外源CO對采后棗果實鏈格孢菌浸染過程中蛋白組的影響
2021-06-19 03:18:28于有偉郭毅晶張靜茹張少穎
食品工業科技 2021年2期
于有偉,郭毅晶,康 琳,王 瓊,張靜茹,張少穎
(山西師范大學食品科學學院,山西臨汾 041004)
棗(ZiziphusjujubaMill.)為鼠李科棗屬植物,在我國已經有4000年的栽培歷史,棗果實滋味甘甜,口感清脆,含有各種豐富的糖、礦物質、氨基酸、維生素等營養物質,特別是維生素C的含量很高,而且還含有豐富的黃酮、多酚等功能成分,具有養胃、健脾、補血、保肝等醫療保健作用,是一種藥食同源的果品[1]。但是鮮棗不耐貯藏,采后極易感染鏈格孢屬(Alternaria)、鐮刀菌屬(Fusarium)、青霉屬(Penicillium)、毛霉屬(Mucor)等微生物,其中,由鏈格孢菌(A.alternata)引發的黑斑病是采后棗果實的主要病害,引起果實腐爛變質,影響貨架期,降低了鮮棗的商品價值[2-3],因此,研究棗果實的采后病害防控有重要意義。
近年來,CO在園藝產品采后生理方面的研究表明,通過CO熏蒸可以抑制鮮切蓮藕的褐變[4],延緩鮮切月季的衰老[5],保持桃、冬棗等果實的采后品質,延緩果實的采后衰老,延長其貨架期[6-8]。高濃度的CO還可以作為殺菌劑,抑制蘋果貯藏期間的Botrytiscinerea,Moniliniafructicola和Penicilliumexpansum的生長[9]。王東[10]的研究也表明,CO可能參與了HO-1調控的煙草對煙草花葉病毒的抵御。也有研究表明,低濃度的CO處理在抑制冬棗采后果實的腐爛[7,11]上有積極作用,主要是由于CO處理能通過提高果實抗病相關物質(多酚、黃酮、木質素)的含量和抗病相關蛋白(幾丁質酶、β-1,3葡聚糖酶、苯丙氨酸解氨酶、多酚氧化酶)的活性增強棗果實對采后病害的抵御能力。目前,CO在果蔬采后貯藏保鮮方面的研究還不多,CO在果實采后抗病方面的研究還僅停留在其抗病作用的初步探究階段,其具體的抗病作用機制還有待于進一步的研究。
從蛋白水平入手,利用蛋白質組學的方法探索植物與病原菌相互作用的分子機制,是植物抗病研究的重要手段。目前,對于采后受鏈格孢菌侵染而患黑斑病的棗果實在蛋白質表達差異方面的研究相對較少,而對外源CO在增強采后棗果實抵抗鏈格孢菌浸染過程中蛋白差異表達調控的研究還未見報道。因此,為了進一步了解CO處理對棗果實鏈格孢菌侵染過程中果肉蛋白質差異表達的影響,本研究用CO處理棗果實再對處理后的棗果實接種病原菌,研究外源CO處理后采后棗果實響應黑斑病過程中的蛋白表達及差異蛋白質的功能和調控作用,為進一步解析CO在誘導棗果實采后抗病性中的作用機制奠定基礎。
1 材料與方法
