低溫脅迫對軍曹魚幼魚脂代謝相關生理生化的影響
2021-06-12 00:23:52蔡潤佳黃建盛張健東潘傳豪王忠良謝瑞濤湯保貴
廣東海洋大學學報 2021年3期
蔡潤佳,張 靜,黃建盛,陳 剛,張健東,潘傳豪,王忠良,謝瑞濤,湯保貴,3
(1.廣東海洋大學水產學院,廣東 湛江 524088;2.南方海洋科學與工程廣東省實驗室(湛江),廣東 湛江 524025;3.廣東省水產經濟動物病原生物學及流行病學重點實驗室,廣東 湛江 524088;4.廣東恒興飼料實業股份有限公司,廣東 湛江 524000;5.農業農村部華南水產與畜禽飼料重點實驗室,廣東 湛江 524088)
水溫是影響魚類生存最重要環境因子之一,與魚類多種生命活動密切相關[1]。1971 年,Lyons 等[2]指出,低溫可導致膜脂從液相向凝膠相轉變,并導致生物膜內物質大量向膜外泄露,打破生物膜內外離子平衡。自此,低溫與生物脂代謝的關系廣受關注。在魚類方面,邵彥翔等[3]發現,隨水溫不斷降低,云紋石斑魚 (Epinephelus moara,♀) × 鞍帶石斑魚 (E.lanceolatus,) 雜交后代血清甘油三酯(TG)呈先升后降再升趨勢。He 等[4]研究表明,低溫脅迫對吉富羅非魚(GIFTOreochromis niloticus)肌肉脂肪酸不飽和度有明顯提高作用,對增強細胞膜流動性有較大作用。Alba 等[5]應用轉錄組測序技術對低溫脅迫下的金頭鯛(Sparus aurata)進行肝臟轉錄組分析,結果發現肝臟在低溫下產生的大量差異基因富集在脂代謝的相關通路上。可見,調節脂代謝是魚類應對低溫脅迫的重要措施之一。
軍曹魚(Rachycentron canadum)屬于鱸形目(Perciformes)軍曹魚科(Rachycentridae)軍曹魚屬(Rachycentron),適宜溫度范圍為21~ 31 ℃,屬于暖水性魚類,分布于地中海、印度-太平洋(東太平洋除外)等海域,在我國南方部分海域也有少量分布[6]。軍曹魚對低溫的不耐受性導致其在我國只能在南方養殖。目前,關于軍曹魚在低氧脅迫[7]、鹽度適應[8]、營養[9]等已有研究報道,而軍曹魚溫度相關研究主要涉及溫度對生長和抗氧化酶活性的影響[10-11],對低溫條件下的軍曹魚生理調節機制了解極為有限。筆者研究低溫條件下軍曹魚的脂代謝,旨在補充軍曹魚在低溫條件下生理調節機制資料,為進一步培育軍曹魚耐低溫品種提供參考。
