芥菜胞質雄性不育系結莢性狀的篩選和細胞學鑒定
2021-06-08 03:41:40劉志新徐玉穎姚培杰許可凈韓一鳴傅強萬正杰
華中農業大學學報(自然科學版) 2021年3期
劉志新,徐玉穎,姚培杰,許可凈,韓一鳴,傅強,萬正杰
華中農業大學園藝林學學院/園藝植物生物學教育部重點實驗室,武漢 430070
芥菜(Brassicajuncea)是一種原產于我國的十字花科蕓薹屬蔬菜[1]。研究表明,西北地區為我國芥菜的初生起源地,四川盆地為我國芥菜的次生起源地[2]。目前,芥菜在全國各地均有栽培,其中,長江中下游地區和西南地區是主產區[3-5]。由于芥菜具有營養物質豐富、栽培容易、多樣化的加工與食用方式等特點,深受消費者歡迎[6]。芥菜作為一種常異花授粉植物,新品種主要來自于系統育種和優勢雜交育種[7],芥菜雜交種的營養體在生長勢、抗逆性、豐產性等方面較常規種具有明顯優勢[8],雜交種生產過程中,人工授粉存在種子生產成本過高、生產效率低下的問題,因此,細胞質雄性不育系在芥菜雜交種選育和制種中得到了大規模的應用[9]。
長期以來,雄性不育作為植物雜種優勢利用和生殖發育研究的熱點內容之一,已取得大量研究成果[10]。余鳳群等[11]在對甘藍型油菜花藥敗育的研究中發現,花藥敗育主要分為小孢子發育異常、花粉母細胞發育異常、花粉囊發育異常3類。在微觀水平上,胡永敏等[12]進行了5種甘藍型油菜細胞質雄性不育系的細胞學觀察實驗,發現PolCMS材料的敗育大多發生在孢原細胞時期;許代香等[13]通過對甘藍型油菜顯性核不育系D3A的花藥敗育研究發現,不育系的花藥敗育主要是花粉囊的異常發育所導致。