長雙歧桿菌特性及應用研究進展
2021-06-19 06:02:22關嘉琦岳瑩雪史佳鷺MDMasumuzzaman冷友斌霍貴成李柏良
食品工業科技 2021年12期
關嘉琦,邱 冀,岳瑩雪,史佳鷺,MD Masumuzzaman,冷友斌,李 春,霍貴成,,李柏良,
(1.東北農業大學食品學院,黑龍江哈爾濱 150030;2.黑龍江飛鶴乳業有限公司,北京 100015)
雙歧桿菌屬(Bifidobacterium)是一種革蘭氏陽性、不運動、細胞呈桿狀、一端有時呈分叉狀的嚴格厭氧的細菌。雙歧桿菌作為腸道有益微生物,在保持機體健康方面起重要作用[1]。已知的32 種雙歧桿菌中有12 種來源于人類。長雙歧桿菌(Bifidobacterium longum)是在嬰兒和成人的腸道微生物群中發現的最常見的雙歧桿菌物種,可在人體腸道中定植[2]。2012 年出版的《伯杰氏系統細菌學手冊》[3]中長雙歧桿菌含3 個亞種,即長雙歧桿菌長亞種(B.longumsubsp.longum)、長雙歧桿菌嬰兒亞種(B.longumsubsp.infantis)和長雙歧桿菌豬亞種[4]。美國食品藥品監督管理局(FDA)和歐洲食品安全局(EFSA)認定長雙歧桿菌具有安全性[5],我國法規也批準長雙歧桿菌可用于食品及保健食品[6]。長雙歧桿菌在人類健康中發揮著重要作用,具有防治便秘[7]、抑制腸道致病菌[8]、調節腸道平衡、降低膽固醇[9]、促進營養物質的消化吸收、延緩衰老[10]和增強機體免疫活性[11]等生理功能。
本文綜述了長雙歧桿菌亞種分型鑒定的方法、包括定植在內的生物學特性;從長雙歧桿菌緩解腦腸綜合征以及自身免疫綜合征等方面,綜述了長雙歧桿菌對宿主發揮的重要益生功能及其作用機制;最后對其應用及前景進行綜述,以期加深和完善關于長雙歧桿菌的認識。
1 長雙歧桿菌生物學特性
長雙歧桿菌是母乳喂養嬰兒腸道中最豐富的雙歧桿菌種類之一。人體腸道內的雙歧桿菌數量隨著年齡的增大而逐漸減少,種類也逐步發生變化,初期以嬰兒雙歧桿菌和短雙歧桿菌為主要優勢菌,后轉變為青春雙歧桿菌和長雙歧桿菌[6]。另外,雙歧桿菌的形態也存在菌種特異性,如B.longum亞種主要為勺型[12]。
1.1 長雙歧桿菌亞種的分型鑒定
雙歧桿菌具有高達87.7%~99.5%的種間相似度,因此,對雙歧桿菌的分型鑒定提出了更高要求[13]。目前,在雙歧桿菌親緣物種鑒定方面,傳統的生理生化和16S rDNA 同源性分析,對親緣關系比較近的物種分辨率不高,故多使用全基因組測序及系統發育樹[14?15]、PCR-RFLP[16]、PCR-DGGE[17]、多位點測序分型(MLST)[18]、擴增片段長度多態性分析(AFLP)[19]等方法來區分不同亞種。
