5種蝦脊蘭菌根顯微結構觀察
2021-06-22 00:46:36李曉芳葉紅環楊加文
西北植物學報 2021年5期
李曉芳,張 梅,徐 建,葉紅環,楊加文,周 艷,周 慶
(貴州省植物園,貴陽 550004)
蝦脊蘭屬(Calanthe)為蘭科(Orchidaceae)多年生地生草本植物,全屬約150種,中國有49種和5變種,主產于長江流域及以南各省區,貴州產26種1變種[1-2]。蝦脊蘭屬名在希臘語中意為“美麗的花”,因花朵唇瓣外翻,形似小蝦的尾巴而得名[1,3]。蝦脊蘭屬植株典雅清新,花型新穎獨特,花色豐富艷麗,具有較高的觀賞價值,被譽為“花中西施”[4-5]。此外,蝦脊蘭屬植物全草入藥,有清熱解毒、活血散結和止痛消腫等功效,可用于治療瘰疬、淋巴結核、跌打損傷、腰肋疼痛和痔瘡等疾病[6-7],具有較好的藥用價值。但由于蝦脊蘭屬植物一般生長在林下或林緣,為中型草本植物,且花大花多,色彩艷麗,因此長期的人畜干擾對蝦脊蘭屬野生資源及其生境造成了嚴重的破壞。因此,弄清5種蝦脊蘭菌根的顯微結構特點,菌根真菌的侵入途徑與分布特征等,以期為蝦脊蘭屬植物的保護及其內生真菌的利用提供參考。目前,國內外對蝦脊蘭屬的研究主要集中在種質資源調查與分類學[8-17]、遺傳多樣性[18-25]、組培繁育[26-37]、化學成分與藥用價值[6-7,38-40]以及保護生物學[41-44]等方面,僅蔣雅婷等[28]對無距蝦脊蘭營養器官的解剖結構及其生態適應性進行了初步的研究,除此之外,本屬其他種類營養器官形態結構的研究尚未見報道。本研究用石蠟切片法觀察5種蝦脊蘭菌根的顯微結構,并探討內生真菌在其根內的分布特征及其侵染方式,以期為蝦脊蘭屬的保護與內生真菌的研究奠定基礎。
1 材料和方法
所選材料來自貴州省植物園蘭花溫室5種蝦脊蘭植物的新鮮營養根(表1)。具體方法如下:取新鮮健康的營養根中部3~4 cm根段,每種3份,流水沖洗干凈,切成3~4 mm長的小段迅速放入FAA中固定48 h。用常規石蠟切片法,切片厚度為8~12 μm,番紅-固綠對染,中性樹膠封片,然后置于Nikon eclipse E100正置光學顯微鏡下觀察并拍照。
2 結果與分析
2.1 5種蝦脊蘭植物菌根顯微結構比較
觀察發現,5種蝦脊蘭成熟植株根簇生于假鱗莖上,根肉質,細長,粗細均勻,平均直徑多1~3 mm;根尖呈乳白色。由根中部橫切面可見,根由根被、皮層和中柱3部分組成(圖版 Ⅰ,1、4、7、10、13),只在細胞大小、層次及排列方式等方面存在差異,詳見表2。根被常由3~6層排列緊密且無細胞間隙的薄壁細胞(死細胞)組成,根被最外的一層細胞為表皮,表皮細胞壽命較短,常磨損脫落。部分根被細胞的外壁向外突出形成根毛,僅異鉤距蝦脊蘭無根毛。
皮層可分為外皮層、中皮層和內皮層3層,位于根被內側,由9~13層排列疏松、細胞間隙較大的大型薄壁細胞組成,占根部橫切面積的60%以上。外皮層是緊挨根被的細胞體積較大且排列規則的1層細胞,除通道細胞外,其余均為死細胞。中皮層由8~12層細胞組成,是活力最旺盛的細胞,其細胞較大,多角球形,內含物十分豐富,是菌絲侵入菌根后定殖的部位,部分細胞內明顯可見菌絲、菌絲結、淀粉粒、針狀結晶體等結構(圖版 Ⅰ,3、6、9、12、15)。其中,銀帶蝦脊蘭皮層細胞層數最多,有18~20層,這可能與銀帶蝦脊蘭所處生長環境腐殖質較多,根部能吸收更多營養有關。
