三種餌料對盾形陀螺珊瑚生長性能的影響
2021-06-12 00:23:42鄭子豪黃文靖張艷萍
廣東海洋大學學報 2021年3期
鄭子豪,沈 城,黃文靖,張艷萍,李 鋒,劉 麗
(1.廣東海洋大學水產學院,廣東 湛江 524088;2.徐聞國家級珊瑚礁保護區管理局,廣東 湛江 524100)
珊瑚礁是全球最重要的生態系統之一,提供極其重要的生態服務功能和巨大的經濟利益[1]。近年來,由于人為因素影響,全球范圍內珊瑚一直處于退化狀態[2-4],珊瑚小塊移植、培育珊瑚幼體、人工珊瑚礁投放等方式是修復珊瑚礁的主要手段[5]。珊瑚小塊移植作為珊瑚保護的主要手段,其優點是可以通過無性繁殖方式在短時間內得到大量可用于移植的珊瑚碎片,可以大大降低珊瑚礁修復成本。因為珊瑚小塊移植存活率與珊瑚的大小成正比[6-7],所以最大化加速珊瑚小塊成為提升珊瑚移植效率的關鍵因素。珊瑚具有自養和異養兩種營養方式,有研究指出,異養營養最高可以為珊瑚提供鈣化所需有機碳的66%[8]。異養營養還可以為珊瑚提供氮和磷,這兩種營養物質無法從蟲黃藻的光合作用中獲取,必須通過異養營養獲得[9]。所以,確定珊瑚的合適餌料是解決珊瑚人工養殖難題的重要途徑。
輪蟲、鹵蟲是目前進行珊瑚養殖的主要活體餌料[10-11],但由于珊瑚觸手大小、捕食能力及捕食方式的區別,不同珊瑚的適口餌料有所區別。珊瑚的捕食對象不僅僅是浮游動物,近來許多研究表明珊瑚也會將浮游植物作為捕食對象[12-13],小球藻作為富含不飽和脂肪酸的海洋微藻,如果以小球藻作為珊瑚餌料,可以為珊瑚生長提供充足的不飽和脂肪酸。Colan 等[14-15]對比了鹵蟲、微藻以及配合餌料對鹿角珊瑚生長性能的影響,結果表明鹵蟲、微藻喂食下珊瑚的生長性能均高于配合餌料投喂的珊瑚,這說明活體餌料是珊瑚養殖的最佳選擇。上述實驗中使用未過濾海水養殖的珊瑚的生長性能也不及兩種活體餌料,說明珊瑚對于不同種類活體餌料有選擇性,挑選不同種最適合的活體餌料能使珊瑚獲得更高的生長性能。通過人工喂食方式可以大大降低珊瑚養殖和移植成本,這無疑能有力地促進珊瑚礁的恢復工作。