長雙歧桿菌的種內基因組多樣性涉及與碳水化合物代謝、細胞膜合成、移動元素相關的基因,這反映了三個亞種(長亞種、嬰兒亞種和豬亞種)的不同生態特異性[20]。越來越多的人以此作為區分長雙歧桿菌不同亞種的依據,而這需要找到能夠可靠區分不同亞種的特異性基因[21?22]。有文獻表明高度保守的熱應激蛋白60 基因廣泛分布于雙歧桿菌中,能彌補16S rDNA 同源性分析的不足[23],提高雙歧桿菌親緣物種鑒定的準確性。李明杰等人采用特異性基因擴增的方法,確認了長雙歧桿菌長亞種糖激酶基因和長雙歧桿菌嬰兒亞種唾液酸酶基因可實現長雙歧桿菌嬰兒亞種的快速區分[4]。
1.2 長雙歧桿菌定植特性
長雙歧桿菌在宿主腸道的定植是其發揮益生功能的基礎,而長雙歧桿菌在人體試驗中已被證明具有優越的腸道定植能力。長雙歧桿菌在兒童腸道中可定植超過5 年,即使在10 年后仍能檢測到該菌株[20]。人體腸道中長雙歧桿菌的定植和豐度因個體而異,這與分娩方式、喂養方式、年齡、地理和生理狀態有關[24?25]。雙歧桿菌等腸道微生物在人體消化道內分布情況如圖1 所示,可以發現,雙歧桿菌主要存在于結腸區域,而其他乳桿菌主要分布于小腸區域,這也是雙歧桿菌與其他乳酸菌的顯著差別。李讓等人借助PMA-qPCR 技術,發現B.longumBBMN68 在胃和小腸中存活率均較低,在結腸和直腸中存活率較高,并可在結腸中進行增殖[26]。長雙歧桿菌定植后會對人體腸道原住微生物群產生有益影響,例如通過增加青霉酸鹽、生物素和丁酸鹽水平與微生物群落相互作用,影響腸道代謝等[27]。
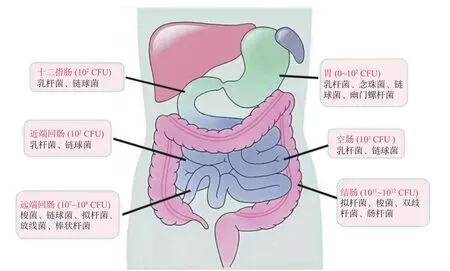
圖1 人消化道內的微生物分布[28]Fig.1 Distribution of microorganisms in human digestive tract[28]
1.2.1 糖代謝能力對長雙歧桿菌定植的影響 長雙歧桿菌的定植與碳源、膽鹽、酸、免疫反應、與其他微生物的競爭以及腸道粘膜清除有關。功能基因組分析揭示,長雙歧桿菌能夠在人類腸道中定植和存活,是因為長雙歧桿菌能抵抗宿主腸道環境的選擇性壓力,并具有多種與不同碳水化合物代謝相關的基因,其分解多糖的能力會影響其在哺乳動物腸道中的定植,例如在富含多糖(如淀粉,粘蛋白等)的環境中表現出物種特異性的適應性。碳源的競爭優勢增強了長雙歧桿菌定植并成為優勢菌的能力。
1.2.1.1 母乳低聚糖代謝對優先定植的影響 長雙歧桿菌可通過利用母乳低聚糖(HMO)和植物源性多糖快速增殖,最終成功定植于人體腸道[1],成為哺乳動物的第一批腸道共生菌。目前長雙歧桿菌已經進化出兩種同化HMO 的方式[29?30],一些長雙歧桿菌菌株利用胞外糖苷酶(例如α-l-巖藻糖苷酶和LnbX)將HMO 降解為細胞外的單糖和雙糖,包括唾液酸和巖藻糖等[31?34],再將降解的糖導入或與其他雙歧桿菌共享,即先定植的雙歧桿菌通過交叉喂養方式促進不能直接獲得乳源碳水化合物的雙歧桿菌完成定植。