1.1 材料與儀器
供試冬棗(沾化冬棗) 采自臨汾市堯鄉冬棗種植基地,白熟期采收,采摘后迅速運回實驗室,選擇大小一致、無病蟲害和機械損傷的果實作為試驗材料;鏈格孢菌(BNCC115062) 購自北納生物有限公司;CO氣體(純度99.99%) 購自北京華能特種氣體有限公司。
AKTA Purifier 100全自動蛋白層析系統 美國通用電氣醫療集團;759S紫外可見分光光度計 上海棱光技術有限公司;MDF-U53V超低溫冰箱 日本三洋集團;H1850R臺式高速冷凍離心機 湖南湘儀實驗室儀器;EASY-nLCTM1200色譜系統和Q-Exactive質譜儀 美國Thermo Scientific公司。
1.2 實驗方法
1.2.1 鏈格孢菌孢子懸浮液的配制 參照He的方法[12],將鏈格孢菌接種于PDA平板上,28 ℃下培養10 d。用含有0.05% Tween-20的無菌水將孢子從平板上沖洗取出,三層紗布過濾后,顯微鏡計數,將孢子懸浮液的濃度調整為1×105個/mL。
1.2.2 試驗處理及取樣 選取60個大小一致、無病蟲害和機械損傷的棗果實,用2%的次氯酸鈉溶液清洗晾干后,再平均分成2組,每組30個果實。根據前期預實驗的CO濃度篩選結果,10 μmol/L CO熏蒸2 h的處理效果最佳,因此,本實驗分別進行如下處理:CO+A.alternata處理(記為COA),將果實置于密閉干燥器中用10 μmol/L CO熏蒸2 h,取出后在室溫下放置24 h,然后在棗果實的中部接種A.alternate的孢子溶液5μL;A.alternata接種處理(記為A),用打孔器在在赤道處打深 2 mm,直徑為3 mm的小孔,再用移液槍向小孔中注入5 μL孢子溶液進行接種。處理后兩組果實均置于帶孔的塑料盒中于室溫下(20±2 ℃,相對濕度75%~85%)貯藏。分別于處理后0、24、48和96 h后,削去棗的表皮后,切取距接種點5 mm的棗果肉10 g用于蛋白組的分析。
1.2.3 蛋白組分析
1.2.3.1 蛋白質提取和肽段酶解 樣品采用SDT(100 mmol/L Tris/HCl pH7.6,0.1 mol/L DTT,4%(w/v)SDS)裂解液提取蛋白質[13],用BCA(bicinchoninic acid)法定量后,再用FASP(Filter aided proteome preparation)法進行胰蛋白酶酶解,用C18 Cartridge填料的固相萃取小柱對酶解的肽段脫鹽,肽段經凍干、復溶后,測定OD280定量。
1.2.3.2 肽段標記 分別取100 μg多肽樣本,按照試劑盒說明(ITRAQ-8plex)標記各個樣本。