中柱位于根的中心,由中柱鞘、維管組織和髓組成,其細胞小而密集,易與皮層區別。中柱鞘是中柱最外面的一層薄壁細胞。維管組織由韌皮部和木質部組成,韌皮部和木質部8~12束維管束輻射狀相間排列;木質部的發育方式為外始式,導管孔徑不一,靠近髓部的后生木質部導管孔徑較大;韌皮部靠近中柱鞘分布,細胞較小,團狀排列。髓較大,位于中柱中央,由較小的具細胞間隙的圓形薄壁細胞組成。
2.2 5種蝦脊蘭植物菌根真菌的分布特點
通過觀察可知,5種蝦脊蘭的根有蘭科植物典型的菌根結構,在皮層細胞中明顯可見菌絲和菌絲結分布,且5種蝦脊蘭的外皮層能明顯觀察到通道細胞,而菌絲侵入時也未破壞5種蝦脊蘭的根被,由此推斷,菌絲侵入的方式是通過根被細胞間隙進入通道細胞,穿過通道細胞后進一步向皮層擴展,被侵染的皮層細胞可觀察到大量菌絲與菌絲結(圖版 Ⅰ ,2、5、8、11、14)。侵入皮層的菌絲在皮層細胞內橫向和縱向生長,并在皮層組織細胞內形成大量著色較深、形狀不規則的菌絲結。皮層是菌絲和菌絲結分布的主要區域,菌絲只侵染部分皮層細胞,菌絲結在皮層組織中也分布不均,均在靠近外皮層的幾層皮層細胞中數量居多。菌絲在細胞核附近被消解,細胞核變形膨大。此外,5種蝦脊蘭的根被和中柱多少有些菌絲分布(圖版 Ⅰ,3、6、9、12、15中a、b),除劍葉蝦脊蘭外,其余4種中柱細胞中亦有菌絲結分布,說明這5種蝦脊蘭可能擁有廣泛的菌根真菌類型,且這些共生真菌的侵染能力較強,使它們能夠更好地適應不同的地理和氣候環境。

表1 實驗材料及來源

表2 5種蝦脊蘭根的顯微結構與菌根真菌的侵入途徑與分布特征
3 討 論
3.1 5種蝦脊蘭菌根的顯微結構特征
本研究通過組織切片法,從細胞學角度闡述了5種蝦脊蘭菌根的顯微結構,結果表明其與已報道的無距蝦脊蘭的根相似[28]。5種蝦脊蘭中,4種根毛稀疏,1種無根毛,無形成層,且維管組織所占比例較小,說明其地下部分的吸水和導水能力較弱。有研究表明,某些單子葉植物特有的根被組織具有良好的吸水、保水功能,其細胞層數、厚薄等隨環境水分變化而變化,如鐵皮石斛(Dendrobiumofficinale)菌根的培養基質水分質量分數降低至5.5%時,其根被細胞層數會多至5層以上,從而提高植株的御旱能力[45]。5種蝦脊蘭的根被細胞主要為3~8層,彌補了根輸導組織不發達的缺陷,這也是對土壤含水量充足的一種響應。
蘭科植物根木質部脊數從5~30束不等,5種蝦脊蘭的維管束數為8~12束,與春蘭(Cymbidiumgoeringii)(14束)、流蘇石斛(Dendrobiumfimbriatum)(15~17束)、長距石斛(D.longicornu)(21束)等[46]相比較少,導水率相對低下,與其發達的皮層組織、根被以及不夠發達的根毛相適應。維管束數的多少是植株對生長環境長期適應的結果,有些蘭花的木質部脊數波動幅度較大[47],其適應性有隨維管束的束數增多而增強的趨勢,維管束多的,對土壤的水濕條件要求不嚴,引種易于成活[48]。通過調查和實驗發現,5種蝦脊蘭對生境條件的適應能力較強,在潮濕山谷、溪邊、常綠闊葉林林下以及水分含量較少的巖石空隙或石礫較多的土面均能生長,引種成活率較高。
3.2 5種蝦脊蘭菌根真菌的侵染方式與特點