例如對B.bidifum2010、長雙歧桿菌嬰兒亞種B.longumCC15697、B.adolescentS22L 和B.breve12L 喂養的常規小鼠進行的體內實驗,基于轉錄組學并結合雙歧桿菌的單、雙或多個關聯的亞基因組分析進一步證實了哺乳動物腸道中不同雙歧桿菌之間存在交叉喂養[32],這從一定程度上解釋了人體發育初期腸道內雙歧桿菌種類及數量發生的變化。長雙歧桿菌的交叉喂養也被觀察到直接作用于其他腸道微生物,例如青春期雙歧桿菌和利用乳酸、產生丁酸的細菌(如Eubacterium hallii和Anaerostipes caccae)之間的代謝交叉喂養[35],因此長雙歧桿菌的優先定植,對人體腸道菌群的組成與演化具有重要意義。
而大多數長雙歧桿菌等可以使用轉運體直接內化HMO,然后在細胞內降解它們。對長雙歧桿菌嬰兒亞種B.longumATCC15697 和雙歧桿菌PRL2010的基因組分析揭示了這兩種雙歧桿菌對宿主的衍生聚糖(如HMO 和粘蛋白)的代謝機制。長雙歧桿菌嬰兒亞種B.longumATCC15697 的基因組包含編碼代謝HMO(如巖藻糖苷酶、唾液酸酶、B-己糖胺酶和B-半乳糖苷酶)基因位點,并內化HMO[36]。此外,該菌株的染色體還包含一個與尿素代謝有關的基因位點,尿素是母乳中重要的氮源[36]。決定母乳低聚糖巖藻糖基化模式的母體基因型在嬰兒斷奶前腸道微生物群的聚集中起著重要作用[37]。在這種情況下,特定的嬰兒相關雙歧桿菌,如長雙歧桿菌嬰兒亞種等可以有效代謝HMO 成分,如乳糖-N-四糖[36,38],因此在生命早期建立第一個腸道微生物群方面發揮關鍵作用。
1.2.1.2 特定碳源代謝與原住菌豐度對定植成功的影響 長雙歧桿菌的穩定定植還與腸道菌群中原住長雙歧桿菌豐度低、缺乏特定糖類利用基因有關。一項研究讓受試者單次口服長雙歧桿菌B.longumAH1206,六個月后發現該菌可在30%的受試者腸道內形成穩定的菌落[39]。盡管長雙歧桿菌B.longumAH1206 可以在一部分個體中存活,但移植成功率存在顯著差異。Li 等人一項基于亞基因組分析的表明,當同一物種的其他菌株已經存在,可一定程度說明具備該類物種生存的必需條件,此時新菌株定植成功率更高[40]。然而在長雙歧桿菌含量相對較高的個體中,由于存在系統發育相似性受限作用,原住長雙歧桿菌會競爭性地排除B.longumAH1206。
與常住長雙歧桿菌的總體豐度相比,其基因含量更能預測長雙歧桿菌B.longumAH1206 的定植成功率,這解釋了長雙歧桿菌B.longumAH1206可以在一些長雙歧桿菌含量較高的個體中持續存在的原因。Martiny 等鑒定了60 個亞基因組中的基因,其中與碳水化合物代謝和含半乳糖化合物的利用有關的基因豐度較低,認為與原住菌相比,利用特定碳水化合物的能力的差異是長雙歧桿菌定植的基礎。基于宏基因組分析的結果也進一步證實了Stecher等關于腸道菌群菌株豐度取決于某些限制性營養素的可用性的觀點[41]。