而ogu細胞質不育系材料則存在多種敗育方式,黃邦全等[14]通過種間雜交方式,將ogu蘿卜雄性不育胞質轉入紫菜薹中,經過多代回交,獲得了發育正常的紫菜薹ogu胞質不育系,其花藥敗育發生在孢原細胞分化期;田福發[15]通過種間雜交獲得了Pol紅菜薹不育系、ogu紅菜薹不育系材料,并且進行細胞學切片觀察,發現ogu紅菜薹不育系花藥在四分體時期,絨氈層細胞發育異常,擠壓四分體,最終導致絨氈層和四分體同時解體,引起花藥敗育。我們在實踐過程中發現,芥菜雜交育種過程中,以雄性不育系材料為母本創制雜交種,部分不育系材料會表現出不同程度的結莢畸形現象。為了揭示芥菜雄性不育系結莢畸形性狀的形成機制,筆者對59份芥菜材料進行了結莢情況的鑒定以及細胞學研究,以期為芥菜胞質雄性不育系的品種改良提供理論依據。
1 材料與方法
1.1 試驗材料
試驗材料為華中農業大學十字花科蔬菜遺傳育種課題組所收集來自全國各地的59份芥菜材料,經多年提純復壯,如表1所示,包括29份芥菜保持系、30份芥菜細胞質雄性不育系、hauCMS原始材料(FanY-9A)及其保持系(FanY-9B)。
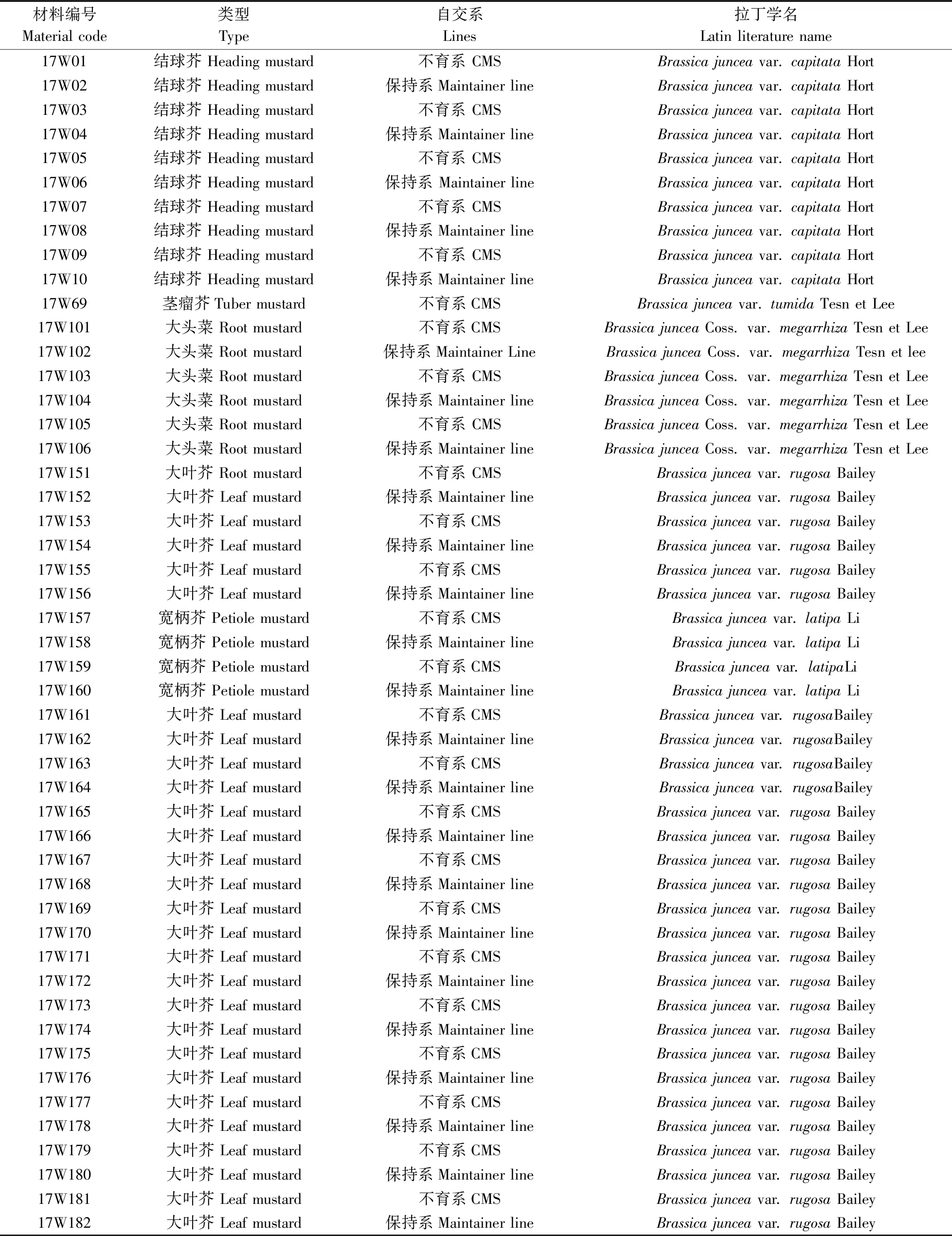
表1 供試材料Table 1 The list of experimental materials
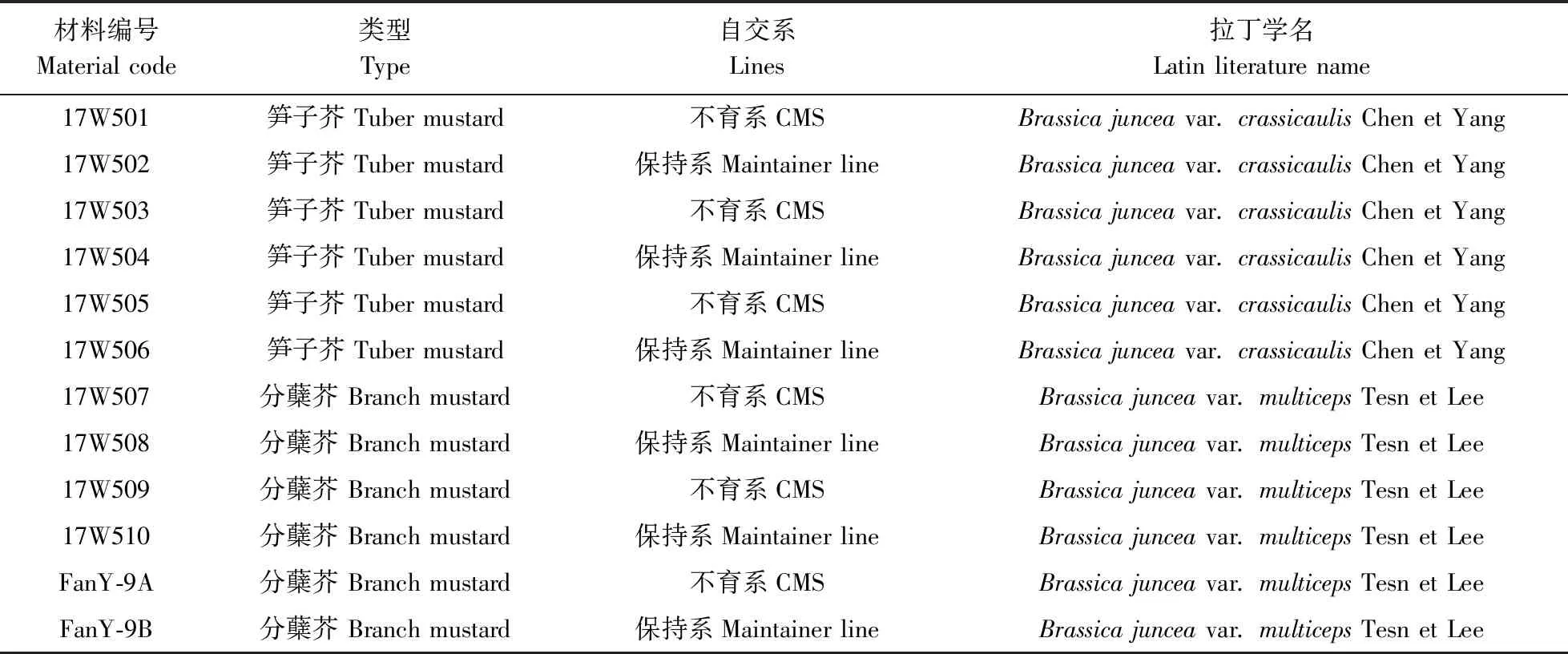