1.2.3.3 SCX色譜分級 混合標記后的肽段,分級使用全自動蛋白層析系統(AKTA Purifier 100)。色譜柱的平衡液(pH3.0)為25% ACN,10 mmol/L KH2PO4;色譜柱的洗脫液(pH3.0)為500 mmol/L KCl,10 mmol/L KH2PO4,25% CAN。色譜柱平衡后,上樣分離,流速1 mL/min。液相梯度如下:0~25 min,洗脫液的線性梯度為0%~10%;25~32 min,為10%~20%;32~42 min,為20%~45%;42~47 min,為45%~100%;47~60 min維持在100%;60 min后,洗脫液重置為0%;邊洗脫邊檢測OD214,洗脫組分的分收集為每2 min/次,各組分凍干后用C18 Cartridge脫鹽[13]。
1.2.3.4 LC-MS/MS數據采集 分級樣品的分離用HPLC系統Easy nLC(納升流速)進行[13]。緩沖液:A液為0.1%甲酸水溶液,B液為0.1%甲酸乙腈水溶液。用95% 的A液平衡色譜柱。上樣柱為Thermo scientific EASY column(5 μm×100 μm×2 cm,C18),上樣后,再經Thermo scientific EASY column分析柱(3 μm×75 μm×10 cm,C18)分離,流速為250 nL/min。液相梯度如下:0~50 min,B液線性梯度從0%~35%;50~58 min,從35%~100%;58~60 min,維持在100%。
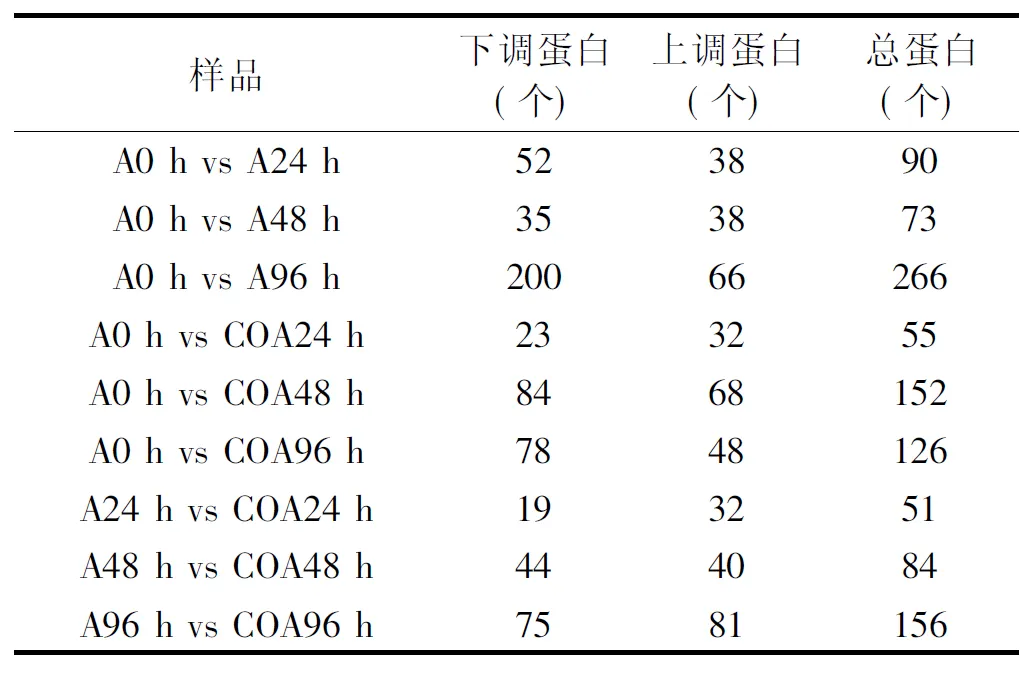
表1 不同時期和不同處理的差異蛋白個數Table 1 Number of differential expressed proteins in different periods and different treatments
質譜分析Q-Exactive質譜儀進行[13],正離子檢測,分析時長為60 min,母離子掃描范圍300~1800 m/z,一級質譜分辨率為70000 at m/z 200,AGC(Automatic gain control)target為3e6,Maximum IT為10 ms,動態排除時間為40.0 s。按照下列方法采集多肽和多肽的碎片的質荷比:每次全掃描后采集10個碎片圖譜(MS2 scan),MS2 Activation Type為HCD,Isolation window為2 m/z,二級質譜分辨率為17500 at 200 m/z,Normalized collision energy為30 eV,Underfill ratio為0.1%。
