東方百合‘索邦’幾丁質酶基因的克隆與表達分析
2021-06-22 00:46:32汪蓮娟韓蕊蓮
西北植物學報 2021年5期
崔 祺,吳 凡,詹 鵬,汪蓮娟,韓蕊蓮, 3,吳 昀*
(1 浙江理工大學 建筑工程學院,杭州310018;2 花卉種質創新與分子育種北京市重點實驗室,國家花卉工程技術研究中心,城鄉生態環境北京實驗室,教育部林木花卉育種實驗室,北京林業大學 園林學院,北京100083;3 西北農林科技大學 草業與草原學院,陜西楊陵 712100)
百合屬于百合科(Liliaceae)百合屬(Lilium)多年生具有地下鱗莖的單子葉草本植物,是重要的商品花卉和園林綠化植物。近年來,隨著從國外引進種球數量的不斷增加,中國百合栽培面積也在不斷擴大。然而,各種病害的發生對百合生產的危害不容小覷,其中由灰霉菌(Botrytiselliptica)引起的灰霉病是最為普遍發生的病害。B.elliptica專性侵染百合的莖、葉及花器官,病害發生后常常出現葉片焦枯、莖稈軟腐、花苞畸形、花瓣萎蔫等癥狀,嚴重時可造成種植區內百合成片死亡,是切花栽培和種球繁育的重要障礙[1]。生產實踐證明,應用抗病品種是最經濟有效又符合環保要求的病害防治措施。
前人對百合種質資源進行了灰霉病的抗性評價,發現部分野生百合,如岷江百合(L.regale)、宜昌百合(L.leucanthum)、大理百合(L.taliense)和湖北百合(L.henryi)[2],以及栽培品種中的東方百合和OT(Oriental × Trumpet)百合雜種系對灰霉病的抗性較強,亞洲百合和喇叭百合系列品種則較為感病[3]。目前少數百合對灰霉病表現出一定的抗性,并不存在完全免疫的種或品種。通過挖掘高抗種質材料中參與抗灰霉病反應的關鍵基因,并利用植物基因工程將其轉入百合中,培育抗病新品種,已成為防治百合病蟲害,促進百合產業健康發展的重要方向之一。
植物幾丁質酶(chitinase)是一類由多基因家族編碼的糖苷水解酶(glucoside hydrolase),各類同功酶的酶活性、氨基酸序列、保守結構域、蛋白等電點、功能位點、亞細胞定位等存在一定差異[4]。前人根據氨基酸的序列特性,將其分為糖苷水解酶18家族(GH-18)和糖苷水解酶19家族(GH-19),這兩大家族又進一步分為5類(Ⅰ-Ⅴ),其中Ⅰ類、Ⅱ類和Ⅳ類屬于GH-19家族,Ⅲ類和Ⅴ類屬于GH-18家族[5]。正常狀況下,幾丁質酶表達量很低或者不表達,但在受到病原菌或昆蟲感染時,其含量迅速升高,通過水解病原菌細胞壁或昆蟲外骨骼的幾丁質達到抑制其生長與增殖的目的[6]。目前已從擬南芥(Arabidopsisthaliana)[7]、水稻(Oryzasativa)[8]、水仙(Narcissustazetta)[9]和菊花(Chrysanthemummorifolium)[10]等多種植物中分離獲得幾丁質酶基因。玉米(Zeamays)[11]、番茄(Solanumlycopersicum)[12]、芥菜(Brassicajuncea)[13]和馬鈴薯(Solanumtuberosum)[14]等植物的幾丁質酶家族成員已經在全基因組范圍內得到了鑒定。