續表1 ContinuedTable 1
1.2 芥菜胞質雄性不育系材料不育類型鑒定
利用CTAB法提取30份芥菜細胞質雄性不育基因組總DNA,獲得基因orf288和orf138的保守序列[16],利用NCBI在線網頁設計引物,orf138正向引物:GAAACGGGAAGTGACAATAC,反向引物:GCATTATTTTCTCGGTCCAT;orf288正向引物CAATGGCTATTACCCCTGCT;反向引物: ATGCCGGGCATTTACCTGATGG。
1.3 芥菜材料的結莢情況調查
將59份材料于2017年9月同期播種育苗,之后統一水肥管理,雜交授粉完成后,待種莢成熟時,取典型角果,拍照記錄。
1.4 花蕾的半薄切片技術
材料盛花期,選取生長良好的花序參照文獻[17]制樣,于顯微鏡下觀察。
2 結果與分析
2.1 芥菜不育系材料不育胞質類型鑒定
利用CTAB法提取30份芥菜細胞質雄性不育系材料,利用設計的2對引物進行PCR擴增、瓊脂糖凝膠電泳檢測,結果(圖1)顯示:以17W05、17W155、17W179為代表的23份材料屬于hau不育胞質類型,以17W501和17W173為代表的7份材料屬于ogu不育胞質類型。
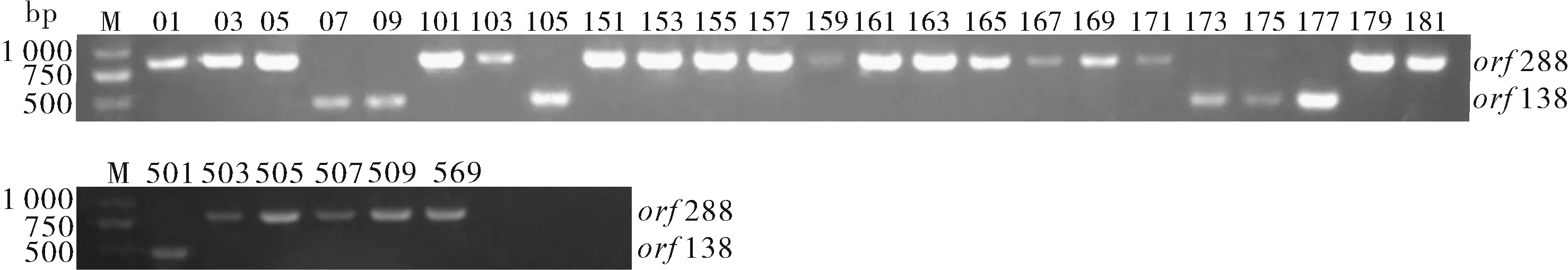
圖1 芥菜雄性不育系材料不育類型鑒定Fig.1 Identification of CMS types of Brassica juncea
2.2 芥菜細胞質雄性不育系的結莢形態