本實驗選取生態位較寬的盾形陀螺珊瑚(Turbinaria peltata)作為實驗材料,隨著徐聞地區水文條件的變化,該物種近年來逐漸成為當地的優勢種[4],說明該物種適合在徐聞地區移植。塊狀珊瑚均存在生長速度較慢的現象,探索如何最大化增加其生長速度是進行該種珊瑚養殖的關鍵。本實驗選取三種不同粒徑大小的浮游生物(輪蟲、鹵蟲、小球藻)作為盾形陀螺珊瑚的餌料,對比喂食不同餌料后珊瑚的生長情況及蛋白、脂肪和脂肪酸變化,為探索適合盾形陀螺珊瑚的合適活體餌料提供基礎。
1 材料與方法
1.1 珊瑚暫養與分割
研究所需珊瑚樣品于2020 年5 月采自徐聞珊瑚礁國家級自然保護區(109°55′18.28″E,20°16′30.55″N),采樣點深度4 m 左右。將整塊盾形陀螺珊瑚運回實驗室暫養在水溫25 ℃、PH8.1、鹽度30 的50 L 水族箱中,光源由全光譜LED 珊瑚燈(Maxspect)提供,每日7:00 至19:00 提供光照,缸底有效輻射約為90 μmol/(m2·s)。暫養到7 d 珊瑚觸手正常伸展,于21:00 將其切割成具有14 個螅體的珊瑚小塊,共20 個,將小塊用阿隆發膠水固定在塑料底座上。將珊瑚小塊5 個一組隨機分成4 組置于5 L 培養箱中,培養至珊瑚觸手正常伸展開始投喂實驗,珊瑚恢復期間不進行喂食。
1.2 分組與投喂方式
實驗設投喂鹵蟲、輪蟲、小球藻和不投喂對照組,每組5 個平行。每天9:00 投喂餌料0.5 g,投喂前1 h 將珊瑚小塊轉移到2 L 燒杯中,通過打氧泵向燒杯持續充氧保證餌料懸浮,投喂1 h 后將珊瑚小塊表面沖洗干凈放回原養殖箱中。本實驗所選三種餌料均取自廣東海洋大學,三種餌料的脂肪酸組成結果見表1。實驗共45 d。在開始實驗的第一天和最后一天稱量珊瑚骨骼干質量,實驗結束后所有樣品立刻用于蛋白、脂肪及脂肪酸提取。
1.3 盾形陀螺珊瑚對三種餌料的攝食情況
為監測盾形陀螺珊瑚對三種浮游生物的攝食速率,在測該參數前需將珊瑚樣品饑餓24 h,將珊瑚樣品放置于1.25 L 的亞克力培養室中,培養室中的培養條件與原養殖箱中的條件相同,同時設置一個空白培養室作為對照。待珊瑚觸手伸展后,向培養室中加入兩種同濃度的餌料濃縮液,1.5 h 后計數生物活餌料殘余量和差值。
1.4 測定指標
1.4.1珊瑚骨骼質量測定 用天平測量珊瑚樣品的浮力質量與空氣中的質量,通過放入燒杯中的排水量與海水密度的比值計算珊瑚體積,同時根據稱量時海水溫度與鹽度計算海水的密度,根據阿基米德原理,計算出樣品的骨骼質量。
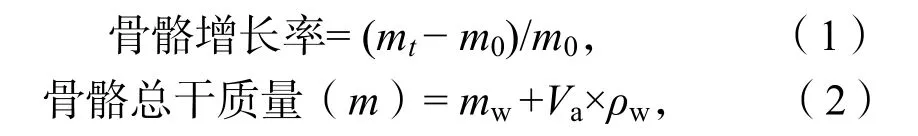