1.2.2 菌毛結構對長雙歧桿菌定植的影響 有報道顯示長雙歧桿菌的最初定植還與長雙歧桿菌的菌毛結構有關,菌毛結構是長雙歧桿菌粘附于腸道的關鍵。菌毛是一種表面相關蛋白,Foroni 等人在長雙歧桿菌B.longumATCC15697 中發現了類似菌毛結構的附屬物[42]。對長雙歧桿菌B.longumNCC2705 和長雙歧桿菌B.longumDJO10A 進行基因組分析,發現其含有由主要的菌毛亞單位編碼基因(fimA)、一個菌毛亞單位編碼基因(fimB)和一個sortase 酶基因(strA)組成的假定的菌毛基因簇。IVb 型菌毛也被證明在宿主的定植和持久性中起著關鍵作用[43]。Pilin 相關基因廣泛存在于長雙歧桿菌株。Chaplin的一項研究發現其所有的25 株長雙歧桿菌株(23 株長雙歧桿菌長亞種,2 株長雙歧桿菌嬰兒亞種)全部含有菌毛基因[20]。
2 長雙歧桿菌益生功能
一些研究表明長雙歧桿菌對宿主健康具有調節作用。例如,長雙歧桿菌可有效預防腦腸疾病和自身免疫性疾病等。這是因為長雙歧桿菌表面相關蛋白、胞外多糖或代謝產物與宿主存在相互作用,通過直接或間接途徑實現對宿主健康的調節作用。另外,相比于健康對照組,患有胃腸道疾病/疾病(如腹瀉、結腸炎、過敏、壞死性小腸結腸炎和肥胖)的患者體內雙歧桿菌的數量明顯減少[44?45],這可能是通過宿主-微生物的相互作用或其與腸道微生物群其他成員的交互作用引起的,因此,雙歧桿菌的數量與多樣性越來越成為評價與預測機體的狀態的指標。
2.1 緩解腦腸綜合征
隨著對腸道菌群-腸-腦軸研究的不斷深入,人們已經發現腸道菌群可通過直接或間接的途徑影響嚙齒動物的大腦發育與行為,對機體健康產生重要的影響。益生菌可通過迷走神經、神經免疫系統、神經遞質、微生物代謝產物等途徑調節腸道菌群組成,從而緩解神經性疾病。目前,已知可用于防治神經性疾病的益生菌大部分屬于乳酸菌[46],而作為人類腸道發育早期優勢菌長雙歧桿菌也被發現具有積極調控作用。
2.1.1 治療IBS 腸易激綜合征(irritable bowel syndrome,IBS)與胃腸動力異常、腦腸調控異常、內臟感覺異常、精神心理等有關,發病機制尚未明確。胃腸道感染為誘發IBS 的病因,稱之為感染后IBS(post-infectious irritable bowel syndrome,PI-IBS)。益生菌近年來也被用于治療IBS,如長雙歧桿菌,但其作用機制目前尚未明確[47]。長雙歧桿菌特異性菌株的結合在逆轉腸道菌群失調方面起著重要作用。這種協同作用可使患者在胃腸道不良條件下存活,與腸粘膜粘連,調節免疫活性,恢復腸道環境。Bonfrate 等人發現長雙歧桿菌B.longumBB536、植物乳桿菌L.rhamnosusHN001 與B6 維生素復合使用,可改善IBS 患者的癥狀和病情,恢復腸道通透性,改善腸道菌群[48]。錢偉等人發現長雙歧桿菌可能通過抑制炎癥小體NLRP6,下調IL-18,從而降低PI-IBS 內臟高敏感性[47]。