1 材料和方法
1.1 實驗動物
廣東海洋大學魚類種子工程與養殖實驗室人工繁育的軍曹魚幼魚,體質量(211.17±9.51)g,體長(32.68±0.78)cm,于湛江恒興南方海洋科技有限公司6 個300 L 養殖桶中暫養5 d。設置低溫組和常溫對照組,每組設3 個平行組。隨機取魚90 尾,每組15 尾。常溫對照組采用流水養殖且不間斷充氣,保持溶氧5 mg/L 以上,水溫(30.5±1.0)℃,每天08:00 和16:00 飽食投喂。低溫組采用冰塊(以密封瓶裝冰)降溫法以2 h/℃勻速降溫至(20.0 ± 0.5)℃,幼魚由于低溫不攝食故不投喂飼料,實驗期間靜水養殖,每天更換50%等溫新鮮海水,不間斷充氣,使溶氧保持在5 mg/L 以上。每天用水質檢測試劑盒監測水質,保持6 個桶氨氮質量濃度小于0.02 mg/L,亞硝酸鹽質量濃度小于0.01 mg/L,pH 7.4~ 7.8。
1.2 樣品采集
根據預實驗結果,軍曹魚幼魚在20 ℃下半致死時間為7 d,故正式實驗周期設為7 d。在1、4、7 d時每桶分別隨機采樣3 尾。采樣前停飼24 h,采樣時用丁香酚麻醉后于尾靜脈處采血;取部分肝臟樣品,用體積分數4%多聚甲醛溶液固定,制作切片;肌肉、部分肝臟和腹腔脂肪樣品分裝入標記有采樣時間、實驗處理方式和組織來源的封口袋;血液置4 ℃冰箱12 h,以3 500 r/min、15 min、4 ℃條件離心,血清于-80 ℃冰箱保存;切片樣品常溫保存;其余樣品-80 ℃冰箱中保存,用以檢測脂肪酸組成。
1.3 生化指標分析
采用全自動生化分析儀(深圳雷杜生命科技,Chemray800)測定血清甘油三酯(TG)、總膽固醇(T-CHO)、低密度脂蛋白膽固醇(LDL)、高密度脂蛋白膽固醇(HDL)含量;血清丙二醛(MDA)含量、總抗氧化能力(T-AOC)均用酶標檢測儀(BioTeK,Epoch)根據南京建成生物工程研究所的商業試劑盒說明書測定。
1.4 肝臟組織學觀察
參考蕭培珍[12]方法制作肝臟切片。樣品經磷酸鹽緩沖液(PBS)沖洗、梯度蔗糖溶液脫水、包埋、冷凍切片機切片、恢復室溫、油紅O 染色、封片等步驟,用體視鏡(Leica M205FCA)觀察并采集圖像,用Image J 軟件統計切片脂滴面積。
1.5 肝臟、肌肉、腹腔脂肪組織脂肪酸組成分析
