海水酸化對(duì)珊瑚藻生長(zhǎng)和鈣化作用的影響
2021-06-12 00:23:44謝子強(qiáng)
廣東海洋大學(xué)學(xué)報(bào) 2021年3期
關(guān)鍵詞:生長(zhǎng)
聶 磊,謝子強(qiáng),彭 丹
(1.深圳信息職業(yè)技術(shù)學(xué)院環(huán)境工程學(xué)院,廣東 深圳 518268;2.廣東海洋大學(xué)深圳研究院,廣東 深圳 518120)
人類現(xiàn)代社會(huì)活動(dòng)高強(qiáng)度排放大量溫室氣體,導(dǎo)致大氣CO2濃度不斷升高,進(jìn)而引發(fā)諸如海平面升高、水循環(huán)異常、全球變暖、海洋酸化等系列重大環(huán)境問題,且相關(guān)問題自工業(yè)革命以來不斷加劇。據(jù)報(bào)道,全球海洋pH 值至21 世紀(jì)末期可能降至7.4,大氣CO2濃度可能加大到現(xiàn)在的5 倍[1]。珊瑚藻(coralline algae)屬于紅藻門(Rhodophyta)紅藻綱(Rhodophyceae)真紅藻亞綱(Florideophycidae)珊瑚藻目(Corallinales),是一類海洋常見大型紅藻,物種多樣性豐富,體內(nèi)含有大量碳酸鈣。珊瑚藻主要有殼狀、枝狀和瘤塊狀三種形態(tài),葉狀體堅(jiān)硬,又分有節(jié)和無節(jié)兩大類。珊瑚藻是海洋生物礁系統(tǒng)中最為重要的功能類群之一,在生物礁生態(tài)系統(tǒng)和礁體建造中具有廣泛和核心作用。開展珊瑚藻研究和保護(hù),對(duì)生物礁生態(tài)系統(tǒng)的可持續(xù)保護(hù)、以珊瑚為主的海洋受損生態(tài)系統(tǒng)的恢復(fù)等具有重大影響[2]。珊瑚藻具有鈣質(zhì)無機(jī)骨架,在生物礁鈣質(zhì)體的建造與維護(hù)方面不可替代,同時(shí)又被認(rèn)為是對(duì)海洋酸化最敏感的藻類生物之一,但目前國內(nèi)外關(guān)于海水酸化態(tài)勢(shì)下珊瑚藻的生理機(jī)制研究仍較少見,至于殼狀珊瑚藻(crustose coralline algae,CCA)的國內(nèi)生理生態(tài)研究更是空白[3]。本研究以南海海域典型的殼狀珊瑚藻擬中葉藻和枝狀珊瑚藻叉節(jié)藻為對(duì)象,比較分析海水酸化對(duì)于殼狀及枝狀珊瑚藻的生長(zhǎng)、光合色素積累、碳酸酐酶(CA)以及鈣化固碳的影響差異,以期為探索珊瑚藻,尤其殼狀珊瑚藻的生態(tài)機(jī)理以及珊瑚礁的生態(tài)系統(tǒng)保護(hù)提供理論支持。
1 材料與方法
實(shí)驗(yàn)采用的叉節(jié)藻(Amphiroa ephedraea)和擬中葉藻(Mesophyllum simulans)樣品于2018 年8 月采集于深圳大鵬灣潮間帶(114°36′N,22°25′E),新鮮采集的叉節(jié)藻,蒸餾水洗去泥沙及其他附著物,去除雜藻,選擇生長(zhǎng)旺盛,藻體完整的樣本滅菌處理3~4 次。以500 mL 錐形瓶作為培養(yǎng)容器,保持每瓶添加的叉節(jié)藻藻體質(zhì)量為(5.0±0.05)g。殼狀藻擬中葉藻連同原有生長(zhǎng)基石一起在錐形瓶中培養(yǎng)[4]。