1.2.3.5 蛋白質鑒定和定量分析 用Proteome Discoverer 1.4檢索質譜測試原始數據的相應數據庫,再進行蛋白質鑒定及定量分析,以差異倍數值大于1.5倍為顯著上調、小于0.667為顯著下調為標準。
1.2.3.6 生物信息學分析 GO功能注釋利用軟件Omicsbean(http://www.omicsbean.cn/)對目標蛋白質集合進行GO注釋。KEGG通路注釋利用軟件Omicsbean對目標蛋白質集合進行KEGG通路注釋。
1.3 數據分析
數據的方差分析和繪圖選用DPS v14.10和Microsoft Excel 2010進行,差異顯著性檢驗(P<0.05)用t測驗。
2 結果與分析
2.1 蛋白質鑒定結果和差異蛋白分析
由表1可以看出,所有樣本共鑒定出的肽段數為5730,蛋白為2158;其中差異蛋白總數為1053,上調的差異蛋白數為443,下調的差異蛋白數為610;與0 h相比,接種處理的冬棗果實在24、48和96 h都有不同數量的蛋白質的差異表達蛋白,其中,處理后24、48 h時,上調差異蛋白數分別為38和38,下調差異蛋白數分別為52和35,而96 h后上調和下調的差異蛋白數量分別上升為66和200;在24、48和96 h時,CO+接種處理果實中的蛋白與0 h的相比也出現了不同數量的差異表達,但是從總量上看,除48 h的樣品外,24和96 h的差異蛋白數量均低于接種處理的;而相對于接種,CO+接種處理在24、48和96 h時樣品的上調差異表達蛋白質數分別為32、40、81,下調差異表達蛋白質數分別為19、44和75。
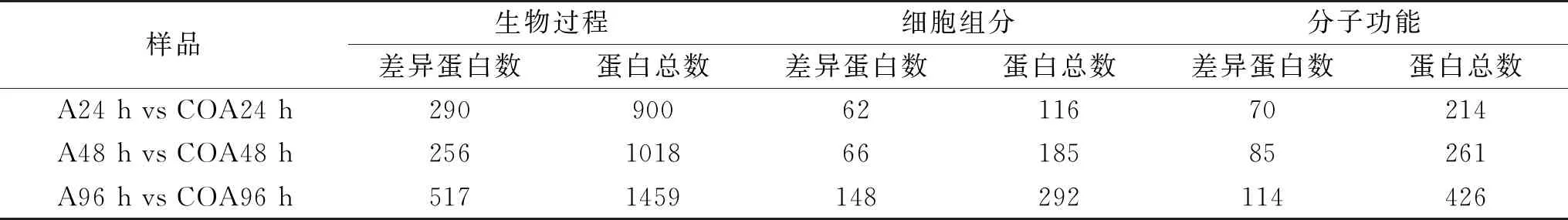
表2 差異蛋白富集統計分析Table 2 Statistical analysis of differential protein enrichment
2.2 差異蛋白的GO分析
如表2所示,相對于單獨接種處理,CO+接種處理后隨著貯藏時間的延長,在細胞組分、分子功能和生物過程三種類別上富集的總差異蛋白數增大。生物過程上,在48 h富集的差異蛋白達到顯著水平的有256個,低于24 h和96 h的蛋白數290和517,而在細胞組分和分子功能上,三個時間點上達到顯著水平的差異蛋白數均呈現上升趨勢。
為了進一步了解差異蛋白質的GO富集情況,分別選取在細胞組分、分子功能和生物過程三個類別上24、48和96 h 時間點的分析顯著性排名前十的條目進行分析(見圖1)。細胞組分上(圖1A),在24、48和96 h時,兩個處理3個時期共有18個類別的顯著性富集,而且隨貯藏時間延長,各組分的富集量增加且差異更顯著,差異蛋白的富集在細胞質膜和細胞器的積累增加,細胞液和細胞質的富集程度在三個時期均較高。