前人研究發現灰霉菌侵染能有效誘導辣椒(Capsicumannuum)[15]、草莓(Fragaria×ananassa)[16]和葡萄(Vitisvinifera)[17]中幾丁質酶基因的表達。幾丁質酶基因也可由非生物脅迫如鹽堿[18]、寒冷[19]和干旱[20],以及植物激素茉莉酸(jasmonic acid, JA)和水楊酸(salicylic acid, SA)誘導表達[14]。將幾丁質酶基因轉入檸檬(Citruslimon)[21]、矮牽牛(Petuniahybrida)[22]和番茄[23]中可增強轉基因植物對灰霉病的抗性。此外,轉化了幾丁質酶基因的植物對其他病害的抗性也得到了增強,如茶樹(Camelliasinensis)對茶餅病的抗性[24]、花生(Arachishypogaea)對黑斑病的抗性[25]、大豆(Glycinemax)對菌核病的抗性[26]以及蘋果(Malus×domestica)對炭疽病和褐斑病的抗性[27]。這些研究說明幾丁質酶基因在植物抗病反應過程中發揮著重要作用。
通過檢索NCBI數據庫,百合屬中只有岷江百合與麝香百合(L.longiflorum)登錄了幾丁質酶基因的全長cDNA序列,同其他植物相比較,有關百合幾丁質酶基因生物學功能的研究鮮有報道。本課題組前期通過分析灰霉菌侵染后的百合葉片轉錄組數據,發現了2個差異表達的幾丁質酶基因家族成員,分別為Ⅲ類和Ⅴ類成員。對Ⅴ類成員進行qRT-PCR分析發現:接種灰霉菌12和24 h后,該基因在抗病百合中的表達水平分別為感病百合的8和4倍,推測其可能參與到百合的抗病反應中[28]。鑒于幾丁質酶基因在植物抗病反應中的關鍵作用,進一步開展百合幾丁質酶基因家族其他成員的研究,具有潛在的理論與現實意義。因此,本研究以高抗品種東方百合‘索邦’(L.oriental hybrid ‘Sorbonne’)接種灰霉菌12 h后的葉片為材料,利用RT-PCR技術克隆到了上述差異表達的Ⅲ類幾丁質酶基因成員,命名為LoChi2,通過生物信息學手段預測分析了目標基因推導的編碼蛋白的結構和功能,同時借助qRT-PCR的方法,對灰霉菌侵染以及SA/JA處理條件下LoChi2基因在百合中的表達模式進行了研究,以期為百合幾丁質酶基因的抗病功能研究奠定基礎。
1 材料和方法
1.1 實驗材料及處理
本研究選取的植物材料為課題組收集的東方百合‘索邦’、新鐵炮百合‘雷山3號’以及亞洲百合‘穿梭’,它們分別對灰霉病的抗性水平為高抗、中感和高感[3],將其種植于浙江理工大學人工氣候室中,栽培期間為長日照條件(光照/黑暗:16 h/8 h),晝/夜溫為 25 ℃/18 ℃,通風良好,定期澆水。灰霉菌(B.elliptica, 菌株編號為ACCC36423)由百合葉片上分離純化獲得,購買于中國農業微生物菌種保藏管理中心。菌株接種在馬鈴薯葡萄糖瓊脂培養基上,并將培養基置于25 ℃的恒溫培養箱中黑暗培養,10 d后用于百合葉片的接種試驗。
