墨蘭×兔耳蘭雜交后代2n雄配子發(fā)生及其細(xì)胞學(xué)機(jī)制
2021-06-08 04:55:54郭和蓉朱嬌2杜國(guó)輝黃紫婷謝利魏倩張志勝
關(guān)鍵詞:途徑
郭和蓉,朱嬌2,杜國(guó)輝,黃紫婷,謝利,魏倩,張志勝
1.華南農(nóng)業(yè)大學(xué)林學(xué)與風(fēng)景園林學(xué)院/廣東省植物分子育種重點(diǎn)實(shí)驗(yàn)室,廣州 510642;2.山東省農(nóng)業(yè)科學(xué)院休閑農(nóng)業(yè)研究所,濟(jì)南 250100
未減數(shù)配子是由于大、小孢子形成和發(fā)育過程中出現(xiàn)異常而形成具有孢子體染色體數(shù)目(2n)的配子,也被稱為2n配子[1]。自然界植物2n配子發(fā)生十分普遍[1-3],2n配子參與下的有性多倍化是植物多倍體形成的主要機(jī)制[3-4],2n配子途徑也是人工創(chuàng)造多倍體資源和選育多倍體品種的有效途徑[4-6]。因此,研究2n配子發(fā)生及其細(xì)胞學(xué)機(jī)制,對(duì)深入研究植物多倍體進(jìn)化機(jī)制,創(chuàng)建高效發(fā)生2n配子資源,推動(dòng)多倍體育種均具有重要意義。