肝臟、肌肉和腹腔脂肪組織(IPF)用烘箱60 ℃烘干至恒重,用粉碎機粉碎樣品,每個樣品稱取50 mg,分別加入2 mL 體積分數0.5%的硫酸-甲醇溶液,于60 ℃水浴鍋中溫浴1 h,再分別加入2 mL 正己烷混合萃取,靜置,取上層清液加入適量無水硫酸鈉,采用氣相色譜法進行脂肪酸組成分析(委托恒興質量檢測中心進行)。
1.6 統計分析
實驗數據用平均值±標準差表示。采用SPSS 19.0 軟件的T檢驗對數據進行差異分析。當P<0.05時表示差異顯著,P<0.01 時表示差異極顯著。
2 結果與分析
2.1 低溫脅迫對幼魚血清脂代謝相關指標的影響
低溫條件下飼養1 d 時,軍曹魚幼魚血清TG含量顯著高于對照組(P<0.05),而4 d、7 d 時,低溫組血清TG 含量均升高,與同期對照組差異極顯著(P<0.01)(圖1_a)。軍曹魚血清T-CHO 含量在低溫條件下呈先降后升再降的趨勢,1 d 時顯著低于對照組(P<0.05),4 d 時顯著高于對照組(P<0.05),7 d 時則下降至與對照組水平(P>0.05)(圖1_b)。低溫脅迫下軍曹魚血清HDL 在1 d 和4 d 時含量無明顯變化(P>0.05),7 d 時則顯著下降(P<0.05)(圖1_c)。血清LDL 含量在低溫脅迫1 d 時與對照組無顯著差異(P>0.05);4 d 時升高并顯著高于對照組(P<0.05);7 d 時含量進一步升高,與對照組差異極顯著(P<0.01)(圖1_d)。
2.2 低溫脅迫對幼魚血清氧化指標的影響
低溫飼養1 d 時,軍曹魚幼魚血清MDA 含量極顯著升高(P<0.01),4 d 時最高,7 d 時下降,但仍顯著高于同期對照組(P<0.05)(圖2_a)。血清T-AOC 整體呈下降趨勢,1 d 時與對照組無顯著差異(P>0.05),4 d 時顯著降低(P<0.05),7 d 時進一步降低,極顯著低于對照組(P<0.01)(圖2_b)。
2.3 低溫脅迫對幼魚肝臟脂滴分布的影響
常溫條件下,軍曹魚幼魚脂滴均勻分布,無明顯空白區域,且脂滴邊緣清晰,融合現象不明顯(圖3)。低溫條件下,1 d 時,脂滴融合,形成形狀不規則的較大脂滴,但無明顯空白區;4 d 時,脂滴融合現象有所減少,大部分脂滴邊緣清晰;7 d 時,出現較多空白區域,脂滴界限模糊,普遍融合成片。統計肝臟油紅O 染色切片脂滴面積,低溫組脂滴面積在1、4、7 d 時均極顯著高于對照組(圖3_g)。
2.4 低溫脅迫對幼魚肝臟、腹腔脂肪和肌肉脂肪酸組成的影響