在百合現蕾期,分別選取上述3個品種健康完整無病害的葉片進行菌塊接種。首先將葉片放入底面鋪有2層濕潤紗布的培養皿中,用無菌打孔器從培養基中切取大小一致的菌塊(菌絲面朝下)接種于葉片背面,對照接種大小一致、不含灰霉菌的培養基,將接種后的葉片置于塑料托盤中,噴灑少量無菌水后使用保鮮膜密封保濕,于25 ℃條件下的培養箱中黑暗培養。依據前期研究結果[29],分別于未接種(0 h),接種6、12、24、48和72 h后收集葉片。為探究LoChi2基因能否對外源激素做出響應,使用0.1 mmol/L SA和0.1 mmol/L JA溶液分別噴灑‘索邦’百合葉片,對照噴灑無菌水,采樣時間同上述灰霉菌處理一致[30]。所有處理在同一時間點均收集3片葉子,各時間點重復3次,液氮速凍后將樣品保存在-80 ℃冰箱中以備提取RNA進行后續試驗。
1.2 方 法
1.2.1 RNA的提取與cDNA的合成使用RNA快速提取試劑盒(RN38,艾德萊生物,北京)提取百合葉片中的總RNA,操作流程參照試劑盒說明書。RNA濃度與純度通過NanoDrop 2000分光度計(Thermo Fisher Scientific,美國)與1%瓊脂糖凝膠電泳確認,檢測合格的RNA進行cDNA合成,操作方法參見反轉錄試劑盒(FSQ-301,Toyobo,日本)說明書。
1.2.2LoChi2基因編碼序列的克隆根據轉錄組測序獲得的LoChi2基因序列,設計克隆引物F1/R1(表1),進行PCR擴增。擴增反應體系為50 μL,包含1 μL cDNA模板、5 μL dNTP(2 mmol/L)、3 μL MgCl2(25 mmol/L)、5 μL PCR 緩沖液(10×)、1 μL高保真PCR酶(1 U/μL)(KOD-401,Toyobo,日本)、32 μL ddH2O以及上下游引物(10 μmol/L)各1.5 μL。擴增程序為:94 ℃預變性2 min, 之后98 ℃ 10 s、68 ℃ 1 min進行45個循環反應,最后68 ℃延伸10 min。割膠回收PCR產物,將回收的cDNA依據試劑盒說明書(VT205,天根生化,北京)連接到pLB載體上。隨后將重組質粒轉入大腸桿菌感受態,菌液PCR鑒定出陽性克隆,委托北京睿博興科生物技術有限公司測序,獲得編碼序列。
1.2.3LoChi2基因系統進化分析基于幾丁質酶的氨基酸序列,采用MEGA6.0最大似然法的方式構建系統進化樹(Bootstrap = 1 000),以NCBI數據庫下載的17條擬南芥幾丁質酶序列(本研究將其編號為AtChi1-AtChi17,表2)為參考標準,對百合LoChi2及其同源基因進行分類。
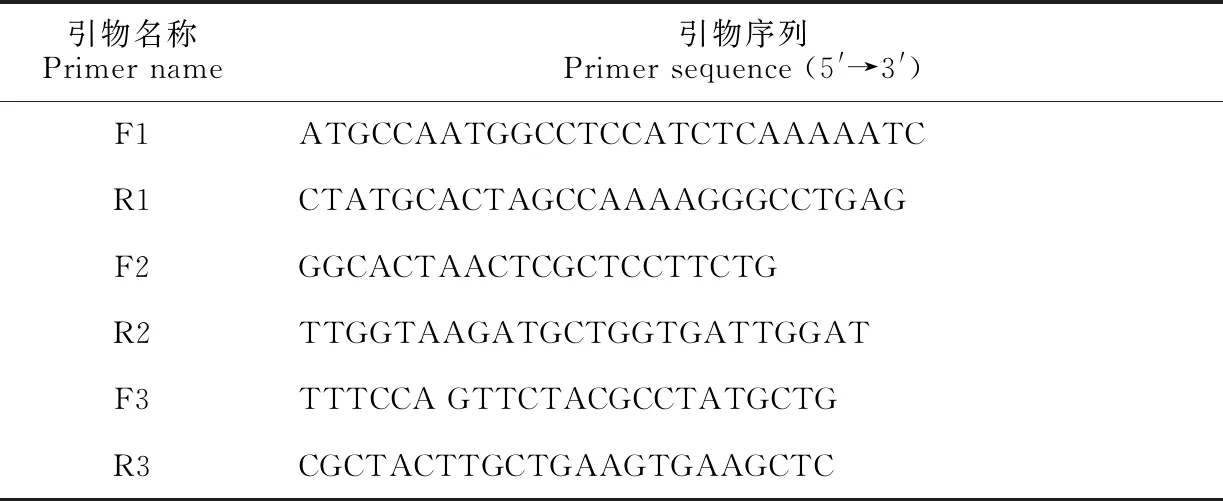
表1 引物序列表
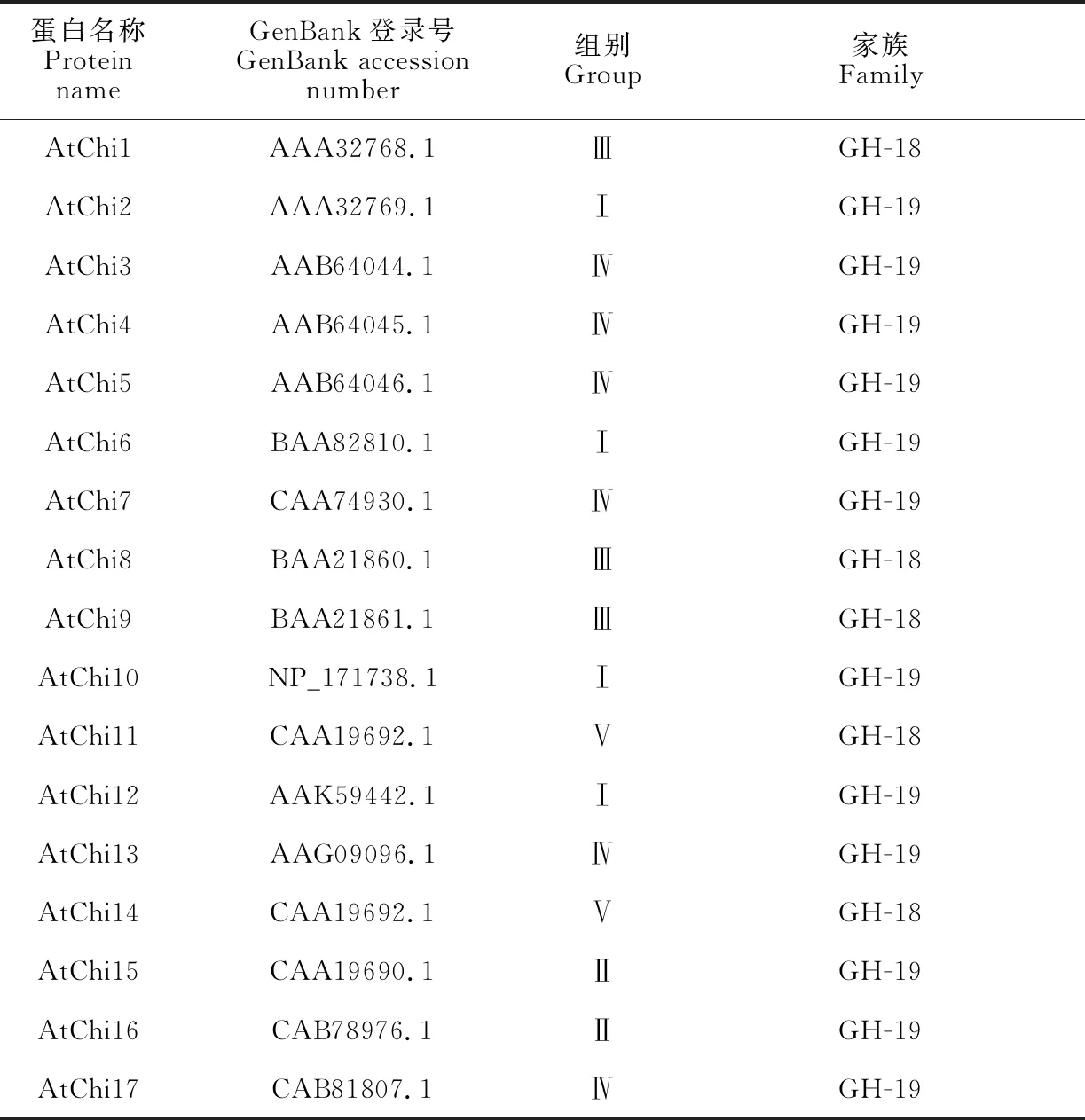