式中,m0為珊瑚的初始均干質量(g),mt為珊瑚的終末干質量(g),mw=實測樣品浮力質量(g),ρw為海水密度,Va為樣品體積。
1.4.2珊瑚蛋白質和脂肪分析 用洗牙器沖洗珊瑚獲組織勻漿液,粗蛋白含量采用凱氏定氯法(GB/T6432-2018)、脂肪含量使用索氏抽提法(GB/T6433-2006)測量,使用食品安全國家標準(GB 5009.168-2016)方法檢測脂肪酸組成。
1.5 統計分析
采用SPSS20.0 軟件對珊瑚骨骼重量、蛋白質、脂肪以及脂肪酸等數據進行統計分析,平均值±標準誤(Mean±SE),顯著性檢驗采用單因素方差分析(one-way ANOVA),并采用Duncan 多重比較檢驗。設顯著性水平α=0.05,極顯著水平β=0.01。
2 結果
2.1 清餌情況
表2 為盾形陀螺珊瑚對鹵蟲、輪蟲、小球藻的攝食情況,珊瑚對這三種浮游生物都表現出一定的攝食傾向,說明這三種浮游生物都有開發為盾形陀螺珊瑚活體餌料的潛力。
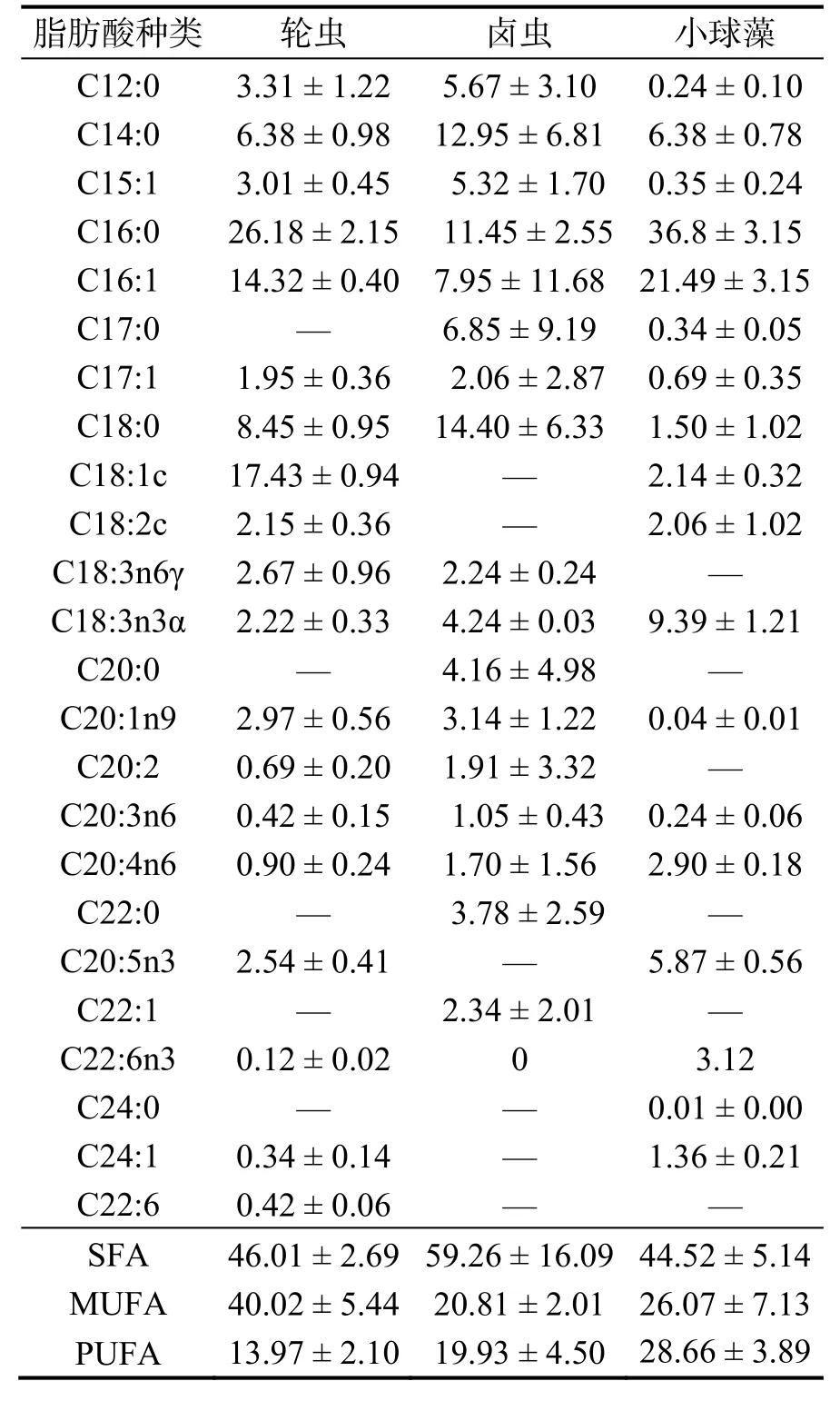
表1 輪蟲、鹵蟲、小球藻脂肪酸組成及其質量分數Table 1 Fatty acid composition and mass fraction of rotifers,artemia and chlorella %

表2 珊瑚對三種餌料的攝食情況Table 2 Feeding rate of rotifers,artemia and chlorella
2.2 鈣化率
表3 為各處理組喂食45 d 后的生長情況,異養營養的攝入顯著提高了盾形陀螺珊瑚的鈣化率,所有喂食組的珊瑚骨骼質量增長均大于對照組(P<0.01),喂食輪蟲的珊瑚小塊骨骼質量增長最多(P<0.05),為(1.43 ± 0.24)g。鹵蟲組和小球藻組的珊瑚骨骼質量增長分別為(0.66 ± 0.09)g 和(0.18 ±0.06)g,對照組中珊瑚的增長質量為(0.16±0.04)g。各組珊瑚在不同的餌料喂養和不喂養的條件下均能有100%的成活率。在不同喂養條件下珊瑚的骨骼增長情況存在顯著差異(P< 0.05),輪蟲喂養條件下的骨骼增長率最高,達到4.27%。輪蟲組和鹵蟲組與小球藻組和對照組骨骼增長率存在顯著差異(P<0.05),小球藻和對照組間骨骼增長率并無顯著差異(圖1)。