蘭花2n配子發(fā)生已有一些研究報(bào)道,蘭屬蘭花(Cymbidium)[6]、蝴蝶蘭(Phalaenopsis)[7-8]、石斛蘭(Dendrobium)[9-10]等均可產(chǎn)生2n配子,雜種2n配子發(fā)生率比非雜種的高[6]。部分學(xué)者對(duì)百合(Liliumauratum)[13]、香石竹(Dianthuscaryophyllus)[14]、燕麥(Avenaventricosa)[15]和蝴蝶蘭[16]等2n配子形成的細(xì)胞學(xué)機(jī)制[11-12]進(jìn)行了探討,但在蘭屬蘭花上缺乏系統(tǒng)的研究報(bào)道。
多倍體育種是蘭花育種的重要方法,目前商業(yè)化的蝴蝶蘭、石斛蘭、大花蕙蘭(Cymbidiumhybridum)品種多數(shù)為多倍體[17-19],但墨蘭(C.sinense)等國(guó)蘭商業(yè)化的品種均是二倍體。本研究以墨蘭、兔耳蘭(C.lancifolium)及其雜種后代為材料,探討蘭屬蘭花2n雄配子發(fā)生及其細(xì)胞學(xué)機(jī)制,創(chuàng)建高頻發(fā)生2n雄配子資源,旨在為國(guó)蘭多倍體育種和蘭花多倍體進(jìn)化研究積累材料。
1 材料與方法
1.1 試驗(yàn)材料
試驗(yàn)材料為‘小香墨蘭’(C.sinense‘Xiao xiang’)、兔耳蘭及其31份雜交后代,雜交后代中的30份材料用于2n雄配子發(fā)生途徑觀察,1份材料用于小孢子母細(xì)胞減數(shù)分裂過程觀察。上述材料種植于華南農(nóng)業(yè)大學(xué)增城基地蘭花大棚中,常規(guī)栽培管理。植株開花后,取不同發(fā)育時(shí)期的花朵或花粉塊固定。