軍曹魚幼魚應對20 ℃的低水溫環境時,肝臟、腹腔脂肪及肌肉組織脂肪酸代謝不同。表1 可見,肝臟脂肪酸中,飽和脂肪酸(Saturated fatty acid,SFA)7 d 內與對照無顯著差異(P>0.05),單不飽和脂肪酸(Monounsaturated fatty acid,MUFA)含量平均值不斷下降,但與對照組差異不顯著(P>0.05),多不飽和脂肪酸(Polyunsaturated fatty acid,PUFA)含量總體上不斷上升,7 d 時極顯著高于對照組(P<0.01),n-6 系列PUFA 7 d 時顯著提高(P<0.05)。表2 可見,腹腔脂肪脂肪酸中,SFA、MUFA含量在低溫脅迫時保持相對穩定(P>0.05),而PUFA 含量在1 d 時即顯著高于對照組(P<0.05),在4 d 和7 d 時含量進一步提高并極顯著高于同期對照組(P<0.01),其中n-3 系列PUFA 從1 d 時起即極顯著高于對照組(P<0.01),n-6 系列在1 d、4 d 時顯著高于對照組(P<0.05),并在7 d 時與對照組差異極顯著(P<0.01)。肌肉脂肪酸組成變化見表3,低溫脅迫下,低溫組三種脂肪酸占總脂肪酸比例與對照組無顯著差異(P>0.05)。
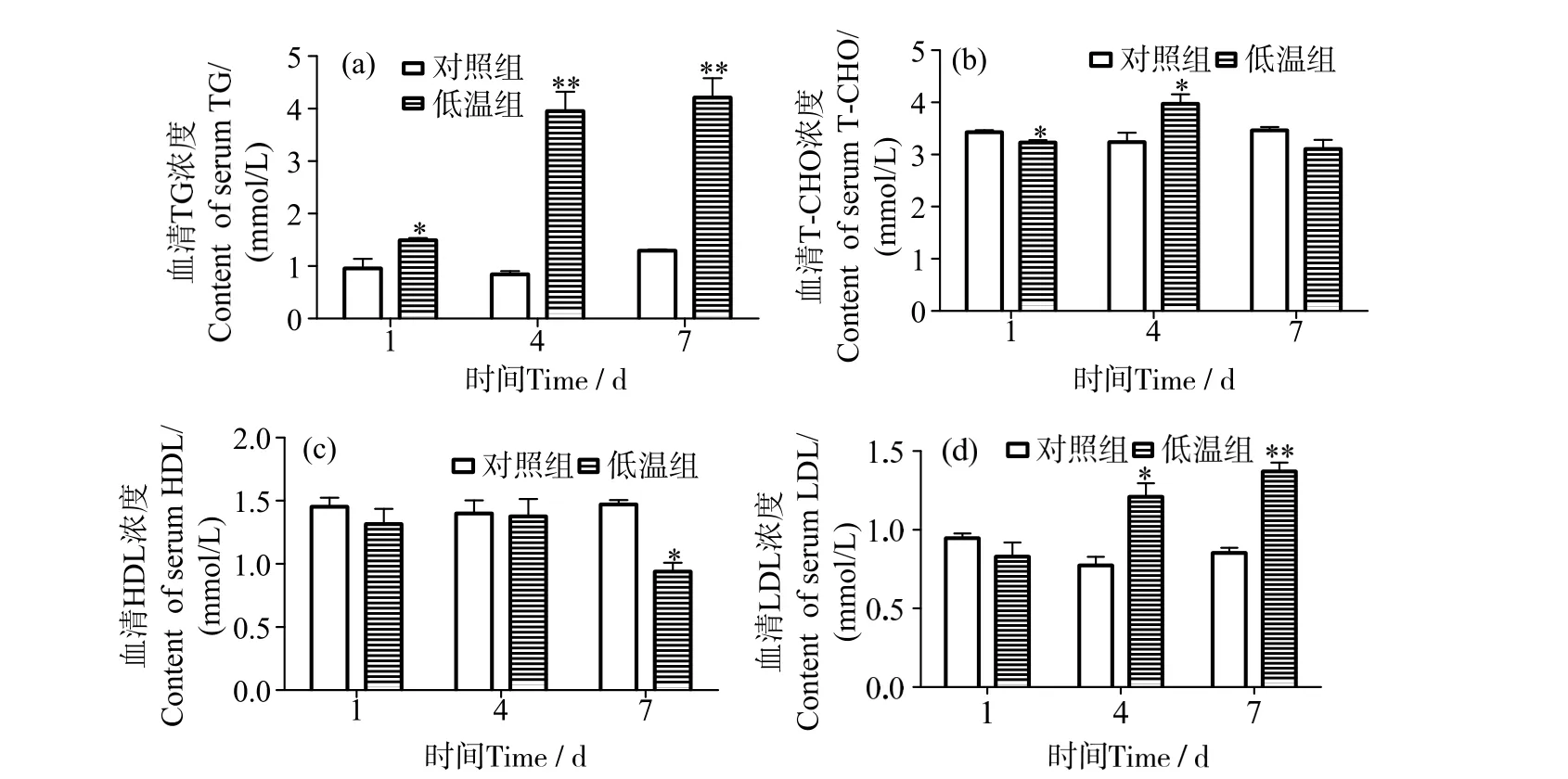
圖1 低溫脅迫對軍曹魚血清TG、T-CHO、HDL 和LDL 含量的影響Fig.1 Effects of low temperature stress on the content of TG,T-CHO,HDL and LDL in serum of cobia

圖2 低溫脅迫對軍曹魚血清MDA 和T-AOC 含量的影響Fig.2 Effects of low temperature stress on the content of MDA and T-AOC in serum of cobia

圖3 軍曹魚肝臟組織學觀察及脂滴面積統計Fig.3 Histological observation of cobia liver and lipid droplet area statistics

表1 低溫脅迫對軍曹魚肝臟脂肪酸組成的影響Table 1 Effects of low temperature stress on fatty acid composition of liver in cobia