1.1 不同培養(yǎng)條件的設(shè)置
實(shí)驗(yàn)設(shè)置3 個(gè)不同pH 值的培養(yǎng)條件:8.2、7.8和7.4,各設(shè)3 個(gè)平行組。pH 值使用經(jīng)標(biāo)準(zhǔn)液校正的pH 計(jì)(FE20,Mettler Toledo,Switzerland)測(cè)定。pH 值8.2 的處理為深圳大鵬灣自然海水,通過向自然海水充入瓶裝CO2氣體獲得7.8 和7.4 的 pH值,充氣時(shí)用pH 計(jì)即時(shí)監(jiān)測(cè)海水pH 值變化,當(dāng)pH 達(dá)到所需值時(shí)停止充氣,放入藻體進(jìn)行靜水培養(yǎng)。光照培養(yǎng)箱控制培養(yǎng)溫度為20 ℃,光照強(qiáng)度為100 μmol·m-2·s-1,光暗比為12 h∶12 h,每天搖動(dòng)藻體 2 次。培養(yǎng)過程監(jiān)測(cè) pH 值,90 d 后測(cè)定不同處理下珊瑚藻的各項(xiàng)生長(zhǎng)和生理生化指標(biāo)。
1.2 生長(zhǎng)分析
培養(yǎng)開始前和結(jié)束時(shí)用蒸餾水清洗藻體,其中擬中葉藻藻結(jié)皮用刀片輕輕刮下。用吸水紙吸干藻體表面,分別測(cè)定藻體的鮮質(zhì)量,按公式(1)計(jì)算出相對(duì)生長(zhǎng)率(RGR)[4]:
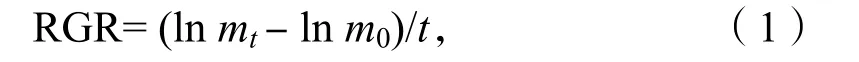
其中,m0為實(shí)驗(yàn)開始時(shí)海藻的鮮質(zhì)量,mt為實(shí)驗(yàn)t天后海藻的鮮質(zhì)量。t為實(shí)驗(yàn)中藻類培養(yǎng)時(shí)間,單位為d。
1.3 光合色素分析
采用分光光度法測(cè)定珊瑚藻葉綠素a(Chl a)、類胡蘿卜素(Car)和藻紅蛋白(PE)質(zhì)量濃度,取 0.1 g 左右的新鮮藻體經(jīng)丙酮研磨后浸提,4 000 r/min 離心10 min,取上清液分別在波長(zhǎng) 480 nm、645 nm和 663 nm 下測(cè)定提取液的光密度,然后再按照下列公式將濃度換算為樣品葉綠素 a 質(zhì)量濃度(單位:mg·L-1)和類胡蘿卜素質(zhì)量濃度(單位:μg·L-1),D代表上清液在相應(yīng)波長(zhǎng)下光密度[5]:
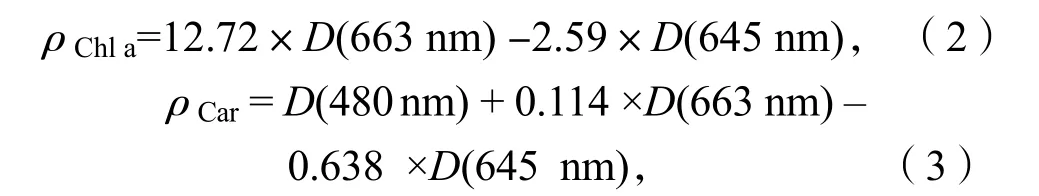
藻紅蛋白(PE)質(zhì)量濃度的測(cè)定:取 0.1 g 新鮮藻體在磷酸緩沖液(pH=6.8)研磨后浸提,經(jīng)4 000 r/min 離心10 min,取上清液分別在波長(zhǎng)455、564 和592 nm 波長(zhǎng)處測(cè)定光密度D[5]。PE 質(zhì)量濃度按公式(4)計(jì)算,單位為mg·L-1:
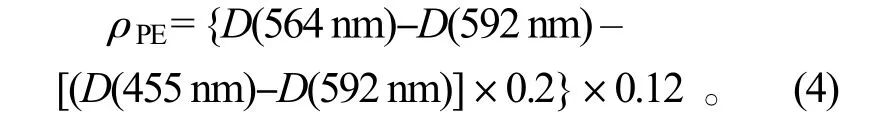
1.4 碳酸酐酶(CA)活性分析
稱取藻樣品0.5 g 置于預(yù)冷研缽中,在冰浴條件下加入3 mL 提取液(5 mmol/L 巰基乙醇,15 mmol/L 巴比妥緩沖液,pH=8.2)進(jìn)行研磨,用4層消毒紗布對(duì)研磨勻漿過濾,濾液在4 ℃環(huán)境中用離心機(jī)7 000 r/min 下離心5 min,取上清液(CA粗提液),4 ℃下冷藏待進(jìn)行CA 活性測(cè)定,以煮沸5 min 冷卻的部分上清液作為對(duì)照。采用pH 計(jì)法測(cè)定碳酸酐酶活性,分別在兩個(gè)冰浴條件下的小燒杯中依次加入15 mmol/L 巴比妥緩沖液、未煮沸或者煮沸的碳酸酐酶粗提液以及冰冷的CO2飽和水,記錄下pH 值由8.2 下降至7.4 所用的時(shí)間。研磨過濾后的固體殘?jiān)半x心后的沉淀物用于葉綠素a 濃度的測(cè)定。CA 活性的計(jì)算公式如下[6]:
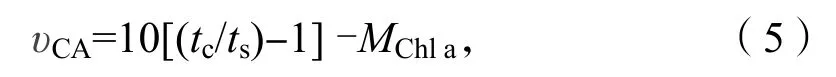
其中υCA代表碳酸酐酶活性,tc代表對(duì)照中pH 值從8.2 下降至7.4 所用的時(shí)間(min),ts代表未煮沸酶液pH 值從8.2 下降至7.4 所用的時(shí)間(min),MChla代表測(cè)定藻體的葉綠素量(μg)。
1.5 藻體中碳成分的含量測(cè)定
珊瑚藻樣品經(jīng)烘箱烘干(于50 ℃條件下)后研磨成粉末狀,準(zhǔn)確稱取1.0 mg 樣品用玻璃微纖維濾紙包裹,置于不密封的玻璃試管中,試管在濃鹽酸揮發(fā)形成的酸霧條件下暴露酸熏24 h,除去樣品中的顆粒無機(jī)碳(PIC),第二次烘干待測(cè)定,樣品有機(jī)碳含量由碳氮分析儀(multi N/C UV,Analytik Jena AG,Germany)測(cè)定。經(jīng)過酸熏處理的粉末樣品測(cè)定結(jié)果為珊瑚藻藻體顆粒有機(jī)碳(POC)含量,未經(jīng)酸熏處理的樣品測(cè)定結(jié)果為總的碳顆粒含量,減去POC 的量即為藻體顆粒無機(jī)碳(PIC)含量。
1.6 鈣化固碳速率和光合固碳速率的測(cè)定
不同培養(yǎng)條件下的珊瑚藻分別測(cè)定其在3 h 的時(shí)間間隔內(nèi)PIC 和POC 含量變化,以PIC 含量的增加速率表示藻的鈣化固碳速率(Cn),POC 含量的增加速率表示光合固碳速率(Pn)。
1.7 數(shù)據(jù)處理
實(shí)驗(yàn)數(shù)據(jù)采用SPSS18.0 分析后,根據(jù)單因素方差分析(one-way ANOVA)檢驗(yàn)數(shù)據(jù)差異顯著性,顯著性水平α設(shè)為0.05。
2 結(jié)果與分析
2.1 海水酸化對(duì)于珊瑚藻光合固碳速率、光合色素含量及碳酸酐酶活力的影響