表2 擬南芥不同類別幾丁質酶信息
1.2.4 LoChi2蛋白的生物信息學分析利用在線網站(https://web.expasy.org/cgi-bin/protparam/protparam)預測LoChi2蛋白的理化性質。利用SignalP(http://www.cbs.dtu. dk/services/SignalP/)、NetPhos(http://www.cbs. dtu.dk/services/NetPhos/)和NetNGlyc(http://www.cbs.dtu.dk/services/NetNGlyc/)分別預測蛋白的信號肽、磷酸化和糖基化位點。利用TMPRED(https://embnet.vital-it.ch/software/TMPRED_form.html)預測蛋白的跨膜結構域。利用在線網站(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)和(https://swissmodel. expasy.org/interactive)分別預測蛋白的二級和三級結構。利用TargetP(http://www.cbs.dtu. dk/services/TargetP-1.1/index.php)進行亞細胞定位的預測。
1.2.5LoChi2基因的表達分析以百合真核延伸因子基因EF1(Eukaryotic elongation factor 1, NCBI登錄號KJ543461)為內參,分別設計LoEF1和LoChi2的熒光定量引物F2/R2和F3/R3(表1),以灰霉菌或SA/JA處理后的葉片組織cDNA為模板,參照SYBR Premix ExTaqTMⅡ(TaKaRa, 寶生生物,大連)說明書,配制qRT-PCR反應液,使用CFX Connect Real-Time PCR Detection System(Bio-Rad,美國)進行擴增反應,反應程序為:95 ℃預變性3 min, 隨后95 ℃變性10 s、60 ℃(LoChi2)/55.0 ℃(LoEF1)值條件下反應30 s,72 ℃延伸30 s,共進行40個循環反應,之后以每5 s升溫0.5 ℃的速度從65 ℃升至95 ℃,獲得擴增產物的熔解曲線。每份樣品進行3次技術重復,利用2-ΔΔCt計算LoChi2基因的相對表達量,數據以平均值±標準差(SD)的形式表示,在SPSS17.0中通過單變量方差分析(One-way ANOVA)計算不同處理與對照之間基因表達水平的顯著性差異。
2 結果與分析
2.1 LoChi2基因的克隆、氨基酸序列比對與系統進化分析