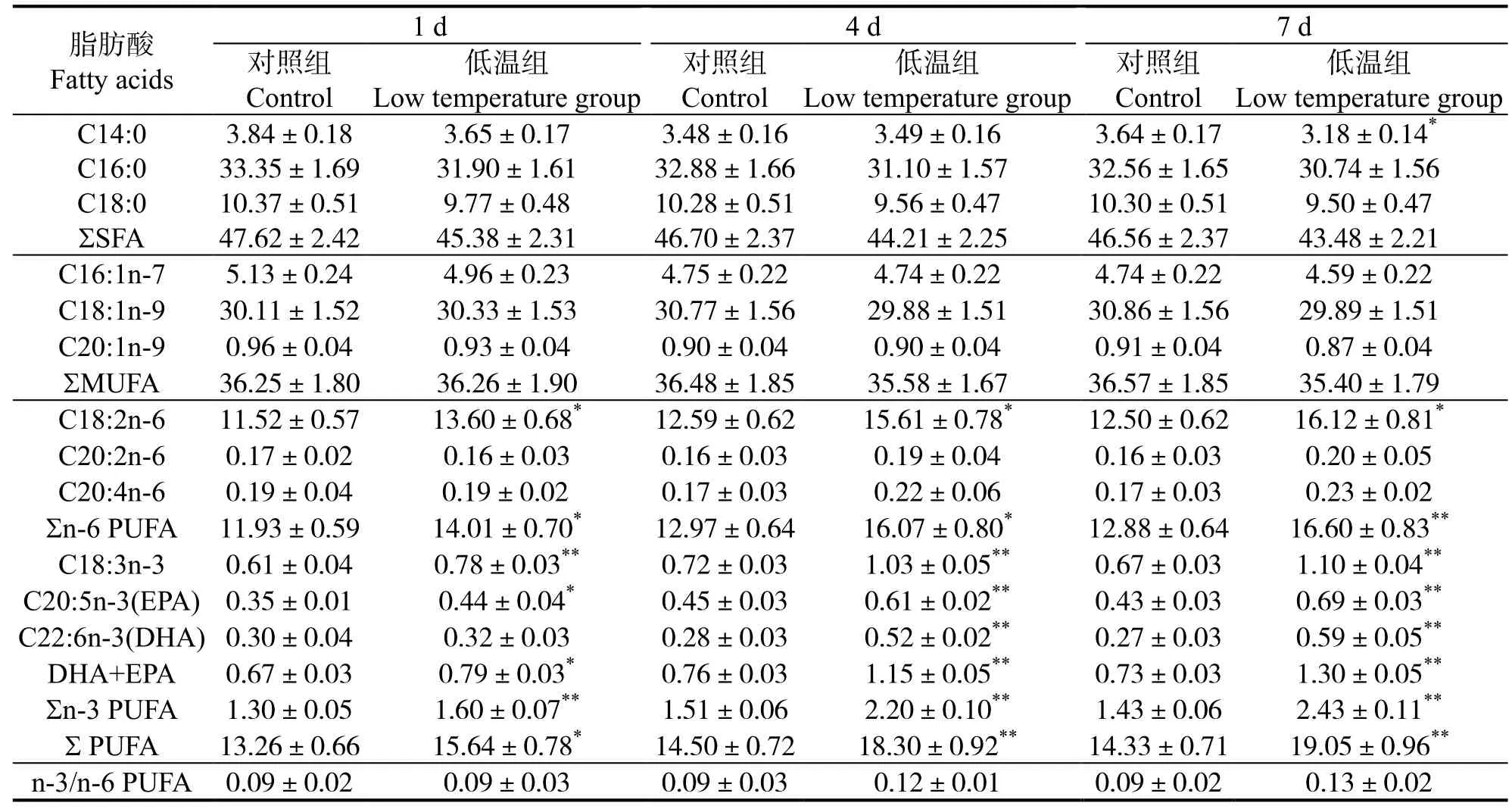
表2 低溫脅迫對軍曹魚腹腔脂肪脂肪酸組成的影響Table 2 Effects of low temperature stress on fatty acid composition of IPF in cobia
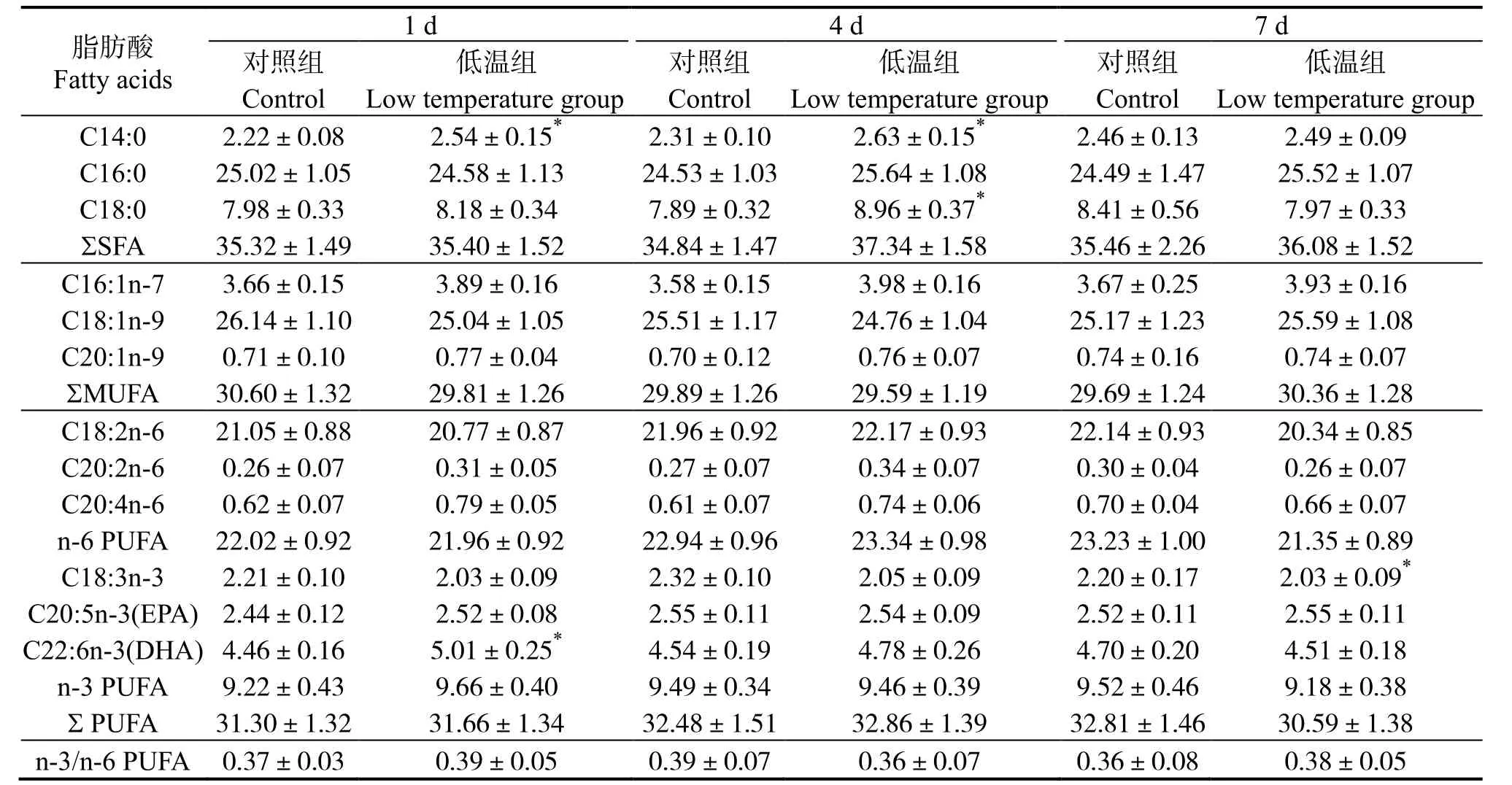
表3 低溫脅迫對軍曹魚肌肉脂肪酸組成的影響Table 3 Effects of low temperature stress on fatty acid composition of muscle in cobia
3 討論
3.1 低溫脅迫對軍曹魚幼魚血脂水平的影響
血液是動物物質代謝的重要載體,血液生化指標改變可指示機體多種生理變化[13]。血脂(TG 和T-CHO)水平可反映機體脂代謝情況。TG 是儲存在魚體的高效能量源,對魚體能量代謝有重要調節作用。本研究中,軍曹魚血清TG 在20 ℃脅迫1 d 時即顯著提高,4、7 d 時持續升高至與對照組差異極顯著。在吉富羅非魚[14]、暗紋東方鲀(Takifugu fasciatus)[15]、鯉魚(Cyprinus carpio)[16]等研究中也發現相似結果。魚類在低溫環境下無氧代謝增強,有氧代謝與脂代謝受抑制,使血清TG 消耗量減少,血清TG 增多[14]。但不同魚類血清TG 對低溫脅迫的響應不同。低溫脅迫褐藍子魚(Siganus fuscescens)導致魚體血清TG 呈先升后降趨勢[17],而斜帶石斑魚(Epinephelus coioides)則呈與褐藍子魚完全相反結果,有先降后升趨勢[18]。多鱗白甲魚(Onychostoma macrolepis)血清TG 在低溫脅迫前3 h 逐步上升,6 h 時顯著下降而后又逐漸上升[19]。這種物種間的血脂變化差異機制有待進一步研究。
血清膽固醇主要是細胞漿膜、類固醇激素等的合成原料,適宜血清膽固醇水平是保持細胞膜流動性的條件之一[20]。當血清T-CHO 含量減少時,可能會干擾細胞膜穩態從而影響多種生化過程[14],本研究中,軍曹魚血清T-CHO 在低溫脅迫1 d 時顯著降低,但在4 d 時升至極顯著升高,7 d 時又降至對照組水平,說明軍曹魚對血清T-CHO 的調節能力較強。