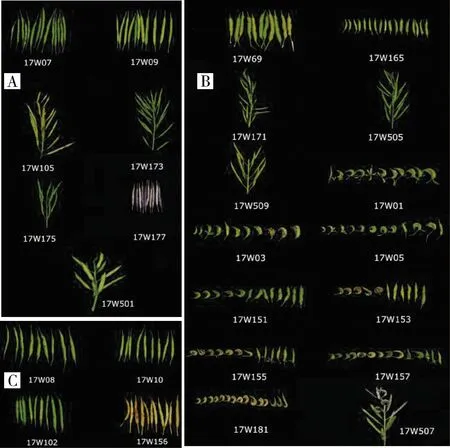
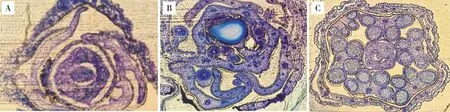
對59份芥菜材料結莢形態的調查結果(圖2)顯示,7份芥菜ogu雄性不育系沒有畸形情況發生,莢果飽滿直立,形態具有一致性(圖2A);芥菜hau雄性不育系材料在畸形方面表現較大的差異(圖2B),其中,17W165、17W171、17W505、17W509和17W69表現出良好的結莢特性,果莢直立飽滿,其他材料則表現出不同程度的畸形;所有的芥菜保持系材料莢果均呈現飽滿、直立的狀態(圖2C)。
2.3 芥菜細胞質不育系花藥敗育及花器官發育的細胞學研究
1)芥菜oguCMS材料花藥敗育及花器官發育觀察。對芥菜ogu細胞質雄性不育系17W173及其對應保持系材料17W174進行半薄切片的細胞學觀察(圖3),可以發現,從花器官的整體發育來看,不育系絨氈層出現降解,小孢子集結成團不能分散,保持系則較為正常。進一步觀察其花藥的敗育情況(圖4),可以發現,四分體時期,絨氈層細胞發育異常,細胞出現空泡化現象;單核靠邊期,絨氈層發生降解,藥室中央的小孢子聚集成團;雙核期,絨氈層已經基本降解;三核期時,小孢子出現空泡化跡象,花藥敗育。
A.ogu細胞質雄性不育系;B. hau細胞質雄性不育系;C.保持系。 A.ogu CMS;B.hau CMS; C.Maintainer lines.

A.ogu細胞質雄性不育系; B.保持系。A.ogu CMS; B.Maintainer line.
