冰川退縮跡地植被原生演替初期關鍵種的碳氮磷生態化學計量特征
2021-06-25 02:20:02莊文化李明明史忠林
水土保持通報 2021年2期
關鍵詞:植物
周 萍, 莊文化, 李明明, 龍 翼, 史忠林, 周 繼
(1.中國科學院、水利部 成都山地災害與環境研究所, 山地表生過程與生態調控重點實驗室, 四川 成都 610041; 2.四川大學 水利水電學院, 四川 成都 610065; 3.四川大學水力學與山區河流開發保護國家重點實驗室, 四川 成都 610065; 4.中國科學院大學, 北京 100049)
生態化學計量學是研究生態系統營養元素分配、循環過程和限制性元素判斷的一種新興手段[1],主要用于研究不同生態系統、不同組織水平、不同有機體內部能量平衡、養分循環和化學元素(主要是C,N,P)的動態平衡[1]。生態化學計量特征不僅反映了養分狀況,同時還能揭示養分的可獲得性及其限制因子[2],這對探討植物養分循環與分配的調控機制、揭示元素平衡、分析生態要素交互作用具有重要意義[1]。高山或極地冰川消退區所形成的冰川裸地作為研究植被原生演替的理想場所,引起了生態學家的廣泛關注[3]。貢嘎山海螺溝地區第四紀冰川廣泛發育,下伸到谷地,現代冰川相當發育[4]。松散固體物質儲量巨大,受全球、局地氣候變化(溫度升高,暴雨增多或減少)、地震等,冰川末端逐漸退縮,冰川退縮后形成了大量原生裸地。由于此處受到的人為干擾少,隨著時間的推移,在原來冰川退縮跡地上重新開始了植物的原生演替,演替序列具有連續性和完整性,有利于研究冰川退縮區植被演替動態[5]。冰川退縮形成的原生裸地在第4 a就有被子植物生長發育,在第5—6 a的冰川退縮跡地就會形成以柳葉菜(Epilobiumhirsutum)和馬河山黃芪(Astragalusmahoschanicus)為關鍵種的群落,最初群落的植物比較稀疏,隨著馬河山黃芪固氮作用的加強,改善了局部生境,在第6—8 a,馬河山黃芪取代柳葉菜成為建群種,再經過3~8 a,沙棘(Hippophaerhamnoides)取代馬河山黃芪成為建群種,冰川退縮跡地朝著草本向灌叢的演替進行。植物對生境的適應是其存活和定居的關鍵,碳氮磷元素是植物生存與生長發育必需的結構元素[6]。植物不同器官的碳氮磷及其生態化學計量特征的研究有助于解釋植物碳氮磷元素在不同器官內的分配規律及植物為適應環境對自身養分的調控能力[7]。近年來,國內外學者對生態系統的化學計量特征研究主要集中在沙地[8]、濕地[9]、荒漠等[10]不同生態系統,但涉及冰川退縮跡地植物C,N,P生態化學計量及植物各器官碳氮磷化學計量耦合等的研究較缺乏[11]。