2.1.2 緩解抑郁癥 抑郁癥是是一種嚴重的情感障礙性疾病[46],約60%的自殺者患有抑郁癥[49]。抑郁癥患者常郁悶不樂,并表現出嚴重的自殺傾向[50]。腦內促炎細胞因子白細胞介素-1(IL-1)水平升高會影響抑郁癥的各個方面,包括行為癥狀、腎上腺皮質激活和神經發生減少,此外,抑制性神經遞質5-羥色胺和GABA,可以參與調節機體情緒,當其水平下降時,便會引發機體抑郁[51]。目前,已有實驗證明長雙歧桿菌菌能夠抑制應激所導致的抑郁:Pinto-Sanchez 等人連續兩周給44 名IBS 患者每日服用1010CFU/mL 長雙歧桿菌B.longumNCC3001,研究發現B.longumNCC3001 通過減少與情緒處理和改善患者生活質量相關的多個大腦區域對恐懼刺激的反應來降低抑郁[52]。另一研究表明,瑞士乳桿菌和長雙歧桿菌通過減少促炎細胞因子和恢復腸道屏障完整性來共同改善心梗后抑郁癥[53]。未來長雙歧桿菌可能會成為防治抑郁癥的手段之一,這些證據將會為治療抑郁等情感障礙疾病提供潛在的新途徑。
2.1.3 改善認知功能損傷 腸道共生菌群可能通過免疫、內分泌和神經系統途徑調節腸道-大腦軸。這導致人們對益生菌調節腸道菌群以靶向中樞神經系統(CNS)功能和改善人類行為的興趣增加,特別是在壓力、情緒、焦慮和認知等領域。長雙歧桿菌B.longum1714TM在臨床前研究中已被證明可減少應激相關行為,改善健康志愿者的應激反應和認知功能[54],可調節與增強智力和減少精神疲勞相關的神經活動。B.longum1714TM在社會壓力中調節神經反應可能與激活大腦應對中心對抗調節負面情緒有關[55]。
Hae 研究發現抗炎長雙歧桿菌B.longumNK46可以通過調節微生物群LPS 介導的NF-κB 通路的激活來改善腸道菌群失調并抑制的LPS 產生,從而減輕認知功能衰退。抗炎長雙歧桿菌B.longumNK46 可抑制5×FAD-Tg 小鼠海馬中β淀粉樣蛋白、β/γ-分泌酶和細胞消亡蛋白酶-3 的表達以及β淀粉樣蛋白積聚,同時降低5×FAD-Tg 小鼠和老齡小鼠lba1+、LPS+/CD11b+和caspase-3+/NeuN+細胞群數量,抑制了這兩種小鼠海馬中NF-κB 通路,從而促使腦源性神經營養因子(BDNF)表達增加[56]。
2.2 緩解免疫相關疾病
自身免疫疾病是指機體對自身抗原發生免疫反應而導致自身組織損害的疾病。而長雙歧桿菌可調控腸道免疫、緩解免疫相關疾病,長雙歧桿菌發揮益生作用的機制主要與其代謝產物胞外多糖、結構物質肽聚糖及脂磷壁酸有關。