分子功能上(圖1B),三個時間段上共有14個類別的顯著性富集,共有的富集包括脫落酸結合、蛋白結合、類異戊二烯結合、激素結合、乙醇結合、蛋白磷酸酶抑制劑活性和磷酸酶抑制劑活性和單羧酸結合。處理后48 h,差異蛋白的分子功能富集最多。生物過程上(圖1C),48和96 h時,有3個顯著共同類別富集,包括過單一生物代謝過程、單一生物細胞過程和單一生物過程,而96 h時的差異蛋白生物過程的富集量更大。
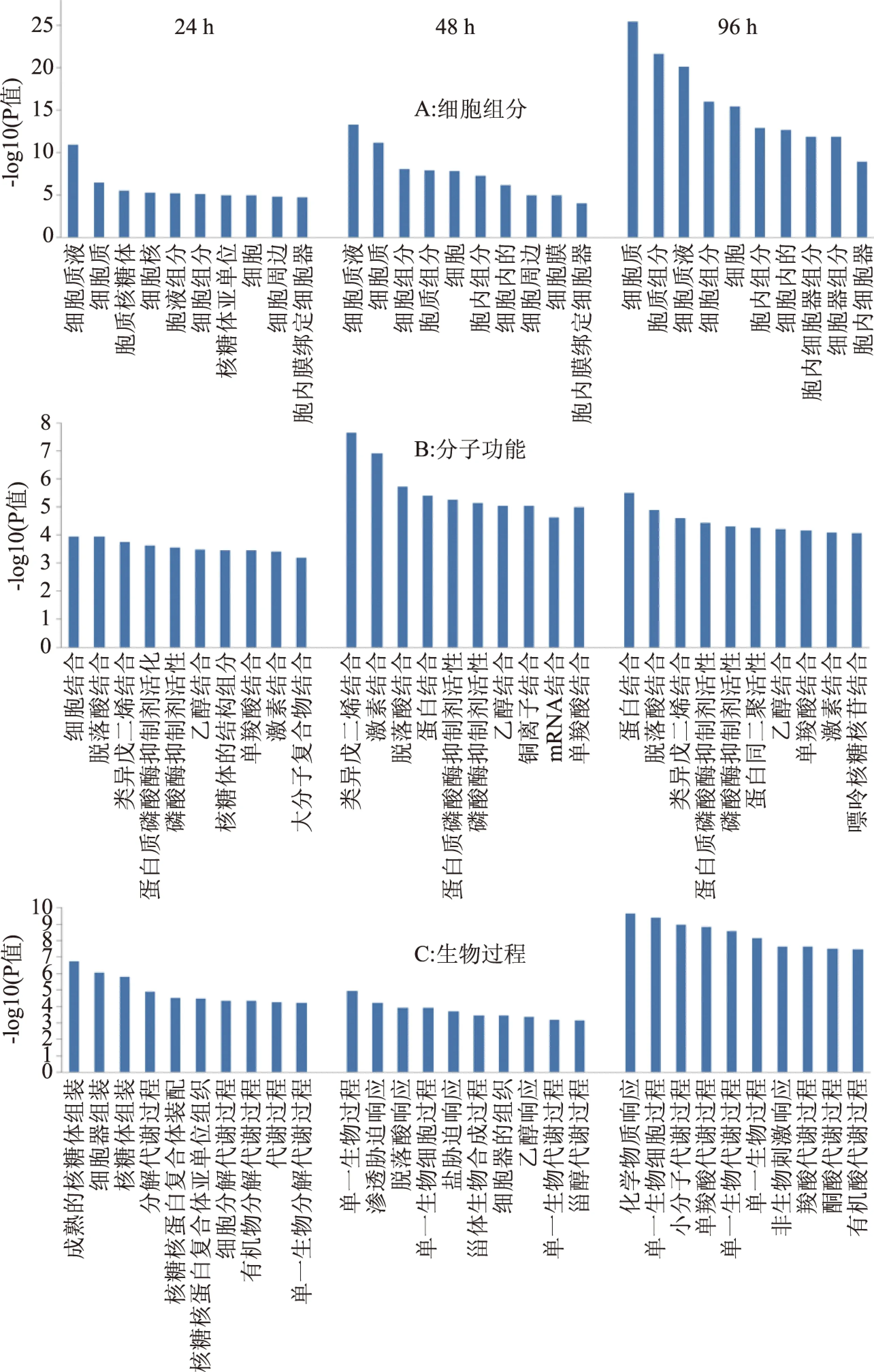
圖1 差異蛋白GO富集分析Fig.1 GO enrichment analysis of differential proteins 注:細胞組分(A)、分子功能(B)和細胞過程(C)。
2.3 差異表達蛋白質 KEGG 通路富集分析
運用KEGG數據庫對差異蛋白進行代謝途徑富集分析。處理后24 h,冬棗果實的差異表達蛋白主要富集在5個代謝途徑(圖2),其中核糖體、甾體生物合成相互作用途徑富集度較高,其后為植物-病原菌、mRNA監視途徑、果糖和甘露糖代謝通路;處理后48和96 h差異表達蛋白富集的主要代謝途徑明顯增多,均為9個。48 h差異蛋白主要富集在鞘糖脂生物合成、植物激素信號轉導、氨酰-tRNA生物合成、磷脂酰肌醇信號系統、光合組織中的碳固定、維生素B6和生物素代謝、賴氨酸生物合成和碳代謝通路;96 h差異蛋白富集在甾體生物合成、糖酵解/糖異生、碳代謝、細胞內吞作用、氨基酸生物合成、單萜生物合成、內質網中的蛋白加工、錯配修復和亞油酸代謝通路。差異蛋白在48和96 h共同富集的途徑為碳代謝,而甾體生物合成途徑在24和96 h都有富集。
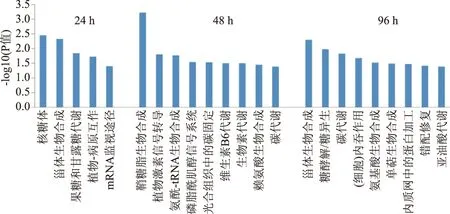
圖2 差異表達蛋白的KEGG 通路富集分析Fig.2 KEGG pathway enrichment analysis of differential proteins
2.4 目標差異蛋白
根據差異蛋白GO富集和KEGG途徑富集的分析結果,發現經CO處理后,棗果實響應A.alternata過程中的差異蛋白主要富集在5個主要的功能過程,分別為:碳水化合物和能量代謝、氨基酸和脂類代謝、次生代謝產物的生物合成和防御相關蛋白、信號的感知和信號轉導、轉錄和轉錄后調控。共篩選出72個主要的目標差異蛋白(表3)。篩選出的差異蛋白主要包括:碳水化合物和能量代謝相關的蛋白16個;氨基酸和脂類代謝相關的蛋白12個;次生代謝產物的生物合成和防御相關蛋白19個;信號的感知和信號轉導的相關蛋白10個;轉錄和轉錄后調控的相關蛋白15個。

表3 差異蛋白功能分類Table 3 The functional classification of DEPs