為此,本研究基于對貢嘎山海螺溝冰川退縮跡地植被演替序列初期的兩種關鍵種的調查,選取了演替初期的柳葉菜和馬河山黃芪2個關鍵種,分別代表冰川退縮跡地植被演替初期第5 a和第8 a的演替群落,探討了演替初期植被不同器官的C,N,P分配特征及化學計量比的變化。并從生態化學計量特征的角度分析不同器官間C,N,P之間的相關關系,這對探索冰川退縮跡地植被演替初期關鍵物種的個體生長、種群動態和限制元素的判斷,預測指示植被的演替進程及從一定程度上揭示演替早期關鍵種的侵入和維持機制提供科學依據和數據支持。
1 研究區概況與研究方法
1.1 研究區概況
海螺溝是貢嘎山東坡最主要冰川河,也是我國最具代表性的季風海洋性山谷冰川之一,水熱條件好,冰川消融速度快,自小冰期開始退縮[12],近百年來沒有冰進過程,土壤有連續成土過程。20世紀30年代退縮加速形成了貢嘎山東坡海螺溝冰川退縮區(29°3 421″N,102°5 942″E),在長達2 km的范圍內形成了一個完整的從裸地到先鋒群落再到頂級群落的連續植被原生演替序列。該區主要受東亞季風影響,該地區氣候濕冷,屬于山地寒溫帶氣候類型,年平均氣溫3.8 ℃,1月平均氣溫最低(-4.38 ℃),7月平均氣溫最高(11.9 ℃),年降水日數在260 d以上,年平均降水量約1 960 mm,年平均相對濕度約90%。
1.2 研究方法
1.2.1 樣品采集 本研究以120多年形成的貢嘎山海螺溝冰川退縮區的末端作為研究區,基于前人對海螺溝冰川退縮區植被演替不同階段的劃分[11],采用以空間代時間的方法,選取了冰川退縮跡地植被原生演替第5年的柳葉菜群落和第8年的馬河山黃芪群落2個典型樣地為主要研究區域,于2019年6—10月在2個樣地分別隨機布設5個1 m×1 m樣方,調查樣方內所有草本和地被物,記錄其種類、數量等,并采集植物根系、莖、葉片、果實樣品帶回實驗室進行預處理。
1.2.2 植物不同器官CNP測定 樣方內選取生長健壯、長勢良好的柳葉菜和馬河山黃芪植株,從上中下3個層次分別采集每株植物成熟且無病蟲害葉片、莖樣品混勻后裝入信封中。在成熟季節分別采集柳葉菜和馬河山黃芪的果實樣品,并挖取植株完整根系沖洗后裝入信封,做好標記并帶回實驗室。將帶回實驗室的植物樣品置于恒溫干燥箱中在105 ℃下殺青0.5 h,然后在80 ℃下烘干到質量恒定,再粉碎并過100目(孔徑為0.15 mm)篩后保存用于CNP測定。葉片、莖、果實、根系有機碳采用重鉻酸鉀外加熱法測定,TN采用凱氏定氮法測定,TP采用高氯酸—硫酸消解鉬銻抗比色法測定[13]。
1.2.3 數據處理 使用SPSS 18.0軟件對數據進行統計分析,采用單因素方差分析法(one-way ANOVA)分析冰川退縮跡地演替初期柳葉菜和馬河山黃芪不同器官的C,N,P含量以及元素生態化學計量比(C/N,C /P,N/P)特征,并運用Excel作圖。
2 結果與分析