長雙歧桿菌胞外多糖(EPS)在宿主免疫調節中起著關鍵作用[57]。EPS 是釋放到細胞外滲透到培養基中的多糖,能夠調節機體免疫力,是一類效果優良的免疫調節劑,它與抗氧化劑、抗癌劑、免疫調節劑和降膽固醇活性都相關[58]。一項體外研究表明,長雙歧桿菌B.longumW11 的EPS 可增加促炎細胞因子IFN-γ,ConA 以兩種不同濃度(100 和200 g/L)刺激PBMCs 中的IL-1β 和IL-6,并增加IL-6(200 g/L)的產生,抗炎細胞因子包括未刺激PBMCs 中的IL-10(100 g/L)[59]。
另外,長雙歧桿菌細胞壁主要成分肽聚糖(Whole Peptidoglycan,WPG)是一種多糖與肽聚糖聚合而成的袋狀結構,它保持了細菌細胞壁的完整性,不論在體內或是體外均可表現出增強體液免疫和細胞免疫,活化巨噬細胞,激活多核白細胞及單核的吞噬活性等免疫活性作用。同時它還有抗腫瘤和抗感染等多種重要生理功能[60]。故不少學者把WPG 作為發揮免疫調節作用的主要活性物質之一。付艷茹等人研究發現完整肽聚糖可促進淋巴細胞轉化、延長生命期增加、Bcl-2 陽性表達率減小而Bax 基因表達增加、提高抑瘤率[61]。
脂磷壁酸又稱膜磷壁酸(Lipoteichoic acid,LTA),是革蘭陽性菌胞壁的共有成分,其主要結構是由16-40 個單體組成的1,3-鏈聚磷酸甘油骨架[62]。B.longumNQ-1501 的膜磷壁酸可刺激體液免疫,促進T 細胞和B 細胞的增殖[63]。然而也有文獻指出LTA 與DCA 協作,可以提高COX-2 表達,而COX-2 誘導的前列腺素E2(PGE2)可以通過EP4 受體抑制抗腫瘤免疫。
2.2.1 緩解過敏 過敏反應是指已產生免疫的機體在再次接受相同抗原刺激時所發生的組織損傷或功能紊亂的反應,引起過敏反應的抗原包括食入式、吸入式等來源的抗原。嬰幼兒食乳為主,然而兒童胃腸道免疫屏障發育不成熟,易發過敏性腹瀉等異常或強烈的免疫反應,例如牛奶蛋白過敏(cow’s milk protein allergy,CMPA)。研究發現長雙歧桿菌B.longumBBMN68 對于緩解兒童CMPA 有積極作用,其機制如圖2 所示[64]。此外,B.longumBBMN68 還可降低患兒外周血嗜酸性粒細胞(EOS)水平及血清免疫球蛋白E(IgE)、炎癥因子 IL-4、IL-6、IL-1β、水平[65]。長雙歧桿菌的干預可保留牛奶蛋白的營養價值,同時維持機體對乳脂肪和碳水化合物的正常攝入[66]。
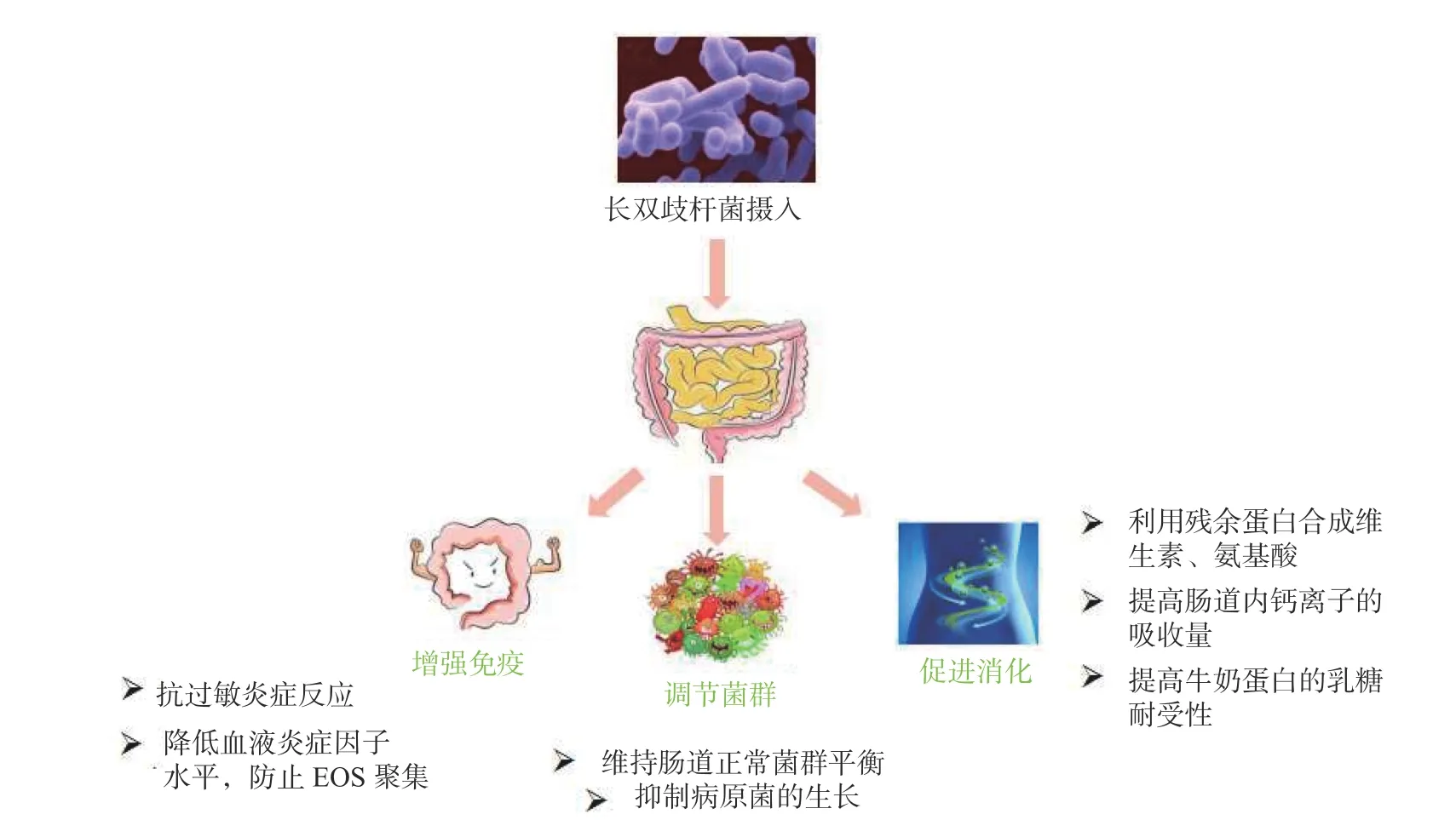
圖2 長雙歧桿菌治療兒童CMPA 主要作用機制[64]Fig.2 Main mechanism of Bifidobacterium longum in the treatment of CMPA[64]