表3 不同喂食下珊瑚生長情況Table 3 Coral growth under different feedings
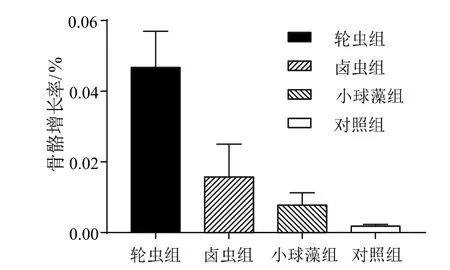
圖1 不同餌料喂食下珊瑚的骨骼增長率Fig.1 Coral bone growth rate under different diets
2.3 蛋白質質量分數
輪蟲喂食下陀螺珊瑚的蛋白質質量分數與其余各處理組均存在極顯著影響(P< 0.01),輪蟲組的珊瑚蛋白質質量分數最高(180 ± 9.64)mg/g,比小球藻組(112 ± 10.44)mg/g 和鹵蟲組(108.67 ±3.06)mg/g 分別高出60.7%和66.7%,對照組的珊瑚蛋白質質量分數最低,為(61.67 ± 8.96)mg/g(圖2)。其中輪蟲組珊瑚的蛋白質質量分數最高,這與該組測得最高骨骼生長率結果相吻合。
2.4 脂肪質量分數
在三個處理組中,輪蟲組的珊瑚脂肪質量分數(70.44 ± 9.36)mg/g 顯著高于其他處理組(P<0.05),其次是鹵蟲組(59.67 ± 1.53)mg/g 和小球藻組(41.03 ± 5.33)mg/g,脂肪質量分數最低的是對照組的珊瑚,為(30.97 ± 1.27)mg/g(圖2)。
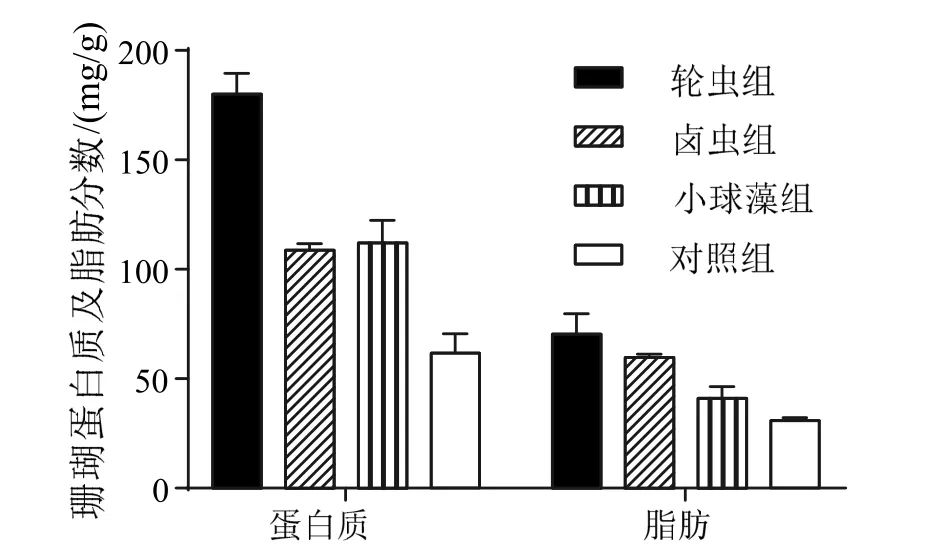
圖2 不同喂食組珊瑚蛋白及脂肪分數對比Fig.2 Coral protein and fat content comparison of different feeding groups
由圖3 可知,蛋白質質量分數在各處理間大都存在極顯著差異,小球藻組及鹵蟲組的蛋白質差異不顯著。輪蟲組和鹵蟲組、小球藻組與對照組脂肪質量分數差異不顯著,其余各處理間脂肪質量分數均存在顯著差異。
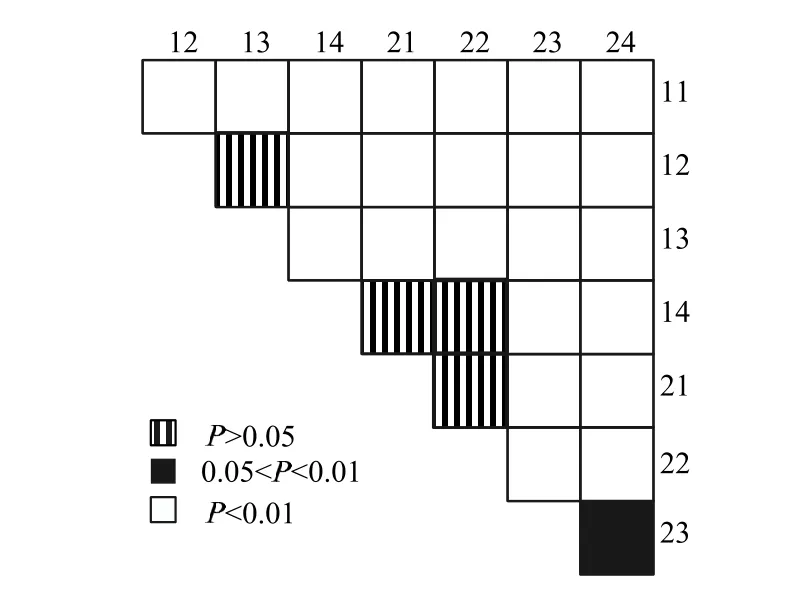
圖3 不同喂食組蛋白及脂肪質量分數anova 分析Fig.3 Anova analysis of protein and fat content in different feeding groups
2.5 脂肪酸組成
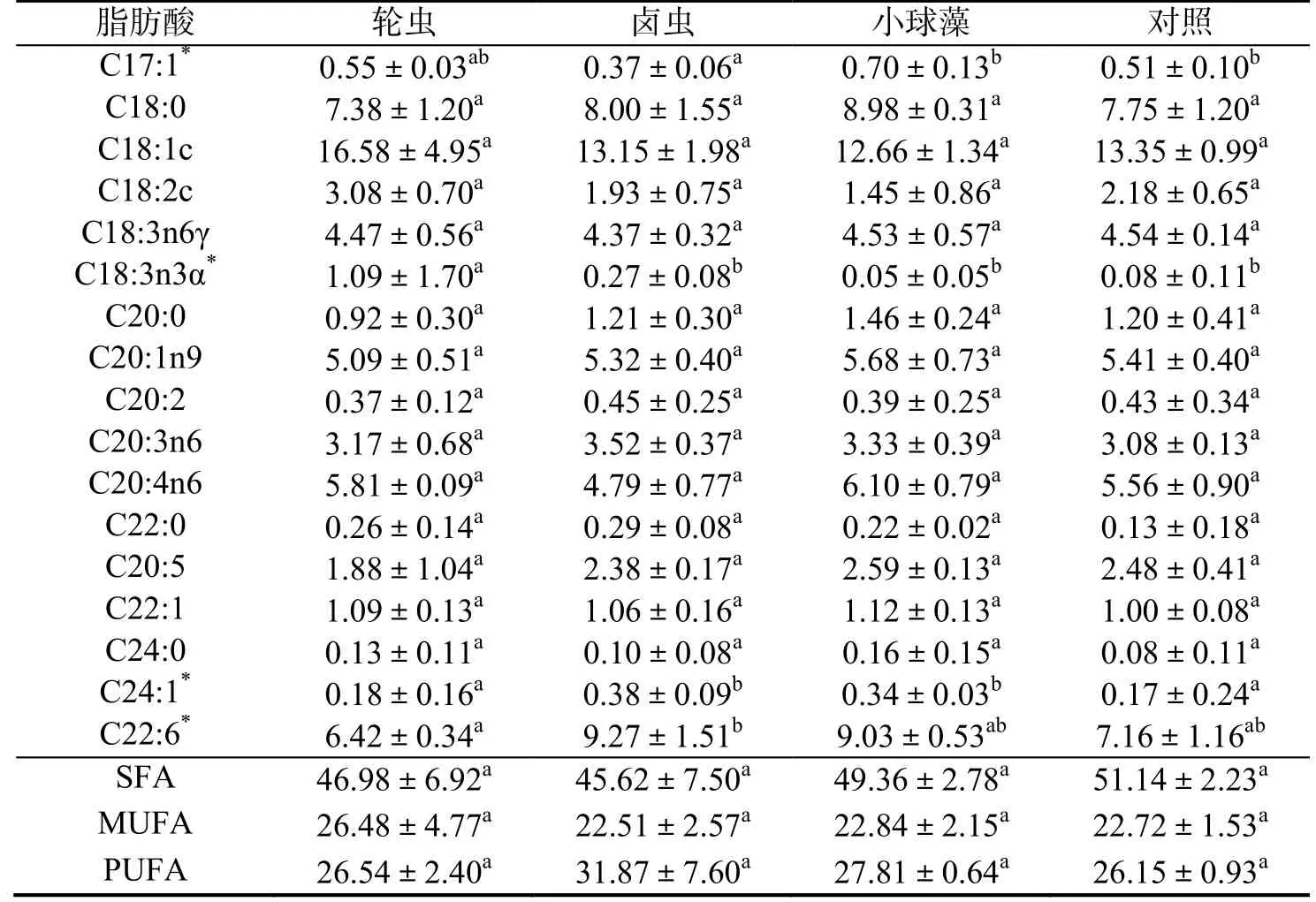