1.2 細(xì)胞學(xué)觀察
取‘小香墨蘭’、兔耳蘭及其30份雜交后代成熟花粉期的花粉塊和編號(hào)45-1 后代不同發(fā)育時(shí)期花朵或花粉塊于卡諾固定液(V95%乙醇∶V冰乙酸=3∶1)中固定24 h,然后轉(zhuǎn)移至70%乙醇于4 ℃冰箱保存?zhèn)溆谩?/p>
參考周建金等[20]的方法制片,制片后在Olympus-I×71倒置顯微鏡和ZEISS光學(xué)顯微鏡下觀察,每片隨機(jī)選擇10個(gè)視野拍照。每份材料觀察3個(gè)花粉塊,觀察小孢子母細(xì)胞減數(shù)分裂時(shí),在40×或100×下拍照,以100個(gè)小孢子母細(xì)胞為1次重復(fù),重復(fù)3次。
1.3 統(tǒng)計(jì)分析
二分體發(fā)生率=(二分體個(gè)數(shù)/觀察總孢子數(shù))×100%;三分體發(fā)生率=(三分體個(gè)數(shù)/觀察總孢子數(shù))×100%;二分體途徑對(duì)未減數(shù)雄配子發(fā)生的貢獻(xiàn)率=(二分體途徑產(chǎn)生的未減數(shù)雄配子數(shù)/未減數(shù)雄配子總數(shù))×100%;三分體途徑對(duì)未減數(shù)雄配子發(fā)生的貢獻(xiàn)率=(三分體途徑產(chǎn)生的未減數(shù)雄配子數(shù)/未減數(shù)雄配子總數(shù))×100%;未減數(shù)雄配子發(fā)生率=(N2D+NTr)/T×100%,其中D代表二分體,Tr代表三分體,T代表總花粉數(shù)。