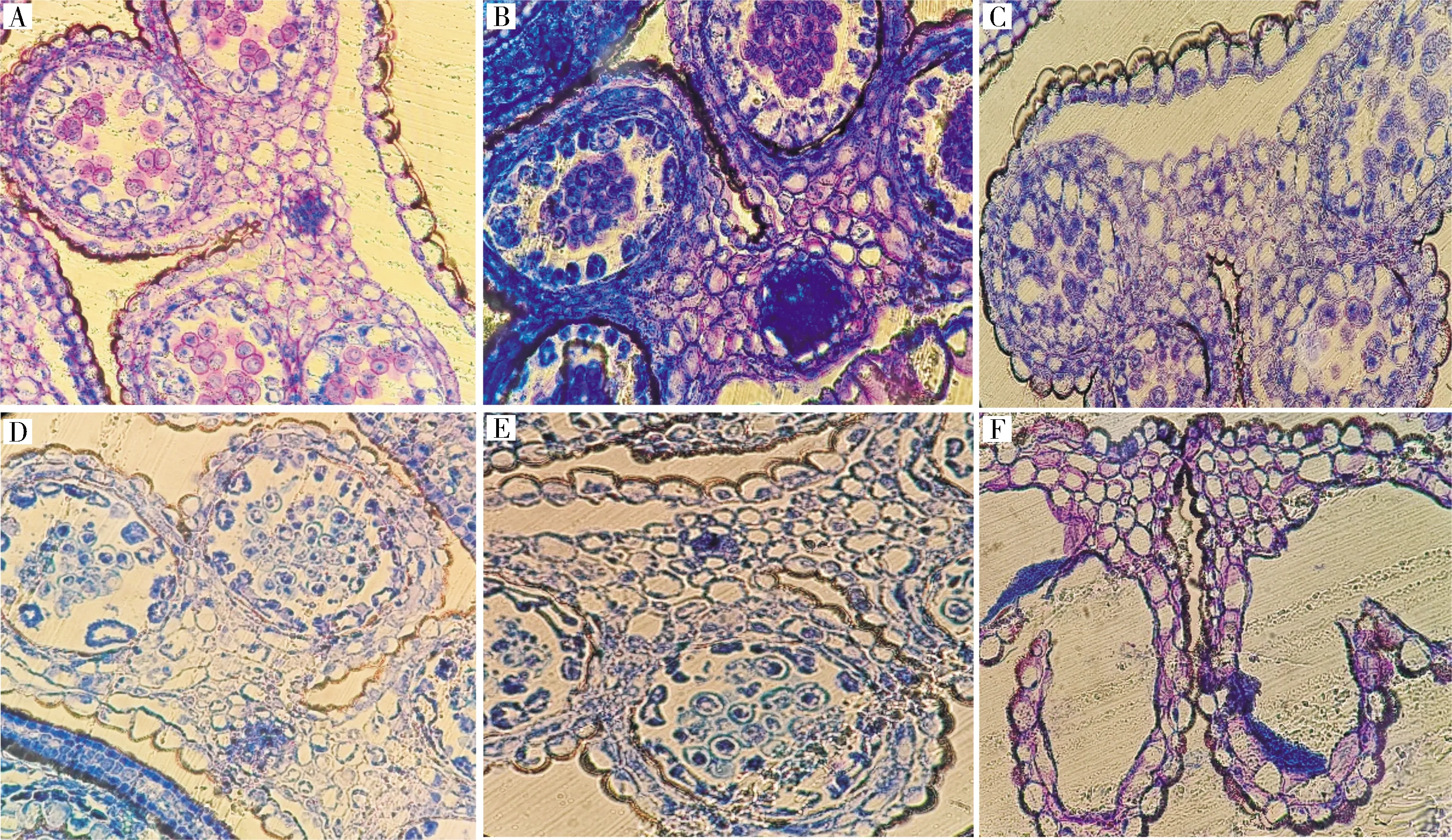
A.四分體時期; B.單核期; C.單核靠邊期; D.雙核期; E.三核期; F.釋放花粉。A.Tetrad; B.Mononuclear; C.Mononuclear marginal; D.Dikaryotic; E.Trinuclear; F.Pollen release.