從5種蝦脊蘭的菌根結構來看,其外皮層均有通道細胞分布,且根被組織未被破壞,由此推斷,5種蝦脊蘭菌絲由通道細胞侵入皮層,并在皮層細胞中形成大量不規則的菌絲結。被侵染的細胞可觀察到大量菌絲與菌絲結,菌絲在細胞核附近被消解,細胞核變形膨大。皮層是菌絲、菌絲結侵染和定殖的主要區域,尤其是在靠近外皮層的幾層皮層細胞中數量居多,根被和中柱有菌絲分布,除劍葉蝦脊蘭外,中柱亦有菌絲結分布。皮層細胞通過消化侵入的菌絲及其新陳代謝活動產生的物質為蘭科植物生長提供營養,同時,又伴隨著新菌絲的生長和侵入,真菌也通過消化蘭科植物新陳代謝活動由根部分泌的及其與植物在消化與反消化的相互作用過程中產生的有機物和無機物,獲得菌絲生長所需的營養,從而確立并維持真菌與蘭科植物穩定的共生關系,進而形成菌根共生體。
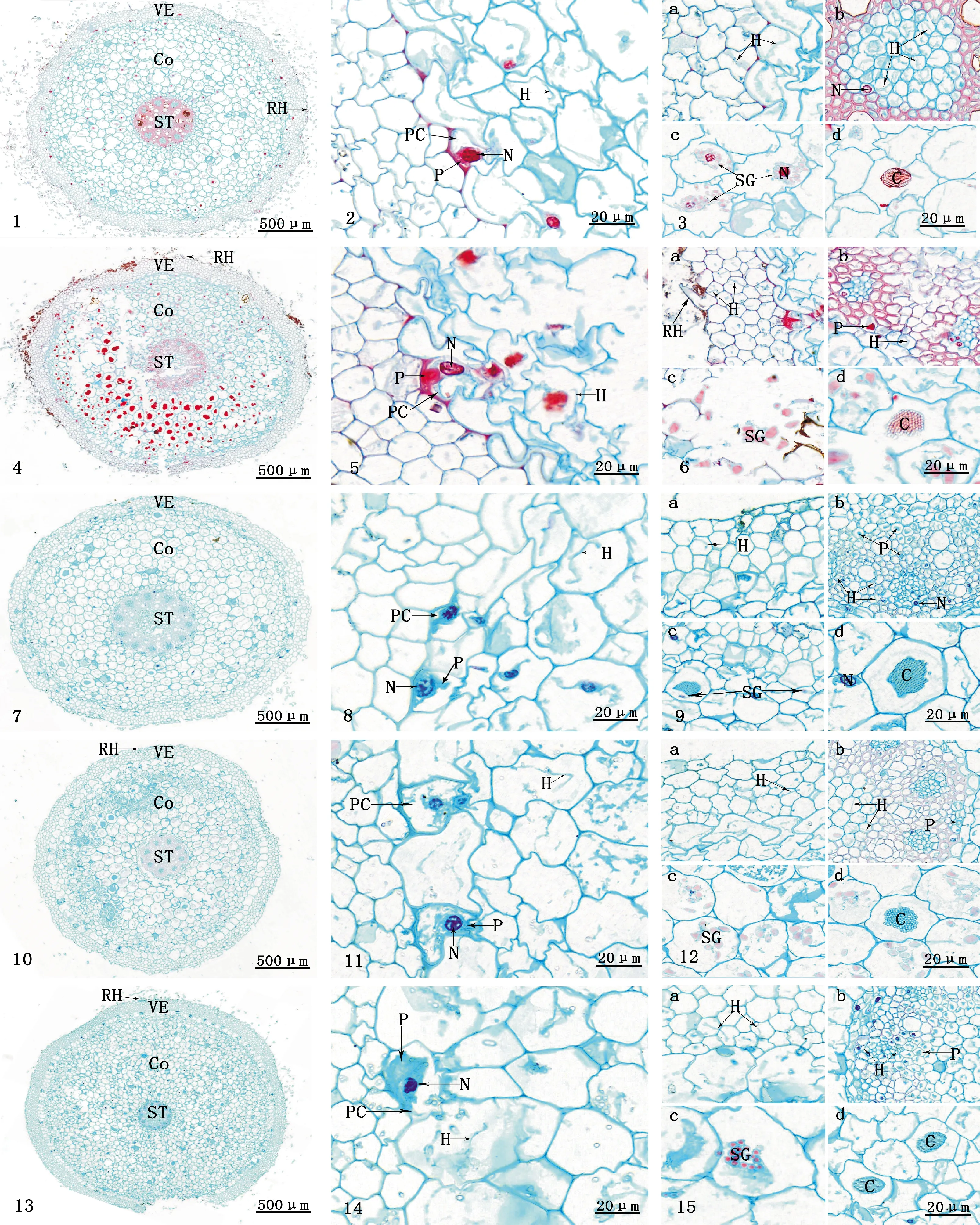
圖版Ⅰ5種蝦脊蘭菌根的顯微結構 1-3.劍葉蝦脊蘭;4-6.澤瀉蝦脊蘭;7-9.異鉤距蝦脊蘭;10-12.中華蝦脊蘭;13-15.銀帶蝦脊蘭;其中,圖1、4、7、10、13分別為5種蝦脊蘭菌根的橫切面,示根被、皮層、中柱和根毛;圖2、5、8、11、14示菌絲、菌絲結與外皮層通道細胞,菌絲在細胞核附近被消解,細胞核變形膨大;圖3、6、9、12、15中的a、b、c、d分別示根被細胞中的菌絲、中柱內的菌絲與菌絲結、淀粉粒、針狀結晶體;VE.根被;Co.皮層;ST.中柱;RH.根毛;H.菌絲;P.菌絲結;N.細胞核,PC.通道細胞;C.針狀結晶體;SG.淀粉粒
PlateⅠ Mycorrhizal microstructures of five Species ofCalanthe
Fig.1-3.Calanthedavidii;Fig.4-6.Calanthealismaefolia;Fig.7-9.Calanthegracilifloraf.jiangxiensis;Fig.10-12.Calanthesinica;Fig.13-15.Calantheargenteostriata;Fig.1,4,7,10 and 13 are the transverse section of mycorrhiza of five species ofCalanthe, showing the velamen, cortex, stele and root hair;Fig.2,5,8,11 and 14 showing the hyphae, pelotons and passage cells in the exodermis. The hyphae is digested near the nucleus and the nucleus is deformed and expanded;Fig.a,b,c and d in 3,6,9,12 and 15 show the hyphae in the velamen cells, respectively. The hyphae and pelotons in the stele, starch grain and crystals; VE. Velamen; Co. Cortex; ST. Stele; RH. Root hair; H. Hyphae; P. Pelotons; N. Nucleus; PC. Passage cell; C. Crystals; SG. Starch grain