由表4 可知,不同餌料喂食下的珊瑚脂肪酸組成存在差異,其中輪蟲組與其他實驗組間存在較大差異。輪蟲組中珊瑚的C12:0、C17:0、C17:1、C22:6、C18:3n3α 這幾種脂肪酸的比例顯著高于其他實驗組(P< 0.05),C18:2c 脂肪酸的比例高于其他實驗組,C24:1、C22:6 的比例顯著低于其他實驗組(P< 0.05),這可能是因為珊瑚骨骼和組織的快速生長消耗了更多的脂肪酸,輪蟲組的飽和脂肪酸含量低于小球藻組以及對照組,表明輪蟲能夠為珊瑚提供更多的不飽和脂肪酸,但同時也存在C24:1、C22:6 等不飽和脂肪酸供應不足的問題。

表4 不同喂食下珊瑚脂肪酸組成及其質量分數Table 4 Fatty acid composition and mass fraction under different diets %
續表4(Continued)
3 討論
了解珊瑚的營養需求對進一步推進珊瑚人工養殖和珊瑚礁修復有著重要意義。本研究表明,餌料類型會影響珊瑚的營養組成,投喂合適餌料會使珊瑚的蛋白、脂肪含量上升,從而加速其鈣化過程。
3.1 餌料對珊瑚鈣化率、蛋白質和脂肪含量的影響
高蛋白水平是活躍生長的標志[16],在本實驗中,盾形陀螺珊瑚在三種喂食條件下的生長性能存在顯著差異,喂食合適的餌料能幫助珊瑚獲得更高的生長性能。高鈣化率往往伴隨著高蛋白水平,這說明珊瑚的鈣化速率與外界蛋白質輸入息息相關,蛋白質的合成有助于提升組織生長和鈣化速率[17],蛋白質和氨基酸會被生物體用來生成新的蛋白,這一過程需要生物體定期攝入蛋白質。當珊瑚體內蛋白含量不足時,蛋白質被從不重要部位轉移到重要部位以維持基本的生理需求[18]。從食物中獲取蛋白質可以提高珊瑚的蛋白質水平,從而使得珊瑚有更多可分配的蛋白質用于鈣化過程。Oomori 等[19]的實驗證明了富含天冬氨酸的蛋白在生物礦化調控過程中的重要性,而該種氨基酸主要的來源是異養而非自養[20]。已經有研究證實珊瑚的蛋白質濃度隨進食而增加[21],Connolly 等[22]研究了喂食對白化珊瑚蛋白質含量的影響,結果表明喂食和未喂食珊瑚的蛋白質含量差異很大,喂食的珊瑚蛋白質顯著高于未喂食的珊瑚,這些結果說明珊瑚可以直接從食物中獲取蛋白質以滿足自身需求。脂肪是餌料的重要組成部分,是維持機體生長和發育重要來源[23]。餌料的蛋白和脂肪的供應與珊瑚生長有著直接關系,充足的能量儲備可以為珊瑚抵御環境變化提供幫助[24]。