[10]。研究[11-13]顯示,內源性NO是由NOS催化產生的,iNOS是NOS中與炎癥有關的亞型,其在受到炎癥因子、內毒素等刺激以后被激活,從而促使大量NO合成,而過量的NO能夠推動炎癥進展,誘導哮喘、癌癥、炎癥性腸道疾病等的發生。本研究結果顯示,RSV感染后的支氣管上皮細胞培養上清中TNF-α、IL-1β、IL-6、IL-8、NO含量均升高,細胞中iNOS蛋白表達水平也升高,提示RSV可誘導支氣管上皮細胞炎癥介質釋放,說明成功構建了支氣管上皮細胞炎癥模型。
miRNA是一類非編碼的RNA分子,能夠通過調控下游靶基因的表達發揮多重作用,在人體組織中普遍表達,參與調控不同類型細胞的生長、代謝、衰老、氧化應激等生理過程[14]。miR-140-5p是一個在人體內多種組織中表達的調節因子,參與腫瘤生長、關節炎進展等過程;與支氣管哮喘的發生發展關系密切[3-5]。本研究結果顯示,miR-140-5p在RSV誘導的炎癥模型支氣管上皮細胞中表達下調,而上調miR-140-5p表達可以減少炎癥模型細胞分泌TNF-α、IL-1β、IL-6、IL-8,降低細胞中iNOS蛋白的表達并減少NO的合成,說明miR-140-5p可抑制支氣管上皮細胞炎癥介質的釋放。
本研究還進一步探討了miR-140-5p的作用機制,發現miR-140-5p和TLR4的3’UTR有結合位點,并且上調miR-140-5p表達可以降低炎癥模型細胞中TLR4蛋白的表達。TLR是存在于細胞表面的免疫受體,可以識別某些病原體或某些特定的分子結構,引發炎癥介質釋放,參與免疫反應過程[15-16]。TLR4是第一個被發現的TLR家族成員,在幾乎所有的細胞系中均有表達,可以誘導IL-6、TNF-α等的表達,放大炎癥信號,激發炎癥瀑布鏈[17-18]。敲低TLR4能夠抑制RSV誘導的支氣管上皮細胞炎癥因子的釋放[19]。本研究結果表明,TLR4可以逆轉miR-140-5p誘導的炎癥模型支氣管上皮細胞炎癥介質的釋放抑制作用,提示上調miR-140-5p可通過降低TLR4表達減少支氣管上皮細胞炎癥介質的釋放。
總之,miR-140-5p在RSV誘導的支氣管炎癥中可能發揮抑制作用,作用機制與靶向下調TLR4的表達有關,這為miR-140-5p在支氣管炎癥中的應用提供了參考。今后我們會繼續探討miR-140-5p其他可能的靶向調控機制。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
光學精密工程(2016年6期)2016-11-07 09:07:19
海峽科技與產業(2016年3期)2016-05-17 04:32:12