珊瑚藻的光合作用是以CO2和為底物,把無機(jī)碳合成有機(jī)物的過程,可以用方程式表示為:CO2+H2O ? CH2O+O2;這是對(duì)CO2的一個(gè)吸收過程。在本研究中,通過向培養(yǎng)水體中充入高濃度CO2的方法降低海水pH 值,珊瑚藻的光合固碳速率(Pn)隨著pH 值的下降而逐步上升,其中pH7.4 條件下藻體的光合固碳能力要高于pH 7.8(圖1)。經(jīng)90 d 培養(yǎng)后,pH7.4 培養(yǎng)條件下的叉節(jié)藻光合固碳速率達(dá)到[(6.21±0.47)mg·g-1·h-1],較pH 7.8 條件下增加了6.15%,達(dá)到了pH 8.2 條件下的26.52%。殼狀珊瑚藻在pH 值降低狀況下的光合能力上升幅度絕對(duì)值不如枝狀珊瑚藻,但相對(duì)差異值則更為明顯。經(jīng)90 d 培養(yǎng)后,擬中葉藻在pH 7.8到 pH 7.4 之間的光合固碳效率上升幅度達(dá)到13.46 %(達(dá)到顯著水平),超過叉葉藻的上升幅度。
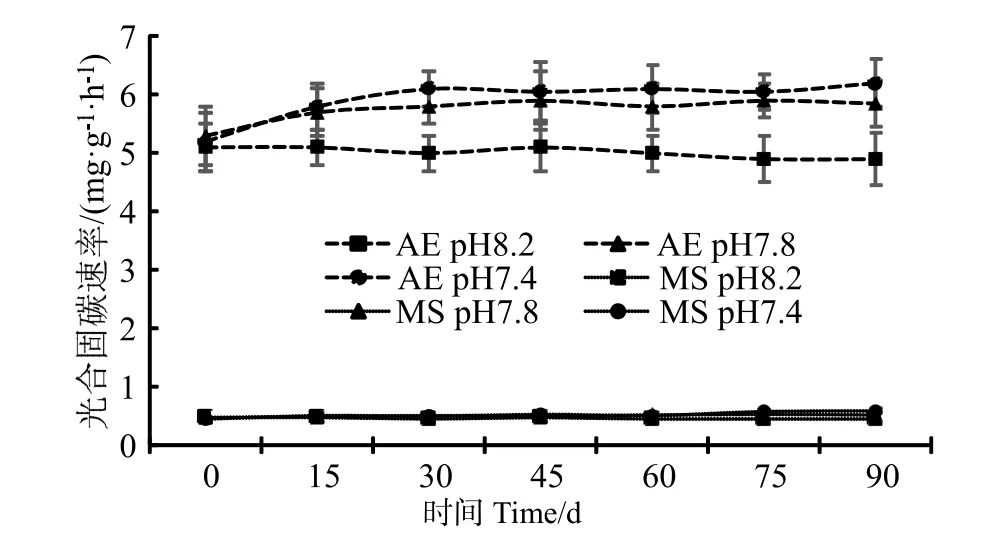
圖1 海水酸化處理對(duì)珊瑚藻光合固碳速率的影響Fig.1 The effects of seawater acidification on the rate of photosynthetic carbon fixation of coralline algae
不同pH 海水處理90 d 后測(cè)定珊瑚藻中的葉綠素a 質(zhì)量濃度,結(jié)果見圖2。與pH 8.2 的對(duì)照組比較可發(fā)現(xiàn),海水酸化處理下實(shí)驗(yàn)組的葉綠素a 與藻紅蛋白(PE)質(zhì)量濃度明顯降低,且隨著pH 值減低而不斷下降,在pH 7.8 和pH 7.4 處理下,叉節(jié)藻分別下降了29.52%和48.30%,而擬中葉藻葉綠素a 分別下降了22.72%和50.15%,均達(dá)到了顯著水平,藻紅蛋白質(zhì)量濃度的變化趨勢(shì)與此類似。在pH 7.8 和pH 7.4 的海水酸化處理下,叉節(jié)藻的類胡蘿卜素含量分別上升了21.52%、和23.47%,而和擬中葉藻則上升了24.33%和33.35%,珊瑚藻的類胡蘿卜素質(zhì)量濃度在海水酸化處理下的變化趨勢(shì)與葉綠素a 與藻紅蛋白(PE)相反。
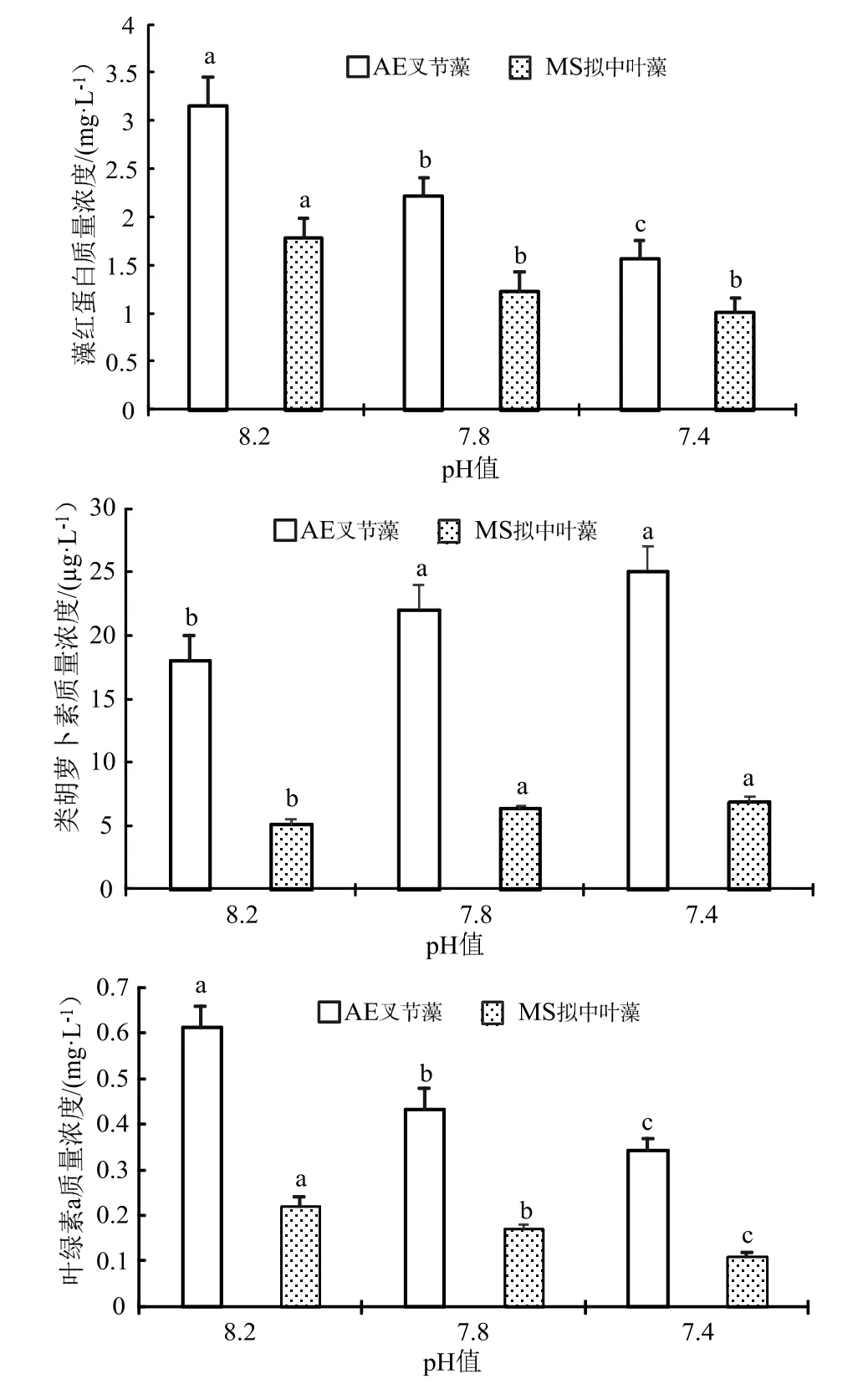
圖2 海水酸化處理對(duì)珊瑚藻光合色素含量的影響Fig.2 The effects of seawater acidification on the contents of photosynthetic pigments of coralline algae
作為與Rubisco 羧化酶(1,5-二磷酸核酮糖羧化酶)同樣屬于光合作用過程中催化(同化)無機(jī)碳的關(guān)鍵(限速)性酶,碳酸酐酶(CA)主要分布在珊瑚藻細(xì)胞內(nèi)的質(zhì)體、葉綠體中或者細(xì)胞質(zhì)膜表面,主要生理功能為可逆性催化CO2(碳酸酐)水合反應(yīng)的迅速進(jìn)行,反應(yīng)式為:CO2+H2O ?+H+[6]。由圖3 可見,海水酸化處理下珊瑚藻的碳酸酐酶活性明顯下降。在pH 7.8 和pH 7.4 處理下,叉節(jié)藻的碳酸酐酶活性分別下降了 23.10%和40.43%,和擬中葉藻則下降36.3%和59.61%,經(jīng)統(tǒng)計(jì)分析均達(dá)到了顯著水平。