在尼羅羅非魚(Oreochromis niloticus)[21]和斜帶石斑魚[20]中均發現低溫會減少血清T-CHO含量。可見,不同物種的血清T-CHO 對低溫脅迫的響應不盡相同。低溫會導致軍曹魚血清LDL 在4 d 和7 d 時升高,HDL 7 d 時降低。LDL 主要從肝臟向外周組織轉運內源膽固醇,而成熟的HDL 是機體從外周組織回收內源性膽固醇的載體,有清除血漿中膽固醇的作用。血清LDL、HDL 對低溫脅迫的響應表明軍曹魚脂代謝出現異常。
3.2 低溫脅迫對軍曹魚幼魚組織脂肪酸組成的影響
脂肪酸對生物體代謝有重要作用。吉富羅非魚在水溫28、22 ℃條件下飼養40 d 后,22 ℃組肌肉SFA、MUFA、PUFA 含量均與28 ℃組無顯著差異,僅n-6 系列PUFA 顯著高于28 ℃組[22];而吉富羅非魚在13 ℃低溫脅迫時,隨著脅迫時間的延長,肌肉中SFA 含量持續下降,MUFA 和PUFA 均呈先升后降趨勢[4];謝妙等[20]發現,斜帶石斑魚在20 ℃條件下脅迫15 d 后,肌肉SFA 含量顯著下降,MUFA和PUFA 含量則顯著上升。可見,組織脂肪酸組成對低溫脅迫的響應與物種、環境條件、脅迫時間等均有較大關系。本研究中,不同組織對低溫脅迫的響應也存在較大差異。在脂肪酸組成方面對低溫脅迫響應最小的組織是肌肉,各脂肪酸均與對照組無顯著差異。其次是肝臟,MUFA 含量有下降趨勢,而PUFA 含量則在7 d 時顯著高于對照組。對低溫脅迫響應程度最大的組織是IPF,其中n-3、n-6 系列PUFA,總PUFA 均不斷增加,在1 d 時即顯著或極顯著高于對照組。研究表明,PUFA 在維持細胞膜結構[23]、參與信號傳遞[24]等方面均有重要作用,所以提高PUFA 的占比可能是軍曹魚對低溫脅迫的適應性調節方式之一。對鯉魚[25]、尖吻鱸[26]等的研究均發現,低溫環境會提高不飽和脂肪酸比例,這是魚體為提高細胞膜流動性,而削弱低溫導致細胞膜流動性差的負面影響的結果。
3.3 低溫脅迫對軍曹魚幼魚氧化應激的影響
魚體在抗氧化系統調節下,體內活性氧(ROS)較低并保持動態平衡[27]。當魚體處于低溫環境時ROS 逐漸積累,過量的ROS 會促進脂肪酸尤其是不飽和脂肪酸發生過氧化反應,ROS 主要攻擊脂肪酸側鏈位置,可從脂質雙鍵中提取氫原子改變脂質結構[28],最終生成丙二醛和壬烯等脂質過氧化終產物,還會進一步生成過氧自由基,進一步加劇機體過氧化[29]。