Tosla 等[25]的實驗證明喂食條件下珊瑚的脂肪含量會顯著增加,增加的量與投食餌料所含脂肪量成正比。在本實驗中,脂肪含量在各實驗組也有顯著差異,喂食浮游動物(輪蟲、鹵蟲)的珊瑚脂肪含量顯著高于喂食浮游植物和不喂食的珊瑚,這與Al-Moghrabi 等[21]的研究結果相似。異養營養對珊瑚鈣化的作用不僅表現在體成分的變化,異養營養的輸入也會提升在輪蟲喂養下珊瑚表現出高的蛋白和脂肪儲備,這將有利于珊瑚快速增長。
3.2 盾形陀螺珊瑚對不同粒徑大小餌料可接受性
目前大多數珊瑚對浮游生物的攝食率和自然條件下的食性研究很少,由于許多室內實驗中浮游動物物種組成受人為因素影響,人們對珊瑚攝食的選擇性知之甚少,珊瑚對不同浮游生物的捕食能力很大程度上與浮游生物的運動能力相關,確定珊瑚的食性對人工養殖珊瑚和珊瑚移植工作大有裨益。在本實驗中,通過清餌率的計算確認了盾形陀螺珊瑚對上述三種活體餌料的攝食情況,結果表明上述三種浮游生物均是盾形陀螺珊瑚的捕食對象。在兩種浮游動物(輪蟲、鹵蟲)中,盾形陀螺珊瑚對輪蟲有較高的攝食率。與Hii 等[26]的實驗相比,盾形陀螺珊瑚對鹵蟲的捕食率比Galaxea fascicularis低很多。不同珊瑚的捕食方式各不相同,Lewis 等[27]指出,Pocillopora只使用觸手取食,而Acropora austera既使用觸手取食,又使用黏液取食。作者推測鹿角珊瑚更適合捕捉體積小、運動能力較弱的浮游生物。在本實驗中,僅在鹵蟲喂食過程中發現盾形陀螺珊瑚有大量黏液分泌的現象,在小球藻及輪蟲喂食過程中均未發現這一現象,推測該種珊瑚在捕食不同粒徑大小的食物時會使用不同捕捉方式。
海洋微藻富含DHA、EPA 等海洋動物必需脂肪酸,能夠為海洋生物提供充足的不飽和脂肪酸補充。Leal 等[28]的研究表明許多珊瑚都存在攝食藻類的現象,尋找合適藻類餌料可能可以幫助陀螺珊瑚獲得更高的生長率。本實驗同時也證明了盾形陀螺珊瑚對微藻捕食行為的存在,鹵蟲組和小球藻組蛋白、脂肪濃度顯著高于對照組且濃度接近,但小球藻組卻未顯示出高于對照組的鈣化率,我們推測鹵蟲實際上為珊瑚提供了更多的蛋白質和脂肪,這兩種物質均已在骨骼和組織的快速生長過程中被消耗。小球藻組相對較低的鈣化率可能是因為珊瑚消化這種食物需要消耗比其他兩種餌料更多的能量,故而降低了該組的鈣化速度。