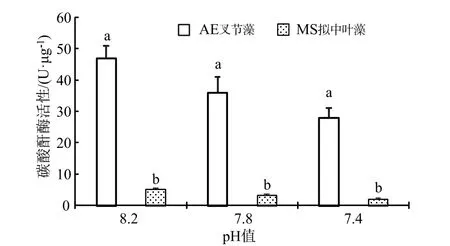
圖3 海水酸化處理對(duì)珊瑚藻碳酸酐酶活性的影響Fig.3 The effects of seawater acidification on the activities of CA of coralline algae
2.2 海水酸化對(duì)于珊瑚藻鈣化固碳速率及相對(duì)生長(zhǎng)速率的影響
珊瑚藻能夠通過鈣化作用(Calcification),將海水中的碳酸氫根轉(zhuǎn)化為氣態(tài)CO2,具體方程式為:海水酸化處理明顯降低了珊瑚藻鈣化固碳速率,在海水pH值7.4 的酸性環(huán)境條件下,叉節(jié)藻經(jīng)過90 d 處理后鈣化能力表現(xiàn)為負(fù)值[(-0.52±0.07)mg·g-1·h-1],較對(duì)照絕對(duì)值下降了110.42%,殼狀珊瑚藻也在酸化環(huán)境下明顯降低了鈣化能力,在海水pH 7.8 酸化處理的75 d,擬中葉藻的鈣化固碳速率就下降為了負(fù)值[(-0.08±0.01)mg·g-1·h-1] (圖4)。pH 7.8 和pH 7.4 的海水處理90 d 后,擬中葉藻的鈣化固碳速率絕對(duì)值分別下降了77.78%和128.89%。
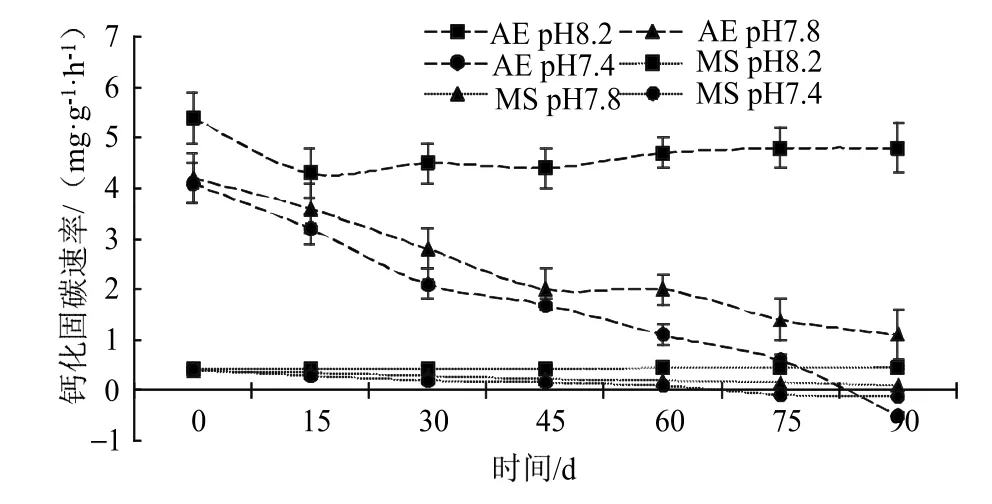
圖4 海水酸化處理對(duì)珊瑚藻鈣化固碳速率的影響Fig.4 The effects of seawater acidification on the rate of calcified carbon fixation of coralline algae
藻體鈣化量(PIC)與有機(jī)物(POC)的比例同時(shí)受到珊瑚藻光合固碳能力與鈣化固碳能力影響。由于海水酸化條件下,珊瑚藻的光合能力明顯提升,鈣化能力卻顯著下降,因此PIC/POC 比值有明顯的降低,pH 7.8 和pH 7.4 酸化海水處理下,叉節(jié)藻和擬中葉藻的PIC/POC 比值均在0.2 以下(圖5)。
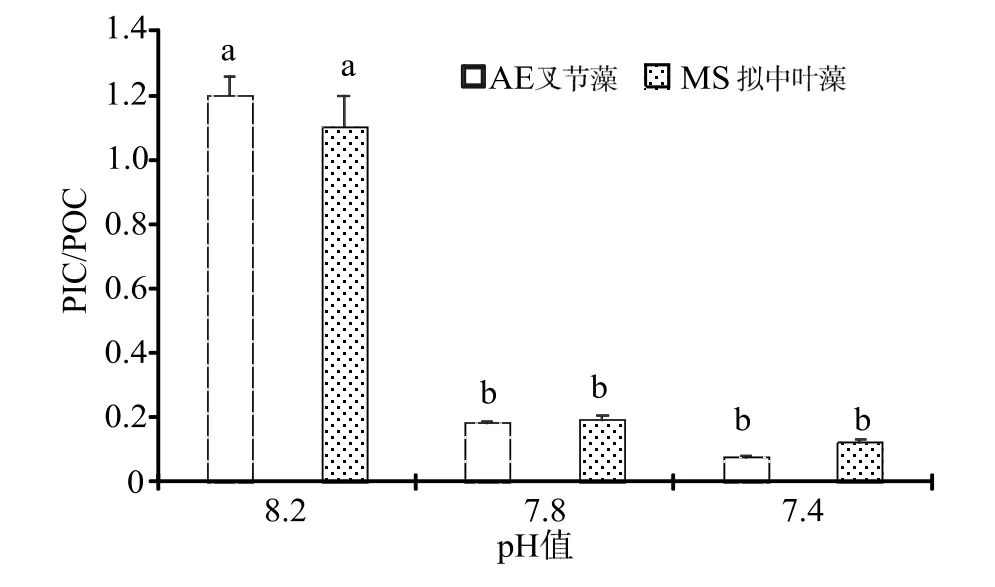
圖5 海水酸化處理對(duì)珊瑚藻PIC/POC 比值的影響Fig.5 The effects of seawater acidification on the ratio of PIC/POC of coralline algae
隨著海水酸化處理的pH 值逐漸降低,盡管光合固碳能力有所升高,但枝狀珊瑚藻的鈣化能力卻急劇下降,因此相對(duì)增長(zhǎng)率(RGR)越來越小,在pH7.4 海水處理下,叉節(jié)藻相對(duì)增長(zhǎng)率較對(duì)照明顯下降了38.24%(P< 0.05),而殼狀的擬中葉藻可能由于生長(zhǎng)緩慢,在90 d 的培養(yǎng)期內(nèi)相對(duì)增長(zhǎng)率的下降程度并未達(dá)到顯著水平(圖6)。