采用RT-PCR的方法,以‘索邦’百合葉片cDNA為模板,獲得了一條清晰的擴增條帶,片段大小與預期相一致,經純化回收、連接轉化和測序,成功得到‘索邦’幾丁質酶基因完整的編碼區序列,長度為915 bp,編碼304個氨基酸。將其氨基酸序列提交至NCBI在線Blastn檢索,下載同源性較高的候選序列,使用DNAMAN 9.0軟件進行序列的多重比對。如圖1所示,‘索邦’幾丁質酶基因編碼的氨基酸序列與同屬植物麝香百合LlChi2的氨基酸序列同源性最高,其次是菠蘿(Ananascomosus)AcChi2、梅花(Prunusmume)PmChi2和煙草(Nicotianatabacum)NtChi2,序列相似度分別為90%、64%、63%和62%,因此將其命名為‘索邦’LoChi2,NCBI登錄號為MW310626。通過分析LoChi2與其同源基因編碼氨基酸序列的保守結構域,發現他們無幾丁質酶結合結構域,無C-端延伸區,但具有GH-18家族典型的結構特征:存在1個GH-18 narbonin催化結構域及8個糖類物質結合位點(圖1)。
為進一步明確LoChi2在幾丁質酶家族中所屬類別,本研究構建了幾丁質酶氨基酸序列的系統發育樹(圖2),該系統進化樹包含了擬南芥幾丁質酶GH-18和GH-19家族共5類(Ⅰ~Ⅴ)成員,依據擬南芥中的分類情況,百合LoChi2及其同源基因AcChi2和PmChi2與擬南芥GH-18家族第Ⅲ類成員AtChi1、AtChi8和AtChi9親緣關系較近,聚為一簇,與其他結構類型的幾丁質酶關系較遠,表明LoChi2屬于GH-18家族Ⅲ類幾丁質酶成員。
2.2 LoChi2蛋白結構特征與亞細胞定位分析
蛋白質的理化性質分析表明LoChi2蛋白的分子量為32.52 kD,原子組成為C1480H2204N358O461S4,理論等電點為4.16,不穩定系數為27.43,平均親水性為0.10,預測LoChi2是一個穩定的疏水蛋白。功能位點預測發現LoChi2存在1個潛在的信號肽斷裂位點,1個糖基化位點以及多個磷酸化位點。跨膜結構域預測顯示LoChi2蛋白共有10個跨膜區域,其中有7個由內到外的蛋白跨膜區域和3個由外到內的跨膜區域,推斷LoChi2屬于跨膜蛋白(圖3,A)。另外,二級結構預測表明LoChi2含有38.82%的α-螺旋,18.09%的β-折疊,17.76%的延伸鏈,36.84%的不規則卷曲;進一步分析其三維結構,發現LoChi2的立體結構外形類似橢圓狀(圖3,B),與目標模板(SMTL ID: 4rl3.1)結構的相似度為60.53%,主要結構元件為α-螺旋和不規則卷曲,與二級結構預測結果一致。同時對LoChi2進行亞細胞定位分析,結果顯示該蛋白作為分泌蛋白分泌于細胞外的可能性最大,概率高達99%,推測LoChi2可能定位于細胞外。
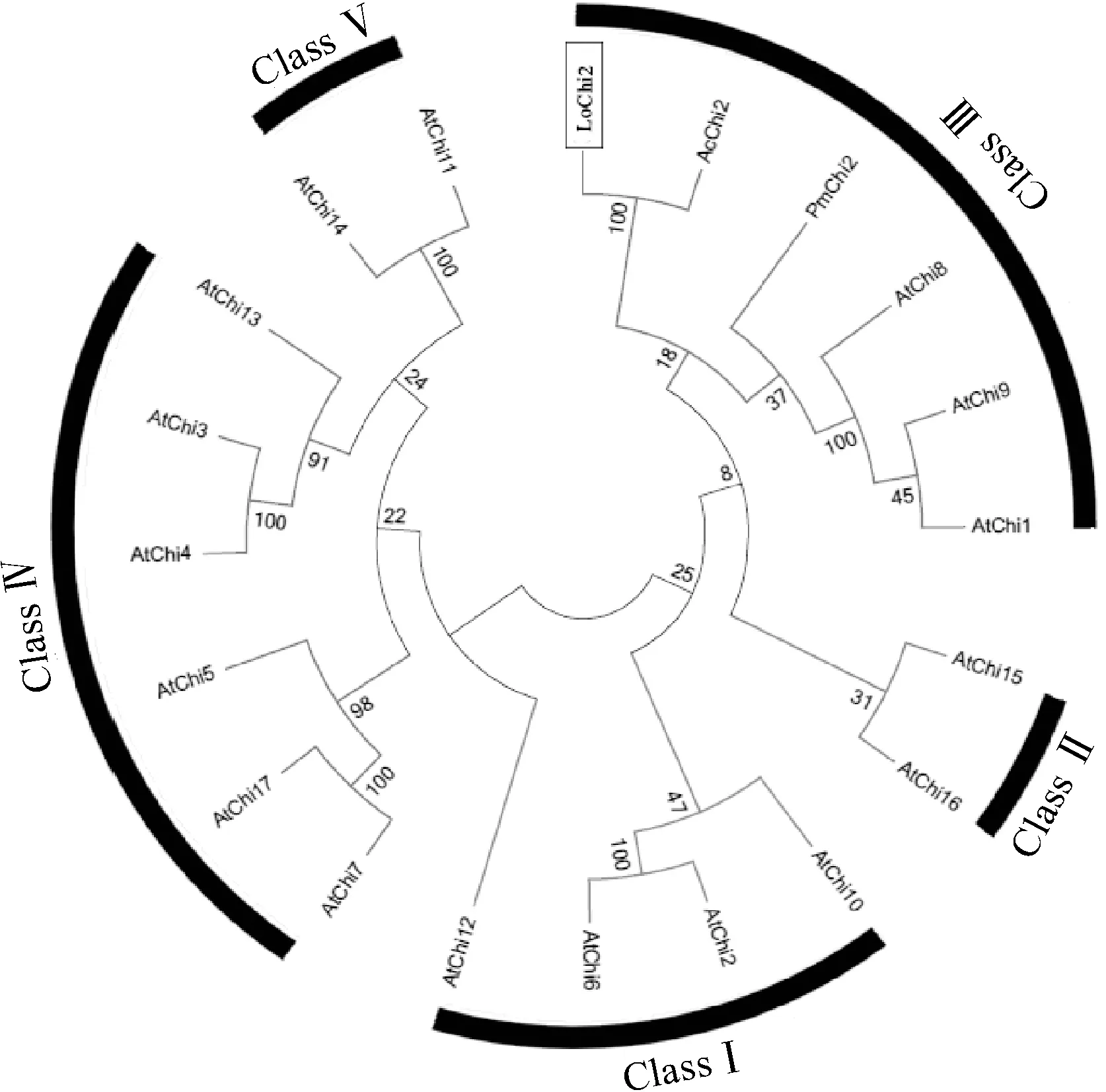
圖2 百合與其他植物幾丁質酶基因編碼氨基酸 序列的系統進化樹Fig.2 Phylogenetic tree based on the amino acid sequences of chitinase genes from lily and other plants

圖3 LoChi2蛋白質跨膜區(A)及其三級結構(B)的預測Fig.3 Transmembrane (A) and tertiary structure (B) predictions of LoChi2 protein