這表明盾形陀螺珊瑚雖然可以捕食小球藻,但由于可能存在的適口性、消化吸收率等問題,小球藻并不適合作為盾形陀螺珊瑚的活體餌料,所以珊瑚餌料的開發除需要了解珊瑚對不同餌料的捕食情況,還需要進一步探究珊瑚對不同餌料的消化速率。Houlbrèque 等[29]認為水螅體大小決定了珊瑚的捕食傾向,在本研究中,輪蟲是最適合盾形陀螺珊瑚的活體餌料。相對于Galaxea fascicularis有著更大螅體的盾形陀螺珊瑚卻更傾向于捕食體積較小的輪蟲,確定不同珊瑚的最佳餌料不能僅以珊瑚螅體大小為依據,更多地需要關注其捕食方式及觸手活躍性,根據不同珊瑚的捕食差異性投喂不同類型的餌料。此外,Wijgerde等[30]指出喂食浮游動植物混合物后鹿角珊瑚的生長率比饑餓狀態下高8 倍,珊瑚捕食浮游植物的行為在Pocillopora中也有發現[31]。這些結果表明,需要針對不同珊瑚屬的捕食特點以及餌料適應性篩選合適的活體餌料用于珊瑚人工養殖。在Forsman等[32]的實驗中,人造餌料喂食下珊瑚的生長速率比喂食活體餌料低,且相對于人造餌料,鹵蟲、輪蟲等活體餌料更經濟實惠。另外,由于個體大小與珊瑚移植后的存活率成正比,確定最佳餌料可以極大地縮短珊瑚小塊的培育周期,這對利用無性繁殖技術恢復退化的珊瑚礁有巨大的幫助。
3.3 餌料對珊瑚脂肪酸分布的影響
脂肪酸在海洋生物能量儲備、個體發育和激素調節過程中起著非常重要的作用[33],對于珊瑚餌料的篩選,除了優化珊瑚餌料的選擇,還需要了解珊瑚對脂肪酸的需求。脂肪酸為珊瑚生長和繁殖提供新陳代謝能量,是一系列基本代謝過程所必需的,如細胞脂質合成、脂質運轉、二十烷類化合物的產生和膜流動性[34]。C20:5 (n-3) (EPA) 和C22:6(n-3) (DHA)是兩種非常重要的脂肪酸,可以調節生物體內的脂質代謝,并有促進生長發育、降低膽固醇以及增加高密度脂蛋白的作用[35]。異養營養為珊瑚供給脂肪酸的途徑有直接和間接兩種方式,直接方式為珊瑚從食物中獲取所需的脂肪酸,間接方式下珊瑚可以通過將食物有機物轉化和促進共生蟲黃藻光合作用來獲得一些食物中含量較低的脂肪酸[2]。對于珊瑚而言,恢復能力對于珊瑚耐受壓力并在壓力事件(如移植)后迅速恢復至關重要。有關研究強調了多不飽和脂肪酸在幫助珊瑚從壓力中恢復的重要性[36-37]。