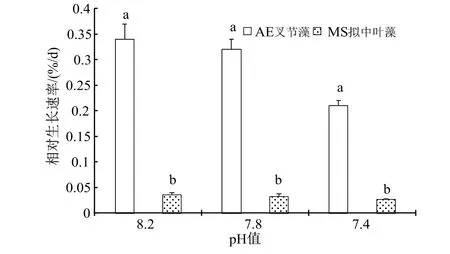
圖6 海水酸化處理對(duì)珊瑚藻生長(zhǎng)的影響Fig.6 The effects of seawater acidification on the growth of coralline algae
3 討論
海洋是地球碳循環(huán)體系里的核心部分,隨著全球變化的進(jìn)程,海水里的CO2體積分?jǐn)?shù)逐漸增大,加劇海洋酸化。海水pH 值不斷降低,極大地改變了海洋碳酸鹽系統(tǒng)面貌,對(duì)于在全球碳循環(huán)中扮演舉足輕重角色的海洋鈣化生物,包括鈣化海藻也將產(chǎn)生重要影響[8]。作為全球海洋底棲世界的最重要成分,珊瑚藻的生理生態(tài)特性在國內(nèi)外已有一些科學(xué)探索,包括光合生理、碳酸鹽骨架礦物學(xué)以及生態(tài)服務(wù)功能等,但仍有大量理論空白等待研究,人類尤其在殼狀珊瑚藻領(lǐng)域了解不多[9]。國內(nèi)外對(duì)于環(huán)境因素對(duì)珊瑚藻的生理影響研究很少,當(dāng)前僅少量報(bào)道研究了光照、海水酸化等因素對(duì)枝狀珊瑚藻的生理影響,至于殼狀珊瑚藻生理生化分析方面的專門研究則更為少見[3,10]。
深圳海域位于我國南亞熱帶海區(qū),尤其東部的大鵬海域水溫、鹽度比較穩(wěn)定,而且水體清澈、透光性好,適宜珊瑚藻棲息生長(zhǎng)[11]。本區(qū)珊瑚藻群落可以分為近岸與離岸兩種類型,近岸型的種類以對(duì)于濁度、沉積速率和懸浮物耐受能力較強(qiáng)的種類為主,并且對(duì)光照強(qiáng)度較敏感。本實(shí)驗(yàn)以本地區(qū)近岸型的典型枝狀珊瑚藻種類叉節(jié)藻和殼狀珊瑚藻種類擬中葉藻為研究對(duì)象,開展海水酸化相關(guān)研究。
3.1 海水酸化對(duì)珊瑚藻的光合作用影響分析
在以往的大型海藻研究中,都已發(fā)現(xiàn)高濃度CO2能促進(jìn)光合作用[12]。本實(shí)驗(yàn)通過向培養(yǎng)水體中充入高濃度的CO2,使pH 值降低(如降至pH=6.8),引起培養(yǎng)海水中CO2和的濃度增加,為光合作用的暗反應(yīng)過程提供了更充足的底物,同樣導(dǎo)致了小珊瑚藻光合作用的升高。其中,pH7.4 條件下藻體的光合固碳效率要高于pH 7.8。殼狀珊瑚藻在pH 值降低狀況下的光合能力上升變化幅度更為明顯(P< 0.01)。
在光合色素方面,實(shí)驗(yàn)發(fā)現(xiàn)高濃度CO2使得珊瑚藻體中葉綠素a 和藻紅蛋白的含量下降,類胡蘿卜素含量升高,這與之前在細(xì)基江蘺(Gracilaria tenuistipitata)等藻類研究結(jié)果相似[13]。保護(hù)色素含類胡蘿卜素量的增加,應(yīng)該是海水酸化刺激下藻體應(yīng)對(duì)的保護(hù)措施;藻紅蛋白起著吸收光能與傳遞能量的重要作用,從實(shí)驗(yàn)結(jié)果看,海水酸化處理對(duì)珊瑚藻藻紅蛋白有明顯的破壞作用。