楊景等人建立了以牛乳β-乳球蛋白(BLG)為過敏原的食物過敏小鼠模型,發現長雙歧桿菌B.longumBBMN68 可調節體內Th1/Th2 細胞失衡。由體內與體外實驗可知B.longumBBMN68 從促進機體CD103+DCs 分化及誘導DCs 半成熟狀態兩個方面調節DCs 功能,緩解過敏癥狀,促進免疫抑制[11]。
Lyons 等人發現B.longumAH1206 可增加幼年、成年和無菌動物體內Foxp3 1 T 調節細胞數量,抑制與抗原提呈、TLR 信號和細胞因子分泌相關的基因表達,提高與維持甲酸代謝相關基因的表達,從而對OVA 致敏動物氣道炎癥起到保護作用[67]。
綜上,長雙歧桿菌對腸道、呼吸道的過敏反應有緩解作用,同時,腸內和腸外疾病如過敏反應都與長雙歧桿菌的數量變化有關,健康兒童(30.3%)和過敏性疾病兒童(11.1%)之間的長雙歧桿菌水平存在統計學上的顯著差異[68]。因此,可以通過對腸道菌群中長雙歧桿菌含量的變化來預測過敏疾病的發生,也可以通過益生菌制劑對腸道菌群進行干預,提高長雙歧桿菌含量,從而實現對過敏癥狀的緩解作用。
2.2.2 緩解結腸炎 結腸炎是一種主要累及直腸、結腸黏膜和黏膜下層的非特異性炎性腸病,益生菌可通過調節免疫反應抑制炎癥、增強腸道黏膜屏障、調節氧化應激反應進行干預。長雙歧桿菌可通過調節免疫有效緩解結腸炎。Schiavi 等人觀察到B.longum35624 sEPS-neg 株(一種EPSs 陰性突變株)在體外可促進人外周血單個核細胞的IL-12p70、IFN-γ和IL-17 分泌。長雙歧桿菌B.longum35624 可預防T 細胞轉移性結腸炎,并促進腸道IL-17+淋巴細胞增殖[69]。Srutkova 等人發現B.longum7952 菌株通過減少腸系膜淋巴結細胞分泌TNF-α和IFN-γ,增加結腸中zonulin-1 和閉塞素,減輕急性結腸炎[70]。植物乳桿菌LC27 和長雙歧桿菌B.longumLC67 抑制脂多糖刺激的巨噬細胞中NF-κB 的激活和TNF-α的表達,減輕乙醇誘導的胃炎和肝損傷、肥胖和2,4,6-三硝基苯磺酸誘導的小鼠結腸炎和肝損傷[71?74]。
3 長雙歧桿菌疾病干預的應用
目前,基于對長雙歧桿菌特性的研究日益深入,長雙歧桿菌已被廣泛用于食品生產中,包括發酵酸奶、發酵型果汁、合生元制劑、微生態制劑等。此外,長雙歧桿菌在醫療保健方面也發揮重要作用。
3.1 長雙歧桿菌作為基因治療載體的應用
實體瘤獨特的厭氧現象使普通抗腫瘤治療難以發揮作用,長雙歧桿菌作為厭氧菌的一種,可利用趨低氧特點在腫瘤組織內選擇性定植,具有安全性及較好的靶向性,可基因治療的主要載體[75]。因此采用酶前體藥物策略,利用長雙歧桿菌攜帶無活性的前體藥物,可獲得高水平的厭氧菌靶向定植及前體藥物激活的實驗結果[76]。Xu 等將內皮抑素基因分別轉化青春雙歧桿菌和長雙歧桿菌,經尾靜脈推注和口服途徑給予實體瘤小鼠,發現雙歧桿菌高靶向性定植于瘤組織中,且強烈抑制腫瘤血管生長,延長了帶瘤小鼠的存活時間[77]。