2.1 冰川退縮跡地植被演替初期兩種關鍵種不同器官碳氮磷含量特征
碳(C)、氮(N)、磷(P)作為組成植物體的基本元素,在植物生長發育及其各項生理活動協調中發揮著重要作用。植物不同器官的碳氮磷存在相關關系,C是植物體干物質的組成元素[14],N是蛋白質的重要組成部分[15],P與細胞結構和DNA,RNA的組成密切相關[1]。冰川退縮跡地演替初期兩種關鍵種柳葉菜和馬河山黃芪不同器官C,N,P含量可反映出該植物對其生長環境的適應特征。柳葉菜根系有機碳含量變化范圍為436~445 g/kg,較馬河山黃芪根系有機碳含量(416~423 g/kg)高(圖1)。葉片的C含量主要來源于土壤、根系和地上部分器官的輸移能力[16],冰川退縮跡地演替初期,退縮跡地土壤養分貧瘠,適宜物種較少,每年的地表生物量及枯落物歸還土壤的量均較少。柳葉菜和馬河山黃芪根系全氮含量表現出與有機碳相反趨勢,由于馬河山黃芪為豆科固氮植物,促進了根系和地上部分對N素的吸收利用[16],因此馬河山黃芪根系全氮含量顯著高于柳葉菜根系全氮含量(p<0.05)。柳葉菜和馬河山黃芪根系全磷含量較低,且未達到顯著性水平,這主要是由于土壤中的P是一種沉積性元素,受成土母質、氣候等影響,主要來源于巖石風化[17],冰川退縮跡地演替初期巖石風化程度低。柳葉菜和馬河山黃芪莖的有機碳含量表現為:柳葉菜>馬河山黃芪,且差異顯著(p<0.05)。而莖全氮和全磷含量均表現為:柳葉菜<馬河山黃芪,且差異顯著(p<0.05)。葉片是植物代謝最活躍的器官,它的養分特征對環境響應的敏感性較高。柳葉菜和馬河山黃芪葉片有機碳含量(311~394 g/kg)和全磷含量(1.49~1.78 g/kg)間的差異均未達到顯著水平。柳葉菜葉片全氮含量顯著低于馬河山黃芪葉片的全氮含量(p<0.05)。柳葉菜和馬河山黃芪果實的有機碳含量顯著高于馬河山黃芪果實的有機碳含量(p<0.05),果實的全氮含量則表現為柳葉菜<馬河山黃芪,且達到顯著性差異(p<0.05)。兩個關鍵種果實的全磷差異未達到顯著性水平。
柳葉菜不同器官的有機碳含量變化為:根系>果實>葉片>莖,柳葉菜葉片的全氮含量最高,其次為根系、果實和莖,柳葉菜全磷含量表現為:果實>葉片>根系>莖。馬河山黃芪不同器官的有機碳含量變化為:黃芪根系>果實>莖>葉片。馬河山黃芪全氮含量最高的器官為葉片,其次為根系、果實和莖,全磷含量變化為:葉片>果實>根系>莖。柳葉菜和馬河山黃芪不同器官的C,N,P平均值均低于全球平均水平和我國平均水平[18],但2個關鍵種不同器官的CNP分配策略相似,均優先保障果實和葉片的NPK含量以繁衍后代和維持自身生長代謝,不斷推動冰川退縮跡地植被演替的進行。

注:不同小寫字母表示在0.05水平差異顯著。下同。
2.2 冰川退縮跡地植被演替初期兩種關鍵種不同器官生態化學計量比特征