2.3 不同抗性百合接種灰霉菌后LoChi2基因的表達分析
為比較不同抗性百合‘索邦’、‘雷山3號’和‘穿梭’中LoChi2基因在轉錄水平上響應灰霉菌侵染的差異,本研究對這3種百合進行了qRT-PCR分析。結果表明:接種灰霉菌后,LoChi2基因在3種百合中均能被誘導,表達量各有不同程度的增加(圖4)。在高感百合品種‘穿梭’中,接種6 h后LoChi2的表達量微微上調,24 h后達到峰值,表達量與對照相比增加了2.6倍,在這之后表達量顯著降低,差不多降至初始水平。而在中感和高抗百合中,表達量在接種6 h后顯著增加,并在之后維持著較高的水平。其中,中感百合‘雷山3號’在整個接種期呈現先增加后降低的表達趨勢,接種48 h后表達量最大,約為對照的15倍;高抗百合‘索邦’在整個接種期內表達量較為多變,呈現先增加后降低,再增加又降低的折線變化趨勢,于接種12 和48 h后出現峰值,分別為對照的25 和23倍。整體來看,接種灰霉菌12、24、48和72 h后,‘索邦’中LoChi2基因表達量的增加幅度要明顯高于‘雷山3號’和‘穿梭’,分別為‘雷山3號’的4.5、1.3、1.5和1.5倍,‘穿梭’的25.0、3.7、20.9和10.3倍。
2.4 SA/JA處理條件下LoChi2基因在百合中的表達分析
許多幾丁質酶家族成員通過植物激素信號轉導途徑參與植物抗病反應,因此進一步分析了LoChi2基因對外源激素JA和SA的響應情況。結果表明:LoChi2基因在SA和JA分別處理后的‘索邦’葉片中均有不同程度的上調表達(圖5)。JA處理6 h后,LoChi2的表達量顯著增加,在12 h處微微下調,之后又迅速增加,48 h達到峰值,約為對照的7.9倍。相比而言,LoChi2對SA的響應要遲緩,SA處理6 和12 h后,LoChi2的表達量變化較小,直至24 h后開始明顯增加,78 h表達量增至最高,約為對照的8.1倍。
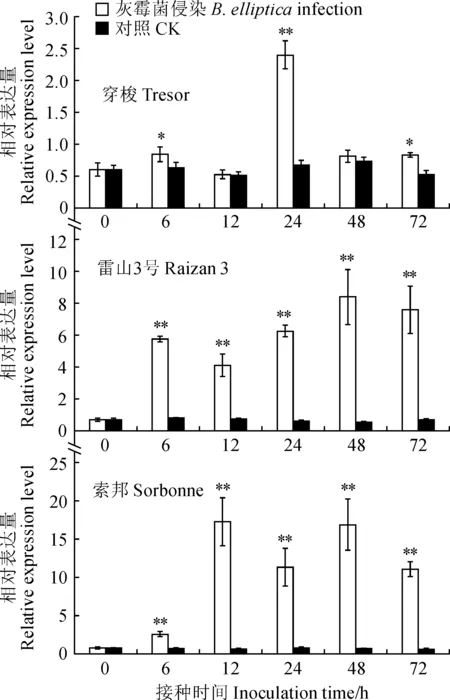
星號代表處理與對照之間基因表達水平存在顯著性差異 (*. 0.01 < P < 0.05, **. P < 0.01)。下同圖4 不同抗性百合接種灰霉菌后葉片中 LoChi2基因的表達分析Asterisks indicate significant differences of gene expression levels between treatments and CK (*. 0.01 < P < 0.05, **. P < 0.01). The same as belowFig.4 Expression analysis of LoChi2 gene observed from lily leaves with different levels of resistance after inoculation with B. elliptica.