數(shù)據(jù)采用SPSS 18.0軟件進(jìn)行統(tǒng)計(jì)分析,以鄧肯氏新復(fù)極差法(Duncan’s)測(cè)驗(yàn)在α=0.05水平上的差異顯著性。
2 結(jié)果與分析
2.1 ‘小香墨蘭’×兔耳蘭雜交后代2n雄配子發(fā)生
試驗(yàn)結(jié)果(圖1)顯示,30份‘小香墨蘭’×兔耳蘭雜交后代2n雄配子發(fā)生率3 a平均為2.28%,顯著高于‘小香墨蘭’(0.19%)和兔耳蘭(0.22%),表明種間雜交顯著提高了雜種后代2n雄配子發(fā)生率。不同 ‘小香墨蘭’×兔耳蘭雜交后代的2n雄配子發(fā)生率不同(表1),平均值變幅為0.87%~ 6.07%。后代45-60 的2n雄配子發(fā)生率最高,為6.07%;45-29的發(fā)生率最低,為0.87%。年份對(duì)大多數(shù)雜種后代2n雄配子發(fā)生率影響顯著,但對(duì) 45-113、45-116、45-105和45-29等雜交后代影響不顯著。
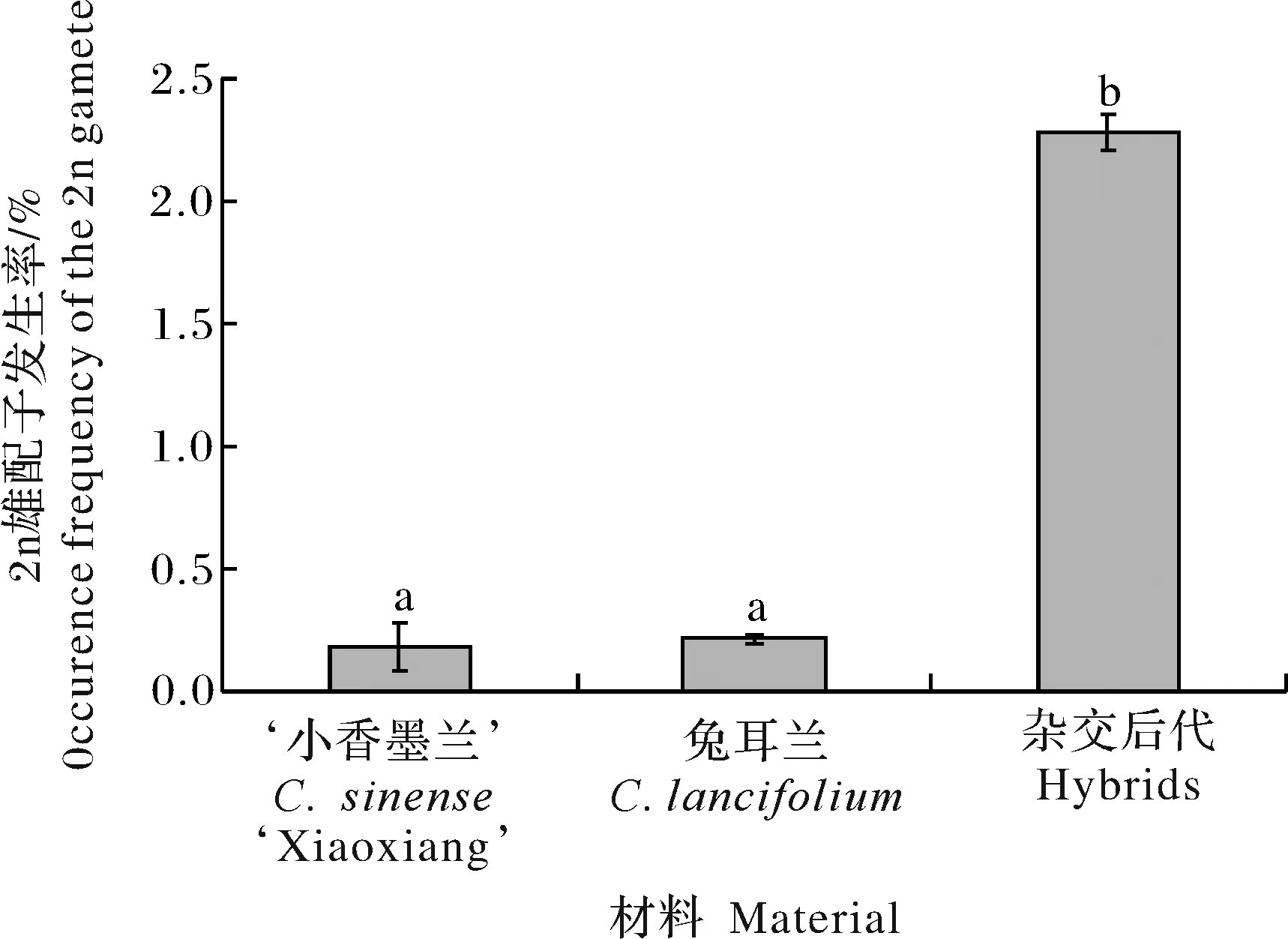
不同小寫字母表示差異顯著,P<0.05。Means in the same column with different small letters indicated significant difference at α=0.05 level.