3.2 海水酸化對(duì)珊瑚藻的鈣化作用影響分析
珊瑚藻通過光合和鈣化兩種方式固定二氧化碳,因?yàn)闊o法直接利用空氣中的無機(jī)碳,珊瑚藻在漫長(zhǎng)的進(jìn)化歷程中形成了一套無機(jī)碳濃縮機(jī)制,亦被稱為“CO2/泵”[14]。碳酸酐酶(CA)在其中扮演了將氣態(tài)CO2分子轉(zhuǎn)換為可溶性的核心角色,再以特殊轉(zhuǎn)運(yùn)機(jī)制,最終為珊瑚藻的光合暗反應(yīng)提供無機(jī)碳底物。在各種大型海藻中,紅藻門的海藻對(duì)利用能力較弱,遠(yuǎn)不如褐藻和綠藻[15]。因此碳酸酐酶(CA)通過CO2濃縮機(jī)制提供足夠的濃度,對(duì)于保障珊瑚藻等紅藻類的光合能力有著重要意義。對(duì)于鈣化藻類的無柄珊瑚藻(Corallina sessilis)相關(guān)研究結(jié)果顯示,海水CO2濃度升高會(huì)導(dǎo)致碳酸酐酶活性下降[16],主要原因可能在于濃度的不斷提高導(dǎo)致了CA 的活性受到抑制,而本研究在枝狀珊瑚藻和殼狀珊瑚藻的實(shí)驗(yàn)結(jié)果也與此類似。
珊瑚藻的生命過程中涵括了鈣化作用和有機(jī)碳生產(chǎn)兩個(gè)過程,藻體通過有機(jī)碳泵(生物泵)從事有機(jī)碳的初級(jí)生產(chǎn),通過碳酸鹽反向泵(溶解泵)完成胞壁沉積CaCO3的鈣化作用,最終,全球廣泛分布的珊瑚藻通過這兩個(gè)剛好相反同時(shí)又相輔相成的化學(xué)過程實(shí)現(xiàn)對(duì)大氣CO2濃度的調(diào)節(jié),進(jìn)而影響全球的氣候變化[17]。珊瑚藻屬于對(duì)CO2濃度升高引起海水酸化過程高度敏感的重點(diǎn)生物種之一,其鈣化作用容易受到空氣和海水中 CO2濃度變化的雙重影響。學(xué)術(shù)界普遍的認(rèn)識(shí)是,當(dāng)空氣CO2體積分?jǐn)?shù)升高時(shí),海水中H+濃度對(duì)數(shù)值將相應(yīng)減小,導(dǎo)致CaCO3飽和度逐漸降低,珊瑚藻鈣化因此會(huì)受到抑制[18]。徐智廣等[19]研究發(fā)現(xiàn)海水pH 值5.5 的酸性環(huán)境條件下,小珊瑚藻鈣化表現(xiàn)為負(fù)值[(-2.53±0.57)mg·g-1·h-1],這可能是過多的H+使得小珊瑚藻的鈣化組織部分溶解所造成。本研究同樣發(fā)現(xiàn)海水酸化處理明顯降低珊瑚藻鈣化固碳速率,海水pH 值7.4 的酸性環(huán)境條件下,叉節(jié)藻和擬中葉藻在實(shí)驗(yàn)處理后鈣化均表現(xiàn)為負(fù)值。在海水pH 7.8 酸化處理的75 d,擬中葉藻的鈣化固碳速率就已下降為負(fù)值[(-0.08±0.01)mg·g-1·h-1] 。海水酸化條件下,珊瑚藻的光合能力明顯提升,鈣化能力卻顯著下降,因此PIC/POC 比值相應(yīng)有明顯降低。pH 7.8 和pH 7.4 酸化海水處理下,叉節(jié)藻和擬中葉藻的PIC/POC 比值均在0.2以下。