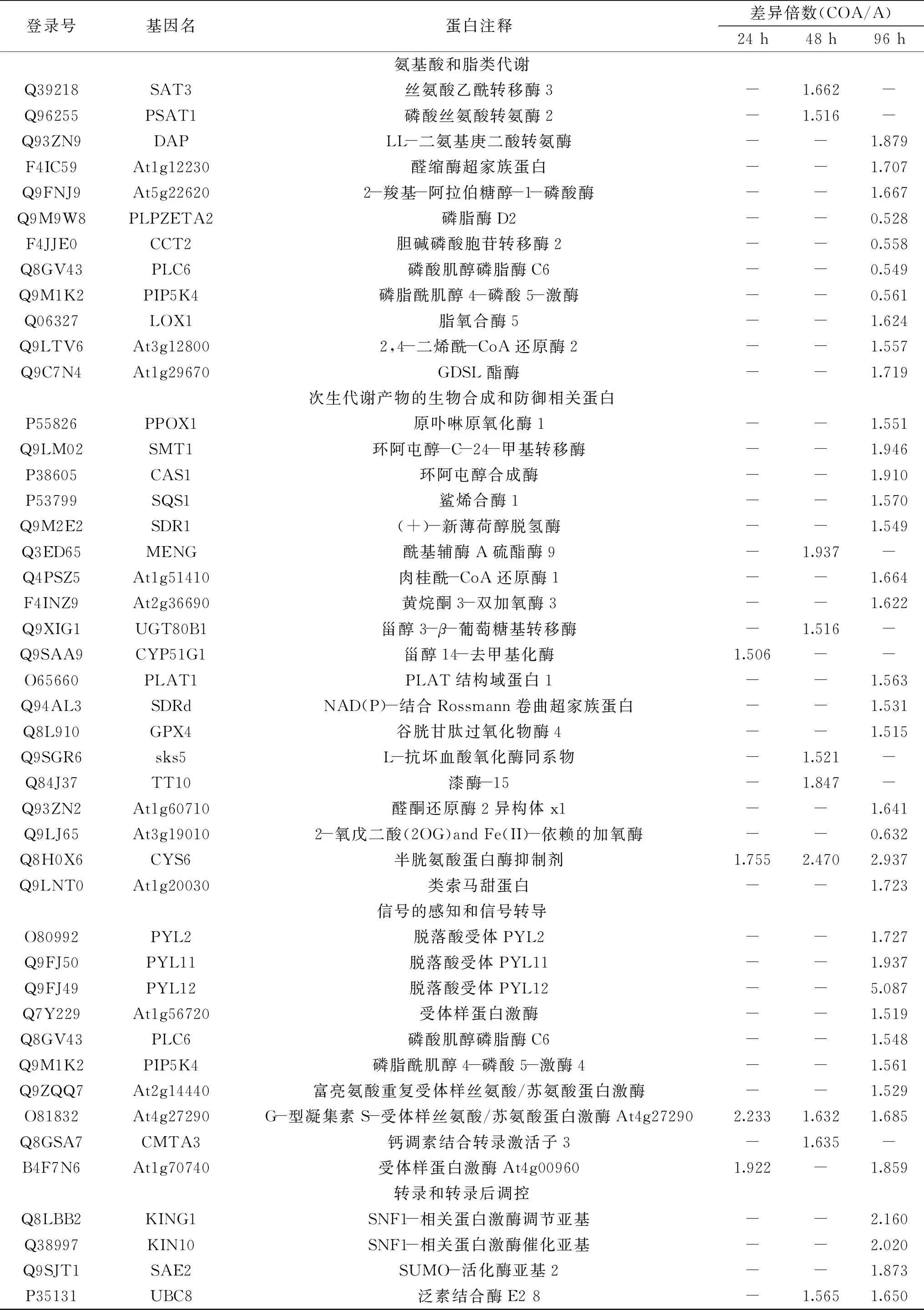
續表

續表
3 討論
3.1 碳水化合物和能量代謝相關蛋白
有證據表明,植物防御需要大量能量,而產能的初級代謝在植物抗性反應中起著重要作用[14-15]。植物抵御病原菌入侵也會導致能量流失,引發細胞內的碳水化合物代謝增強[16]。本研究篩選出16個與碳水化合物和能量代謝有關的差異蛋白。其中,在接種后96 h內,有5個上調的與能量代謝相關的差異蛋白(P93306、F4HXY8、O82628、Q9FT52和Q5M729),6個與糖酵解及三羧酸循環相關的上調差異蛋白(Q9SN86、Q9M039、Q9C9P3、Q9MB58、Q9C9C4和P25696)以及5個上調的糖苷酶蛋白(Q9SU50、Q8VZJ2、F4IHD3、Q9SCW1和Q9FT97)。Li等[17]的研究表明,外源NO處理可以提高糖酵解和三羧酸循環相關蛋白的表達豐度,從而為桃果實應答采后病原菌提供能量,本研究的結果證明,外源CO處理也有類似的效果。有研究表明,可溶性糖在果實對多種脅迫源的響應中起重要作用[18],海藻糖酶、β-1,3-葡萄糖苷酶和半乳糖苷酶作為糖苷水解酶可以將糖苷鍵水解生成小分子可溶性糖,三者均在植物抗病過程發揮作用[19-21]。本研究也發現,外源CO處理提高了棗果實響應鏈格孢菌侵染過程中細胞內的海藻糖酶、2個β-1,3-葡萄糖苷酶和2個半乳糖苷酶的表達量。
3.2 脂類和氨基酸代謝相關蛋白
植物體內的氨基酸是許多其它生物代謝產物的前體物質,而這些代謝產物涉及植物逆境響應等多種功能,是植物免疫系統必需組成部分[22-23]。文景芝等[24]的研究表明,大豆種子分泌物中游離氨基酸在一定程度上可能作為信號分子參與大豆疫霉寄主選擇性和寄主抗病性。此外,L-谷氨酸的代謝物GABA、Met、Lys和Arg,也能誘導番茄果實對灰霉病菌產生抗性[25]。本研究結果顯示,在棗果實和病原菌相互作用的96 h內,CO處理可以誘導氨基酸代謝相關的5個差異蛋白的上調,涉及His、Lys及Met的生物合成。
脂肪酸和脂類是植物細胞的必需成分,不僅為各種代謝過程提供結構完整性和能量,還可作為信號轉導介質[26]。脂類代謝及其代謝產物也參與植物對病原菌脅迫的響應,包括氧化脂類,與脂氧合酶途徑(如與JA的合成)有關;不飽和脂肪酸途徑,與膜脂類組成和防御信號的感知和轉導有關;以及長鏈脂肪酸途徑[27-29]。CO處理后,發現了7個與脂類代謝相關的差異蛋白,其中Q8GV43和Q9M1K2參與磷脂酰肌醇信號調控和磷酸肌醇代謝;Q9M9W8和F4JJE0參與甘油磷脂代謝;Q9LTV6和Q06327參與脂類氧化;Q9C7N4涉及脂肪的水解。有報道表明,Q9LTV6和Q06327參與了植物抗病誘導過程中的JA、SA、Eth和ABA信號響應和調控[30-31]。因此推測,CO可能通過改變棗果實的脂類代謝參與脂類物質介導信號轉導,從而抵御病原菌的入侵。