軍曹魚血清T-AOC 隨低溫脅迫時間的延長而顯著降低,抗氧化能力的降低會導致機體ROS 含量升高,從而阻礙正常脂代謝[30]。且魚體在低溫環境下占比增多的多不飽和脂肪酸比飽和脂肪酸更易積累ROS[31],這可能是軍曹魚在低溫脅迫1 d 時就迅速積累大量丙二醛的原因。肝臟是魚體脂代謝最重要調節器官,低溫可誘導暗紋東方鲀肝臟凋亡相關基因的表達[32],在虹鱒(Oncorhynchus mykiss)[33]、斜帶石斑魚[20]等物種中,低溫會導致谷丙轉氨酶和谷草轉氨酶顯著升高,指示肝細胞受損傷。本研究中,軍曹魚肝臟低溫脅迫后出現脂滴分布不均、融合、邊緣模糊、脂滴面積極顯著增加等現象,也說明了肝臟脂代謝活動出現異常。
3.4 低溫脅迫時不攝食與常溫條件下饑餓的血脂響應機制
低溫條件下,軍曹魚不攝食,與常溫饑餓脅迫同屬內源性營養代謝階段,但魚體響應兩種脅迫的血脂相關指標表現不同。胡文君[34]發現,黃鱔(Monopterus albus)于5 ℃環境時,與25 ℃正常投喂組相比THO 含量顯著降低,HDL 含量顯著升高,而TG 和LDL 含量則與對照組無顯著差異。而當黃鱔于25 ℃環境中進行饑餓脅迫時,THO、TG和HDL 含量均與25 ℃正常投喂組無顯著差異,而LDL 含量則顯著降低,表明血脂指標在低溫脅迫和常溫饑餓脅迫中,除血清TG 外,血清THO、HDL和LDL 等指標均有不同的變化趨勢,故推測低溫脅迫的脂代謝調節機制與饑餓脅迫不同,本研究低溫脅迫結果與停食的關系不大。
4 結論
軍曹魚幼魚在低溫脅迫過程中,機體抗氧化能力降低并積累大量MDA,從提高血清TG 含量、增加血清膽固醇含量(增加LDL、減少HDL)、調整肝臟和IPF 中的脂肪酸組成等方面響應低溫脅迫。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
中老年保健(2021年3期)2021-08-22 06:50:04
昆明醫科大學學報(2021年1期)2021-02-07 01:06:36
現代臨床醫學(2021年1期)2021-01-26 00:56:02
中華養生保健(2020年4期)2020-11-16 01:31:40
中西醫結合肝病雜志(2020年2期)2020-10-27 02:18:50
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