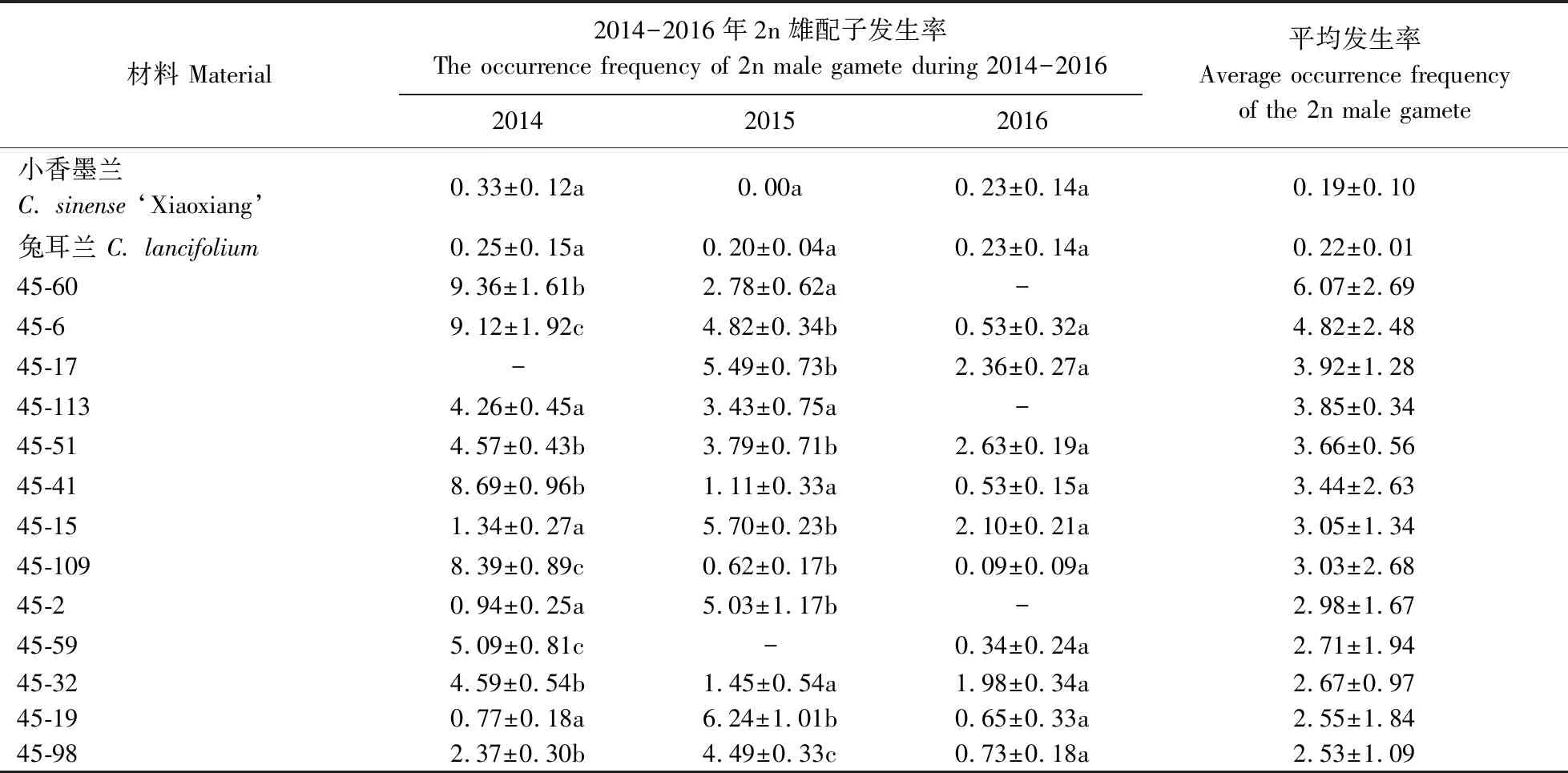
表1 不同年份‘小香墨蘭’×兔耳蘭雜交后代2n雄配子發(fā)生率Table 1 The occurrence frequency of unreduced male gamete in hybrids of C. sinense ‘Xiaoxiang’ × C. lancifolium %
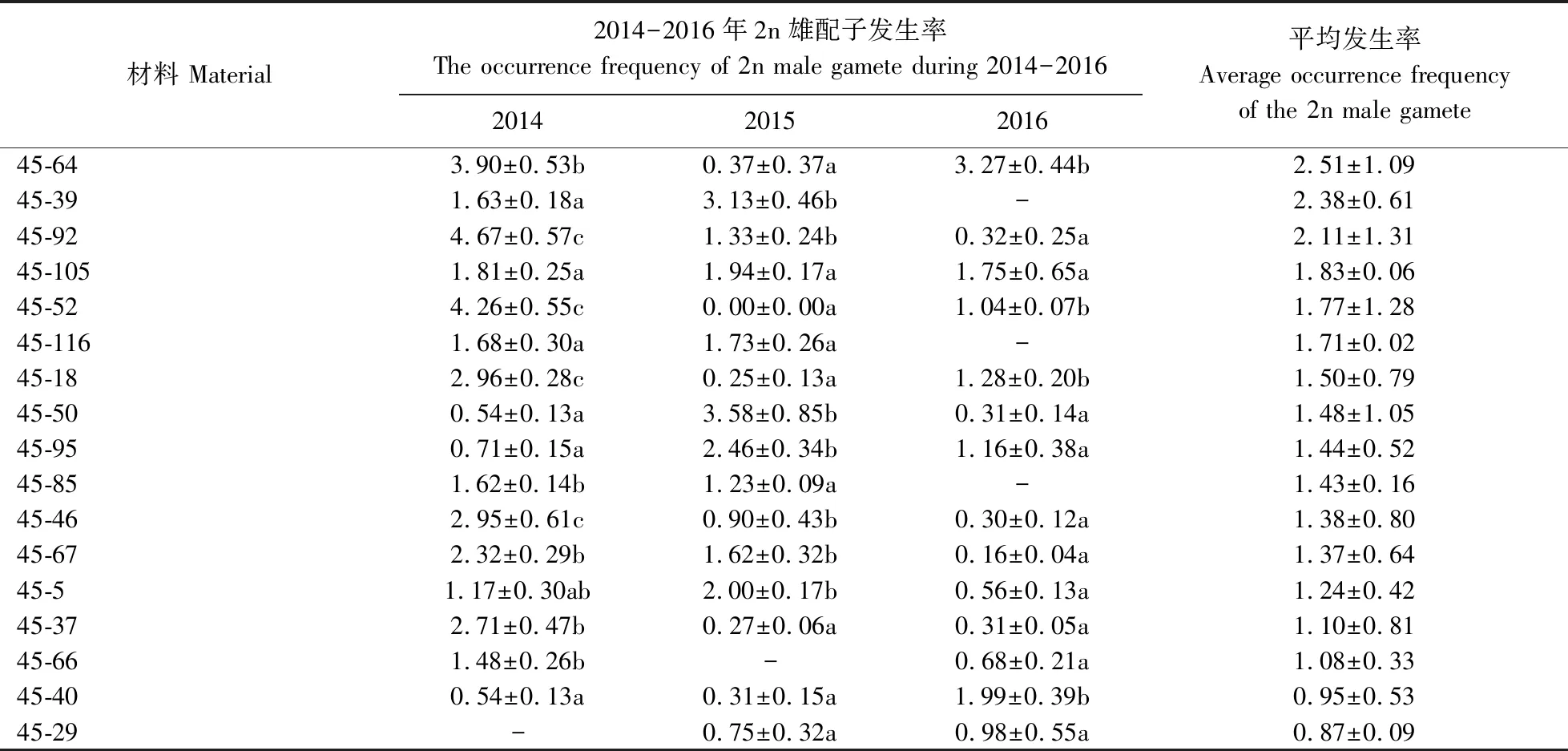
續(xù)表1 ContinuedTable 1
2.2 ‘小香墨蘭’×兔耳蘭雜交后代2n雄配子發(fā)生途徑
對(duì)‘小香墨蘭’×兔耳蘭雜交后代2n雄配子發(fā)生途徑進(jìn)行了鑒定。結(jié)果(圖2)顯示,在‘小香墨蘭’×兔耳蘭雜交后代的花粉塊中,除了正常的四分體外,還有一定比例的二分體(圖2A)和三分體(圖2B),二分體中有2個(gè)2n雄配子,三分體包含了1個(gè)2n雄配子和2個(gè)n配子。說(shuō)明‘小香墨蘭’×兔耳蘭雜交后代2n雄配子發(fā)生主要為二分體和三分體途徑。

A:45-51成熟花粉,箭頭示二分體;B:45-98成熟花粉,箭頭示三分體。A:Maturepollen of 45-51,dyads(arrow);B:Mature pollen of 45-98,triads(arrow).