2)芥菜hau細胞質雄性不育系材料花器官發育及花藥敗育觀察。將果莢正常和果莢畸形的芥菜hau細胞質雄性不育系和保持系的花器官進行對比(圖5),可以發現,2類材料的雄蕊敗育基本都由絨氈層的異常發育所引起;而結莢畸形材料,則直接形成了雌蕊-雄蕊嵌合體,嵌合體內生長著多于正常數量的胚珠。對結莢正常材料雄蕊進行敗育觀察(圖6),可以看到,孢原細胞時期,產生心皮組織;花粉母細胞時期,絨氈層細胞異常膨大;在四分體時期,絨氈層發育異常,小孢子因養分不足隨之降解,花藥敗育。對結莢畸形材料的敗育過程進一步觀察(圖7),可以看到,雌蕊-雄蕊嵌合體在花藥發育初期就開始形成,此時產生心皮組織而非正常的雄蕊原基,之后,心皮組織不斷生長發育,形態各異,整個過程幾乎觀察不到正常的花器官。
A.正常莢果不育系; B.畸形莢果不育系; C.保持系。A.Normal pods with hau CMS; B.Abnormal pods with hau CMS; C.Maintainer line.
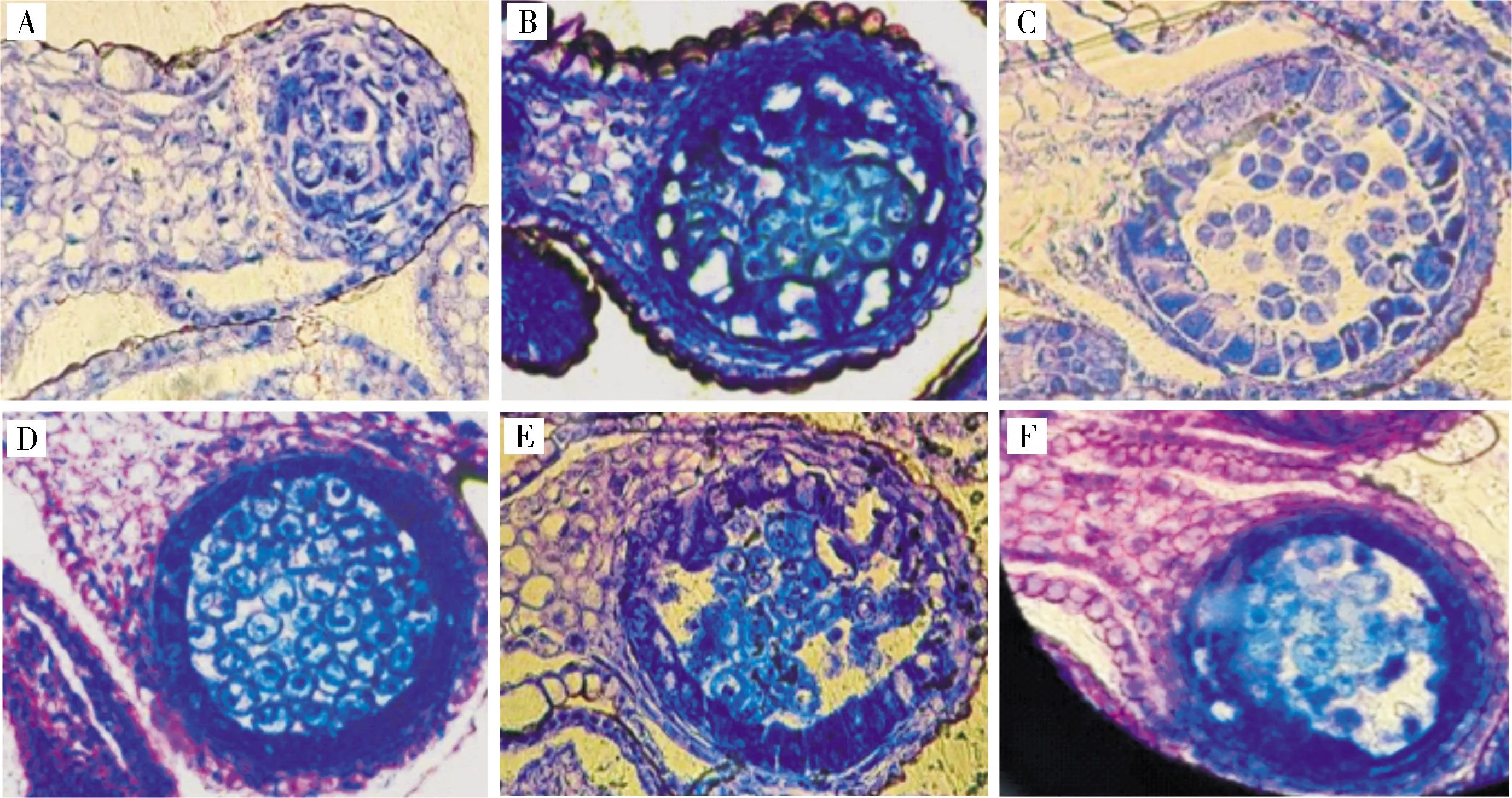
A.孢原細胞時期; B.花粉母細胞時期; C.四分體時期; D-F.配子體發育時期。A.Sporogenous cell stage; B.Pollen mother cell stage; C.Tetrad stage; D-F.Gametophyte development stage.