3.2 富硒長雙歧桿菌緩解損傷的應用
微量元素硒(selenium,Se)是人體必需元素,與蛋白質組成硒蛋白維持生理功能,與25~30 種遺傳特異性酶組成硒酶維持催化功能,在生物化學和生理學上具有重要作用。在高硒培養基中發酵培養雙歧桿菌,通過特定條件使培養基中的無機硒吸收轉化為自身硒蛋白硒代胱氨酸,制備具備硒功能和雙歧桿菌功能的富硒益生菌(Selenium-enrichedBifidobacterium.longum,Se-BL)[78]。飲食中增加硒能夠通過減少氧化應激,來緩解熱應激所導致的豬腸上皮屏障受損[79]。有機硒具有低毒性和可接受的生物利用度,可作為化療預防劑,以防止抗癌藥物伊立替康的毒性,并協同增強體外和體內的抗腫瘤治療效果。
富硒益生菌綜合了微量元素硒和益生菌的優勢,改善雄性小鼠繁殖能力;改善小豬對高溫環境的適應性、調節腸道菌群,改善免疫功能及抗氧化狀態;改善高脂飲食導致的小鼠脂代謝紊亂及肝損傷[80]。富硒益生菌可影響體內硒蛋白基因表達、谷胱甘肽過氧化酶的活性和免疫功能,與CPT-11 聯用后可以增強抗腫瘤作用,能夠緩解四氯化碳引起的小鼠肝纖維化[81]。然而目前關于富硒益生菌的研究多集中于乳酸桿菌,對于富硒長雙歧桿菌的研究仍處于起步階段[82],現富硒長雙歧桿菌的應用進行總結,見表1。現有實驗表明,富硒長雙歧桿菌黏附能力顯著提升,可能與細菌表面Tuf 黏附蛋白的改變有關[83]。
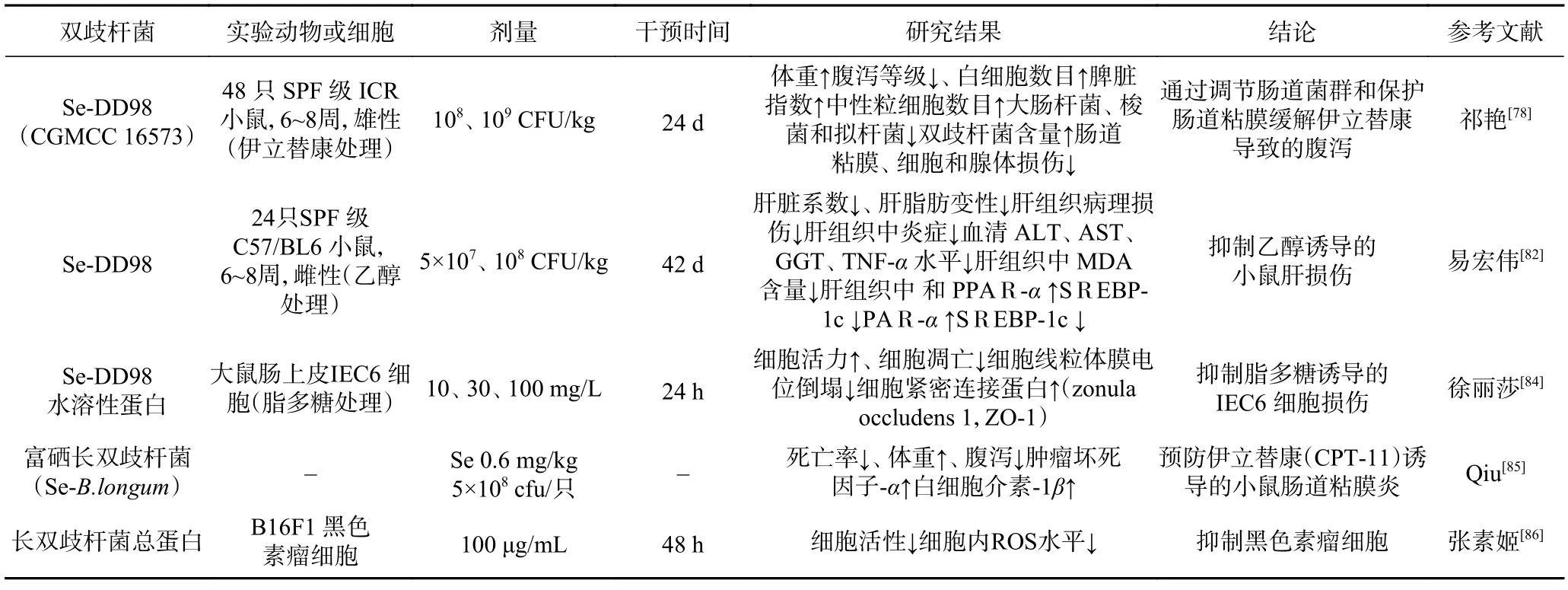
表1 富硒長雙歧桿菌的應用Table 1 Application of selenium enriched Bifidobacterium longum
4 結語
長雙歧桿菌作為一類獨特的益生菌,其嚴格厭氧、結腸定植等特性均顯著差異于其他菌株,這也賦予其特有的益生功能,對于開發并使用長雙歧桿菌制品來維持宿主健康具有積極意義。目前,盡管長雙歧桿菌在益生功能的研究方面已然取得了一些進展,但益生菌的研究已進入到組學時代,基于基因組、代謝組、蛋白組學水平對長雙歧桿菌的相關研究尚處于起步階段,在由表象研究深入到內在分子作用機理的研究有待于進一步深入。其次,已知投入商業化生產的長雙歧桿菌種類太匱乏,而通過臨床研究證明具有開發成為益生菌潛力的長雙歧桿菌并不僅限于已投產的有限菌株,這引人思考。此外,根據長雙歧桿菌的菌株特異性及不同疾病患者的實際情況有針對性地選擇長雙歧桿菌制品,考慮復合益生菌與合生元產品的開發,可大大提高疾病干預效率。長雙歧桿菌的研究為微生態學、食品衛生學、生理病理學、營養保健科學等交叉學科發展提供一種新的思路。