結(jié)果顯示,所有雜交后代2n雄配子發(fā)生均通過二分體和三分體途徑,但不同后代2n雄配子發(fā)生的主要途徑不完全相同(圖3)。二分體途徑對(duì) 45-60、45-51、45-41、45-109和45-2等后代2n雄配子形成的貢獻(xiàn)率分別為63.98%、64.41%、78.91%、72.23%和79.24%,因此,這些后代2n配子形成主要通過二分體途徑。三分體途徑對(duì)45-39、45-18、45-85、45-66和45-5等后代2n雄配子形成的貢獻(xiàn)率分別為70.37%、77.39%、85.20%、85.33%和88.04%,其2n雄配子形成主要通過三分體途徑。
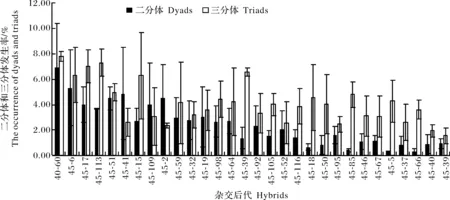
圖3 ‘小香墨蘭’×兔耳蘭雜交后代二分體和三分體的平均發(fā)生率Fig.3 Average incidence of dyads and triads in hybrids of C. sinense ‘Xiaoxiang’ ×C. lancifolium
2.3 ‘小香墨蘭’×兔耳蘭雜交后代2n雄配子發(fā)生的細(xì)胞學(xué)機(jī)制
‘小香墨蘭’×兔耳蘭雜交后代45-1的2n雄配子發(fā)生率為4.94%。對(duì)其小孢子母細(xì)胞減數(shù)分裂過程進(jìn)行觀察(圖4),結(jié)果顯示:45-1小孢子母細(xì)胞減數(shù)分裂存在不同步現(xiàn)象,在中期Ⅰ,5.79%小孢子母細(xì)胞仍處在終變期(圖4A),這些小孢子母細(xì)胞不進(jìn)行減數(shù)分裂第一次分裂(圖4B),而直接進(jìn)行減數(shù)分裂第二次分裂(圖4C),形成二分體,最終發(fā)育成2個(gè)2n雄配子(圖4F)。

A:中期Ⅰ,示處于中期Ⅰ(箭頭)和終變期小孢子母細(xì)胞(箭頭),Bar=10 μm;B:后期Ⅰ,示未進(jìn)行第一次分裂的小孢子母細(xì)胞,Bar=20 μm;C:中期Ⅱ,示未分裂的小孢子母細(xì)胞染色體排列在赤道板上(箭頭),Bar=20 μm;D:中期Ⅱ,示融合紡錘體(長(zhǎng)箭)、垂直紡錘體(短箭)和平行紡錘體(箭頭),Bar=20 μm;E:后期Ⅱ,示融合紡錘體引起兩組染色體被牽引至同一子細(xì)胞中(箭頭),Bar=10 μm;F:成熟花粉期,示二分體(箭頭)和四分體(箭頭),Bar=10 μm;G:中期Ⅱ,示三極紡錘體(箭頭)和平行紡錘體,Bar=10 μm;H:后期Ⅱ,示2組染色體被牽引至同一子細(xì)胞中(箭頭),Bar=20 μm;I:成熟花粉期,示三分體,包含1個(gè)2n花粉(箭頭)和2個(gè)n配子(箭頭),Bar=10 μm。 A:Meiosis metaphaseⅠ,indicating microsporocyte at metaphaseⅠ(arrow) and diakinesis (arrow ),Bar=10 μm;B:AnaphaseⅠ,indicating microsporocyte failed to carry out meiosisⅠ(arrow), Bar=20 μm;C:MetaphaseⅡ,indicating chromosomes arranged on the equatorial plate of the microsporocyte failed to carry out meiosisⅠ(arrow),Bar=20 μm;D:MetaphaseⅡ,indicating fused (long arrow),parallel(short arrow)and perpendicular(arrow )spindles,Bar=20 μm;E:AnaphaseⅡ,indicating two set of chromosomes being pulled to one daughter cell by fused spindle,Bar=10 μm;F:Mature pollen stage,indicating dyad(arrow) and tetrad(arrow),Bar=10 μm;G: MetaphaseⅡ,indicating parallel and tripolar spindle,Bar=10 μm;H:AnaphaseⅡ,indicating two sets of chromosomes being pulled to one daughter cell(arrow),Bar=20 μm;I:Mature pollen stage,indicating triad with one 2n gamete(arrow) and two n gametes(arrow ),Bar=10 μm.