3 討 論
在植物新品種的選育過程中,為避免長期使用單一不育胞質所帶來的潛在危害,需要引進不同類型的胞質并且對其快速鑒定,而分子標記則具有準確、省時、高效的特點[18];本研究利用2對特異性引物(orf138和orf288)對芥菜CMS材料的不育胞質類型進行快速鑒定,發現orf138標記可鑒定芥菜oguCMS,而orf288標記則可將芥菜hauCMS鑒定出來,相較于觀察花器官的方式簡單且快速。從59份材料結莢性狀的田間統計結果中可以發現,所有的保持系和oguCMS結莢表現均非常優異,無畸形情況發生,這與鄒瑞昌[19]的研究結果相似。芥菜hauCMS是華中農業大學1999年所發現的天然芥菜型油菜雄性不育系,具有不育度和不育株率100%的特點,長期以來,在十字花科植物雜種優勢利用中得到了大量的應用[3]。施展等[20]對芥菜hauCMS 進行了綜合遺傳分類后,將芥菜hauCMS的不育細胞質轉育到大白菜中,創制了大白菜hauCMS種質資源6w-9605A。張艷等[21]以甘藍型油菜hauCMS為母本,菜心為父本,進行種間雜交和連續回交,創制了新型菜心hauCMS不育材料。隨著對芥菜hauCMS不育資源利用和研究的不斷深入,筆者課題組也發現其不足之處,將芥菜hauCMS不育源轉入芥菜后,部分材料存在果莢畸形的情況,實際種子生產中,果莢畸形會嚴重影響種子品質甚至制種產量,部分芥菜hauCMS材料結莢畸形可能會影響其后續在十字花科作物中的推廣利用,因此,對芥菜hauCMS不育胞質進行遺傳改良顯得尤為重要。
本研究進一步對芥菜hauCMS和oguCMS材料進行花藥敗育的細胞學觀察,可以發現,芥菜oguCMS和果莢正常的hauCMS材料,雄蕊的敗育均由絨氈層的異常發育所引起,其雌蕊進行正常的授粉受精后,果莢仍能正常發育,這與單奇偉等[22]的研究結果類似;果莢畸形的芥菜hauCMS材料在其花器官發育過程中,除絨氈層發育異常外,還伴隨著雌蕊-雄蕊嵌合體的發生,最終導致果莢畸形,可見,雌蕊-雄蕊嵌合體的形成是造成果莢畸形的原因,這與文獻[23]的研究結果不一致,極有可能是核質互作效應引起的[24]。本研究通過細胞學水平的研究證實結莢畸形由雌蕊-雄蕊嵌合體的生成引起,但更深層次的機制尚待探究,根據花發育模型假說,B基因組是調控雄蕊發育、引起雄蕊心皮化形成嵌合體的關鍵[25],因此,后續可以從B基因組著手,對芥菜hauCMS進行深入研究,探究其敗育的分子機制。