3.3 海水酸化對(duì)珊瑚藻的生長(zhǎng)影響分析
已有的相關(guān)研究顯示[16,20],珊瑚藻的生長(zhǎng)及生理活性對(duì)于海水酸化以及伴隨而來的海水溫度升高有負(fù)面的反應(yīng),海水CO2濃度升高可抑制珊瑚藻的鈣化與生長(zhǎng),破壞藻體的生理結(jié)構(gòu)。在本實(shí)驗(yàn)中,枝狀珊瑚藻隨著海水酸化處理的pH 值下降出,光合固碳能力有所升高,但鈣化能力卻急劇下降,因此相對(duì)增長(zhǎng)率(RGR)越來越小,在pH 7.4 海水處理下,叉節(jié)藻相對(duì)增長(zhǎng)率較對(duì)照明顯下降了38.24%(P< 0.05),而殼狀的擬中葉藻由于生長(zhǎng)緩慢的原因,在90 d 培養(yǎng)期內(nèi)相對(duì)增長(zhǎng)率的下降程度并未達(dá)到顯著水平。
海水酸化對(duì)于殼狀及枝狀珊瑚藻的生長(zhǎng)、光合色素積累、碳酸酐酶活性以及鈣化固碳能力的影響存在明顯差異。殼狀珊瑚藻在海水酸化下的鈣化生理指標(biāo)和生長(zhǎng)數(shù)值變化幅度不算明顯,但相對(duì)差異更為顯著。實(shí)驗(yàn)中殼狀珊瑚藻擬中葉藻可能由于本身生長(zhǎng)緩慢,且由于本實(shí)驗(yàn)設(shè)計(jì)周期的持續(xù)時(shí)間有限,暫未發(fā)現(xiàn)海水酸化對(duì)于藻體生長(zhǎng)的負(fù)面影響達(dá)到顯著水平,尚有待今后更長(zhǎng)周期的培養(yǎng)和實(shí)驗(yàn)。
綜上,一定pH 值范圍內(nèi)的海水酸化處理,對(duì)于珊瑚藻的光合作用有所促進(jìn),同時(shí)明顯降低鈣化量和碳酸酐酶活性,改變光合色素含量與PIC/POC比值,進(jìn)而抑制枝狀藻體生長(zhǎng)。鑒于珊瑚藻在海洋生態(tài)系統(tǒng)中所承擔(dān)的重要角色,在未來研究中,非常值得對(duì)珊瑚藻,尤其是殼狀珊瑚藻的鈣化過程開展更加廣泛和深入的研究,為海洋生物對(duì)于海水酸化響應(yīng)研究,以及全球變化下生物圈碳循環(huán)與碳匯的研究提供相應(yīng)依據(jù)[2-3,21]。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(shù)(2020年3期)2020-12-06 07:32:54
現(xiàn)代裝飾(2020年11期)2020-11-27 01:47:48
中學(xué)生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(jí)(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時(shí)刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