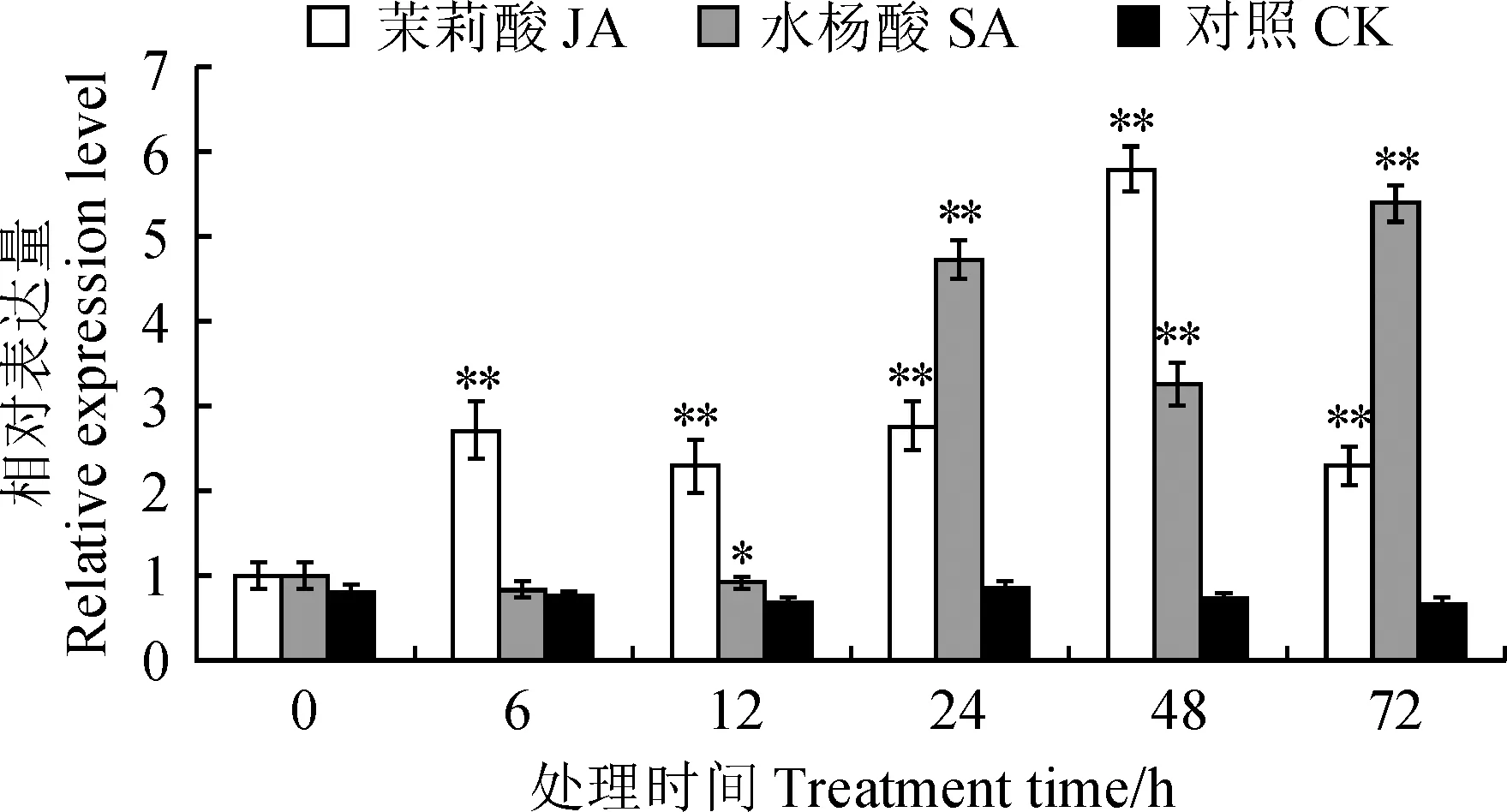
圖5 JA/SA處理下LoChi2基因的表達分析Fig.5 Expression analysis of LoChi2 gene in response to JA/SA treatments
3 討 論
植物受病原菌侵染后,誘導表達的幾丁質酶蛋白能夠直接與病原體的侵染結構接觸,抑制、干擾病菌的生長進程,激發植物產生系統獲得性抗性,發揮廣譜抗病作用。植物中不同類型的幾丁質酶在結構上存在較大差異,Ⅰ類幾丁質酶成員在N末端存在幾丁質結合結構域和C末端催化結構域,這兩個結構域中間包含一段鏈接區域。Ⅱ類成員的催化域同Ⅰ類具有較高同源性,但缺乏幾丁質結合域和鏈接區域。Ⅲ類和Ⅴ類成員與其他類成員序列相似度較低,但均具有GH18 narbonin催化結構域。Ⅳ類與Ⅰ類較為相似,但幾丁質結合結構域和催化結構域的長度均小于Ⅰ類[6]。本研究從東方百合‘索邦’中分離到了1個幾丁質酶基因LoChi2(MW310626)完整的編碼區序列,大小為915bp,編碼304個氨基酸殘基。蛋白保守結構域和系統進化分析發現LoChi2屬于GH-18家族的Ⅲ類幾丁質酶成員,存在保守的GH18 narbonin催化結構域、跨膜結構域以及信號肽、糖基化、磷酸化位點,預測為疏水的分泌蛋白,且定位于細胞外。這一結果與楊郁文等[31]研究類似,他們發現棉花(Gossypiumhirsutum)Ⅲ類幾丁質酶成員也為分泌蛋白,且具有信號肽位點,定位于細胞外。周潔等[32]在柳樹(SalixJiangsuensis2345)中獲得了1個Ⅲ類幾丁質酶基因的全長cDNA 序列,該序列編碼的蛋白為疏水蛋白,具有GH-18家族特征性催化結構域,進一步證明了不同植物中Ⅲ類幾丁質酶的保守性。此外,朱晨等[20]發現茶樹Ⅰ類幾丁質酶成員也具有多個磷酸化位點,由此推測不同類型幾丁質酶成員在蛋白翻譯中可能發生過多次磷酸化修飾。
病原菌侵染后,多數情況下抗病品種中幾丁質酶的表達量要高于感病品種。例如,Ebrahim等[33]發現接種鐮刀菌后,抗病芒果(Mangiferaindica)品種中幾丁質酶活性要遠遠高于感病品種,因此認為幾丁質酶活性可作為篩選抗病和感病品種的標記,在抗病和感病荔枝(Litchichinensis)品種中也存在同樣的現象[34]。基因表達方面,Vasanthaiah等[35]發現,葡萄幾丁質酶基因在抗病品種中的高效表達是其產生高抗性,抵御炭疽病的重要原因之一。同樣,棉花[31]和甘蔗[36](Saccharumofficinarum)幾丁質酶基因在抗性品種中表達水平也要高于感病品種。本研究發現,在高抗品種‘索邦’和中感品種‘雷山3號’葉片接種灰霉菌6、12、24、48和72 h后,LoChi2基因的相對表達量大幅度增加,且‘索邦’明顯高于‘雷山3號’,而高感品種‘穿梭’僅在接種6、24和72 h處有明顯增加,增加幅度較為平穩。百合中LoChi2基因的轉錄水平與品種間抗病性呈現出正相關,說明LoChi2是參與灰霉菌防御反應的關鍵抗病基因。