在減數(shù)分裂中期Ⅱ,1.63%二分體形成融合紡錘體(圖4D),導(dǎo)致后期Ⅱ(圖4E)兩套染色體進(jìn)入同一個(gè)子細(xì)胞形成2個(gè)2n雄配子(圖4F);4.63%二分體形成三極紡錘體(圖4G),導(dǎo)致在減數(shù)分裂第二次分裂后期,2組染色體進(jìn)入同一子細(xì)胞中,另2組染色體分別進(jìn)入不同子細(xì)胞(圖4H),形成三分體(圖4I),產(chǎn)生1個(gè)2n雄配子和2個(gè)n雄配子。
3 討 論
高效發(fā)生2n配子資源的創(chuàng)建有助于利用2n配子途徑選育多倍體植物新品種。已有研究表明,不同種植物、同種植物不同個(gè)體2n配子發(fā)生率差異明顯[2-3],遠(yuǎn)緣雜交是獲得高效發(fā)生2n配子資源的有效手段[6,21]。本研究中墨蘭和兔耳蘭種間雜交后代2n雄配子發(fā)生率顯著高于親本,45-60的 2n雄配子發(fā)生率是其父本的30倍、母本的32倍,45-51、45-15等后代2n雄配子平均發(fā)生率大于3%,每年發(fā)生率均在1%以上,這些資源的獲得為國(guó)蘭多倍體育種提供了親本材料。
蘭花小孢子形成過程中,減數(shù)分裂前加倍,減數(shù)分裂Ⅰ、Ⅱ后期失調(diào),不進(jìn)行減數(shù)分裂Ⅰ或Ⅱ以及減數(shù)分裂Ⅱ紡錘體的異常是2n雄配子和多倍體配子發(fā)生的主要原因[22]。本研究發(fā)現(xiàn),墨蘭和兔耳蘭種間雜種2n雄配子均通過二分體和三分體方式產(chǎn)生,二分體是由于小孢子母細(xì)胞減數(shù)分裂不同步,導(dǎo)致減數(shù)分裂Ⅰ不分裂,而減數(shù)分裂Ⅱ正常分裂或由于減數(shù)分裂Ⅱ細(xì)胞紡錘體異常定向,形成融合紡錘體所致;三分體的形成都是由于減數(shù)分裂Ⅱ紡錘體異常定向形成三極紡錘體所致。這一結(jié)果與前人在其他品種蘭花上的研究結(jié)果[16,22]不完全一致,朱嬌等[16]對(duì)蝴蝶蘭未減數(shù)配子發(fā)生機(jī)制的研究結(jié)果表明,二分體的形成是由于小孢子母細(xì)胞減數(shù)分裂不同步導(dǎo)致的,沒有發(fā)現(xiàn)因減數(shù)分裂Ⅱ紡錘體異常定向?qū)е露煮w發(fā)生的現(xiàn)象,這可能是材料不同所致。進(jìn)一步的研究表明,不同后代2n雄配子發(fā)生途徑不完全相同,一些后代主要通過二分體方式產(chǎn)生,另一些后代則主要通過三分體途徑發(fā)生,說(shuō)明2n雄配子發(fā)生受多基因控制,鑒定、克隆和解析這些基因的功能將成為今后研究的主要方向之一。
2n配子發(fā)生主要受遺傳因素控制[12,23],但外界環(huán)境也會(huì)在很大程度上影響其發(fā)生頻率[14,24-26],如高溫[27-28]和低溫[29-30]是具有重要影響的環(huán)境因素。Zhou等[14]研究表明,香石竹同一品種在3月、6月、9月和12月的2n雄配子發(fā)生率差異明顯,月份對(duì)不同品種2n雄配子發(fā)生影響也不同。本研究對(duì)30份雜交后代連續(xù)3 a的觀測(cè)結(jié)果也表明,墨蘭和兔耳蘭多數(shù)雜交后代不同年份間的2n配子發(fā)生率差異顯著,且2n配子發(fā)生率高的后代年份間差異大,其原因有待進(jìn)一步研究。
猜你喜歡
語(yǔ)數(shù)外學(xué)習(xí)·高中版中旬(2023年2期)2023-05-10 13:26:53
語(yǔ)數(shù)外學(xué)習(xí)·高中版中旬(2022年5期)2022-07-13 20:47:51
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年10期)2019-11-25 07:33:58
中學(xué)生數(shù)理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
中學(xué)生百科·大語(yǔ)文(2017年10期)2017-11-04 06:56:38
中國(guó)衛(wèi)生(2016年3期)2016-11-12 13:23:26
公民與法治(2016年22期)2016-05-17 04:20:13
中國(guó)衛(wèi)生(2014年12期)2014-11-12 13:12:52
癌變·畸變·突變(2014年6期)2014-02-27 06:15:03