3.3 次生代謝產物的合成和防御相關蛋白
植物可以合成多種次生代謝產物來對抗生物和非生物的脅迫[32-33]。多酚、黃酮、木質素,甾醇及其它萜類物質等次生代謝產物都能幫助植物抵御病原菌的入侵[34]。本研究的結果顯示,外源CO處理后,1個卟啉代謝相關蛋白(P55826),9個甾醇、木質素、黃酮醇及其它萜類合成相關蛋白(Q9LM02、P38605、P53799、Q9M2E2、Q3ED65、Q4PSZ5、F4INZ9、Q9XIG1、Q9SAA9)上調,因此,CO可能通過上調次生代謝產物的合成增強棗果實抵御病原菌的入侵的能力。研究發現,病原菌入侵時,果實組織會在短期內產生大量活性氧(ROS),阻止病原菌的侵害[35]。低濃度的ROS也可作為信號分子,誘導防御基因的表達。而當植物細胞內的ROS積累過多時,會導致細胞膜脂過氧化,破壞寄主的防御系統。植物體內的ROS清除系統在保持ROS的動態平衡中起關鍵作用[36]。本研究在CO處理組的棗果實中發現了7個與ROS平衡或氧還平衡相關的差異蛋白(O65660、Q94AL3、Q8L910、Q9SGR6、Q84J37、Q93ZN2、Q9LJ65);還發現2個上調的抗病相關的差異蛋白Q8H0X6和Q9LNT0。由此可見,外源CO在維持ROS的代謝平衡和提高抗病相關蛋白的表達量上發揮積極作用。
3.4 信號感知和信號轉導
在植物對抗病原菌入侵的初期,植物細胞與病原菌的相互識別是關鍵[37]。植物通過細胞表面的受體分子與病原菌激發子相結合,啟動多種防御機制,使細胞對病原菌產生抗性[38]。本研究發現,外源CO處理誘導了Q9ZQQ7、O81832、Q7Y229和B4F7N6的表達。已有研究證實,以上4種蛋白均參與了植物細胞對病原菌激發子的識別,介導下游防衛反應的發生[39-41]。
在植物脅迫應答過程中,CO信號與其他脅迫響應信號網絡重疊,且可能通過與這些信號分子協同作用調控脅迫反應[42]。本研究分析了CO誘導棗果實抵抗病原菌侵染過程中涉及的信號轉導的差異蛋白,根據這些差異蛋白在信號轉導途徑中的作用把它們分為與Ca2+信號、磷脂酰肌醇信號、ABA信號及JA信號相關的四個類別。Ca2+作為第二信使,參與了植物細胞與病原菌互作的信號轉導過程,而磷脂酰肌醇信號又參與Ca2+信號的調節。本研究發現外源CO處理后,1個Ca2+信號相關的差異蛋白(Q8GSA7)和2個磷脂酰肌醇信號相關差異蛋白(Q8GV43和Q9M1K2)上調,這表明了CO可以通過Ca2+途徑調控棗果實對病原菌的防御反應。