喂食輪蟲條件下,C18:1c、C18:2c 這兩種脂肪酸比例皆高于其他實驗組,這兩種脂肪酸被認為是珊瑚從蟲黃藻中獲得的[38],這兩種脂肪酸的增多說明異養營養會增強共生營養,異養營養的輸入可以為共生蟲黃藻提供更多營養,通過增加蟲黃藻的密度使其光合速率提升[19],進而獲得更多的光合產物。亞麻酸(C18:3n3)是合成n3 和n6 多不飽和脂肪酸的前體,珊瑚可以通過自養和異養兩種途徑獲得這種脂肪酸[39],輪蟲組C18:3n3 比例的升高說明珊瑚正在快速地進行細胞分裂,這與該組測得的最高鈣化率相吻合[20]。不同餌料投喂下珊瑚的多種脂肪酸(C12:0、C17:0、C17:1、C18:3n3、C22:6)比例發生變化,輪蟲組、鹵蟲組中C12:0、C17:1 兩種脂肪酸的含量顯著高于其他兩個實驗組,這與這兩種餌料中這些脂肪酸比例相對較高的結果一致,另外,小球藻喂養條件下珊瑚的DHA(C22:6)的含量顯著增加,這些結果與李磊等[28]的實驗結果吻合,這說明投喂餌料的脂肪酸分布會改變珊瑚體內脂肪酸的分布。EPA 和C20:4n6 在各處理組沒有顯著差異,這些脂肪酸是已知的二十烷基酸前體,它們對許多生理過程至關重要,包括色素沉著和免疫功能[40],這表明由于它們不可或缺的性質,珊瑚或蟲黃藻合成的這些脂肪酸會被珊瑚選擇性保留[41]。本實驗中輪蟲組的DHA、EPA 顯著低于其他幾個實驗組,說明在高速生長的情況下,不進行營養強化的輪蟲體內的DHA 和EPA 不足以補充珊瑚體內這兩種脂肪酸的消耗。通過人工方式為輪蟲補充不飽和脂肪酸是解決輪蟲脂肪組成不均勻的有效方式,王斌等[42]對比了不同營養強化方式的輪蟲喂食下凡納濱對蝦的脂肪酸組成情況,結果表明輪蟲體內的脂肪酸組成對凡納濱對蝦的脂肪酸組成有直接影響。所以,尋找合適的方式強化輪蟲以對珊瑚進行特定的脂肪酸補充有利于人工養殖珊瑚的進一步發展。
4 結 論
本實驗以徐聞地區優勢珊瑚屬——盾形陀螺珊瑚為研究對象,探究三種餌料喂食下珊瑚體成分及鈣化率的變化,得到了結論:異養營養的輸入能夠顯著地提升珊瑚的鈣化率、蛋白質及脂肪含量。在選取的三種活體餌料中,輪蟲喂食下珊瑚的各個指標均高于其他餌料,是人工養殖盾形陀螺珊瑚較佳的活體餌料。輪蟲存在部分脂肪酸含量不足的現象,需進一步對其進行脂肪酸強化為珊瑚提供更充足的脂肪酸補給。未來需要進一步研究輪蟲喂食的合適密度及頻率,再輔以相應的營養強化手段,以找到最佳的盾形陀螺珊瑚飼喂方案。