幾丁質酶的表達受抗病相關激素JA和SA的調節,異源表達某些幾丁質酶基因會導致轉基因植物激素信號通路的改變,從而影響其對病原菌的抵御能力。前人通過對馬鈴薯噴施JA和SA發現有3個幾丁質酶基因的表達量在處理24 h以內顯著增加[14],外源噴施SA和JA還可誘導茶樹[37]、花生[38]和小麥[39](Triticumaestivum)中幾丁質酶基因的表達,增強植株抵御病蟲害的能力。在擬南芥中過表達草莓幾丁質酶基因Chit2可通過激活SA通路關鍵防御基因PR1(Pathogenesis-related protein 1)以及JA通路關鍵基因PDF1.2(Plant defensin 1.2)的表達來提高轉基因植株對炭疽病的抗性[40]。本研究中,LoChi2基因表達量在JA和SA處理后的‘索邦’百合葉片中均有不同程度的增加,暗示了該基因可能在這兩條抗病信號通路中扮演著重要角色。
灰霉菌與植物的互作非常復雜,它是很多基因相互作用的過程[28],幾丁質酶作為一類重要的病程相關蛋白,在植物抗灰霉病轉基因育種中具有潛在的優勢。郭林霞等[23]發現轉杜仲(Eucommiaulmoides)幾丁質酶基因EuChit1番茄提高對灰霉病的抗性,與抗氧化酶SOD、POD和CAT活性提高以及病程相關基因PR1、PR2和PR5表達量上調密切相關。Núez deGonzlez等[41]將水稻幾丁質酶基因RCH10成功轉入了百合品種‘Star gazer’中,對9個轉基因株系的葉片接種灰霉菌后測定孢子數量,發現所有株系均極顯著低于野生型,各株系對灰霉病的抗性水平與RCH10基因的表達量呈正相關,并且在營養生長和生殖生長階段植株的表型性狀,包括株高、莖粗、葉片和種球大小以及花色、花量和花期并未發生明顯改變。類似研究在矮牽牛中也獲得了成功,轉幾丁質酶基因的矮牽牛對灰霉病抗性顯著強于野生型, 且葉片中蛋白提取液能夠有效抑制灰霉菌菌絲生長[22]。由此推測LoChi2基因在百合抗灰霉病育種方面具有廣闊的應用前景。
綜上所述,高抗百合品種‘索邦’中的幾丁質酶基因LoChi2具有植物GH-18家族Ⅲ類成員典型的結構特征。灰霉菌侵染后,LoChi2在抗病百合品種中的表達量顯著高于感病品種,且受外源激素JA/SA誘導表達。根據試驗結果推測LoChi2是參與百合灰霉病防御反應的關鍵抗病基因,隨著百合基因工程技術的不斷發展,LoChi2將成為百合抗灰霉病轉基因育種中新的候選基因。