研究還發現了3個與ABA信號轉導相關的蛋白(O80992、Q9FJ50和Q9FJ49)和1個與JA生物合成相關的蛋白(Q06327)。有研究表明,外源ABA可以誘導植物對病原菌的抗性[43-44],而且ABA的受體蛋白O80992可能是一種PR蛋白,參與植物的抗病過程[45]。基于以上分析,我們認為Ca2+、磷脂酰肌醇信號、ABA及JA信號可能參與了CO對棗果實抗病性的誘導。
3.5 轉錄和轉錄后調節
基因轉錄和轉錄后的調節在植物細胞抵抗病原菌入侵的過程中也發揮著至關重要的作用[17]。本研究發現,CO處理改變了一些與蛋白質合成、加工修飾和降解相關的蛋白表達,包括7個核糖體蛋白、2個SNF1-相關蛋白激酶、3個介導泛素化的蛋白和3個參與蛋白加工的熱激蛋白。雖然,有關核糖體參與植物與病原菌互作的報道很少,但有研究表明,核糖體蛋白L12參與了煙草和擬南芥對病原菌脅迫的響應[46],而核糖體蛋白S30也參與了花生對青枯病防御[47]。由此可以推測,CO處理后,核糖體差異蛋白可能參與棗果實對病原菌的防御應答。SNF1-相關蛋白激酶,是一類Ser/Thr蛋白激酶,參與植物的抗病信號調控[48]。本研究中檢測到的2個SNF1-相關蛋白激酶均為上調表達,推測其可能在CO誘導的棗果實抗病過程中發揮積極作用。泛素化是植物體內一種廣泛存在的細胞反應調控機制,參與植物的抗病調控[49]。本研究中3個介導泛素化蛋白的上調表達也暗示蛋白質的泛素化修飾可能參與了CO誘導的棗果實對黑斑病的防御反應。在本研究中發現的3種熱激蛋白在外源CO處理下被鑒定出上調表達。熱激蛋白參與植物的抗病[50],Li等[17]也發現,在NO處理誘導的桃果實抗病過程中,果實中的熱激蛋白表達上調,這與本研究結果相似。
4 結論
本研究的蛋白組學分析結果顯示,CO在棗果實響應A.alternate的早期侵染過程中的作用包括:a.感知病原菌入侵的信號并使轉導激活,然后通過轉錄及轉錄后的調節來調控下游蛋白的表達;b.次生代謝產物的合成啟動并與抗病相關蛋白協同作用;c.碳水化合物、氨基酸和脂類代謝等基礎代謝途徑重構以響應A.alternate的脅迫。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
音樂探索(2022年2期)2022-05-30 21:01:37
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
航空世界(2020年10期)2020-01-19 14:36:20
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