冰川退縮跡地植被演替初期的柳葉菜和馬河山黃芪兩種關鍵種的根系C/N,葉片C/N和果實C/N之間存在顯著差異(p<0.05),2種關鍵種的莖C/N之間差異不顯著(圖2)。柳葉菜不同器官的C/N高于馬河山黃芪,C/N高說明植物固碳效率較高,柳葉菜較馬河山黃芪處于演替的更早期,因此柳葉菜不同器官對碳的固定效率提高。柳葉菜和馬河山黃芪兩種關鍵種不同器官的N/P變化范圍為5.94~17.82,也表現出根系N/P,葉片N/P和果實N/P之間存在顯著差異(p<0.05),柳葉菜和馬河山黃芪的莖N/P之間差異不顯著。C/P可表征植物磷素的利用效率與碳同化能力,柳葉菜和馬河山黃芪根系C/P和莖C/P之間差異不顯著,而兩種關鍵種的葉片和果實C/P比存在顯著差異(p<0.05)。
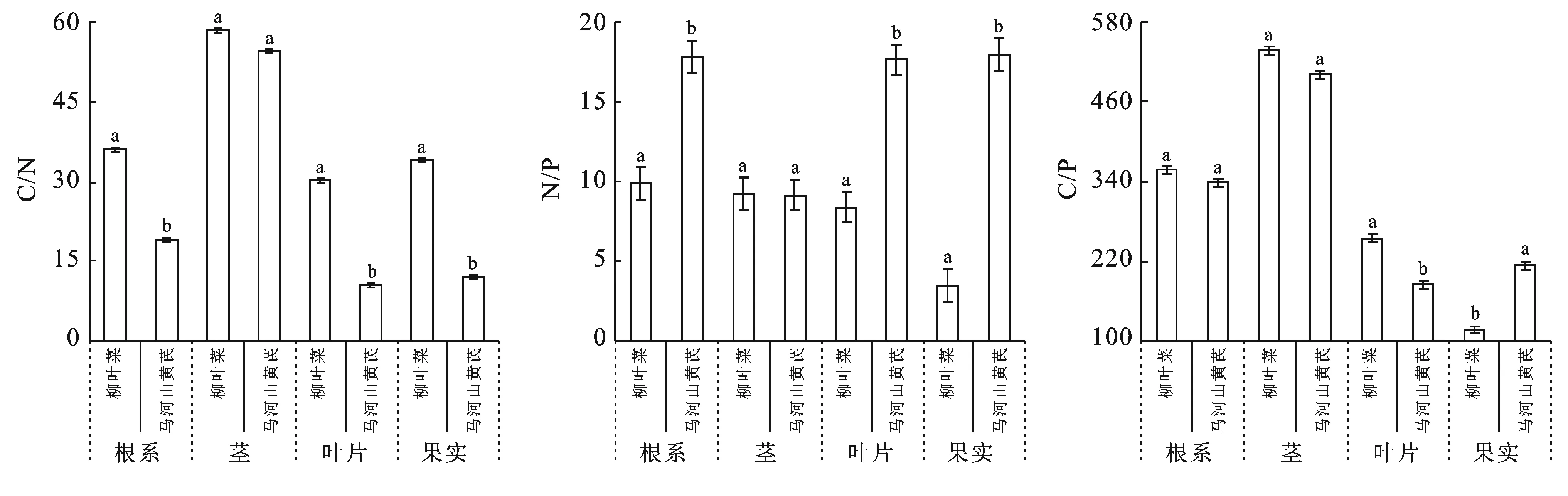
圖2 柳葉菜和馬合山黃芪不同器官生態化學計量比特征
2.3 冰川退縮跡地植被演替初期兩種關鍵種不同器官碳氮磷及生態化學計量比間相關性分析
冰川退縮跡地植被演替初期柳葉菜根系C含量與莖N含量、果實C含量之間呈現極顯著正相關(p<0.01)(表1),柳葉菜根系P含量與根系C含量、葉片P含量及果實P含量間顯著正相關(p<0.05),柳葉菜莖C含量與葉片P含量間極顯著正相關(p<0.01),柳葉菜莖N含量與果實C含量間極顯著負相關(p<0.01),柳葉菜葉片C含量與葉片N含量間極顯著正相關(p<0.01)。

表1 柳葉菜不同器官C,N,P間相關性
冰川退縮跡地植被演替初期柳葉菜根系C/N與莖C/N、葉片C/N間存在顯著正相關關系(p<0.05)(表2),柳葉菜根系N/P與葉片N/P間,根系C/P與莖C/P間存在顯著負相關關系(p<0.05),根系C/P與葉片C/P間存在極顯著正相關關系(p<0.01),根系C/P與果實N/P間顯著正相關(p<0.05)。柳葉菜莖N/P與莖C/P間極顯著正相關(p<0.01),柳葉菜莖N/P與葉C/P,莖N/P與果實N/P間顯著負相關(p<0.05)。柳葉菜莖C/P與葉片C/P間顯著負相關,莖C/P與果實N/P間極顯著負相關(p<0.01)。柳葉菜果實N/P與葉片N/P,果實N/P與果實C/P間存在顯著正相關關系(p<0.01)。

表2 柳葉菜不同器官生態化學計量比間相關性
冰川退縮跡地植被演替初期馬河山黃芪不同器官的C,N,P含量相關性分析結果表明(表3),馬河山黃芪不同器官的C,N,P含量間相關性較低,馬河山黃芪莖P含量與葉片P含量間存在極顯著正相關關系(p<0.01),葉片P含量與果實N含量間存在顯著正相關關系(p<0.05)。

表3 馬河山黃芪不同器官C,N,P間相關性
冰川退縮跡地植被演替初期馬河山黃芪根系C/N與葉片C/P間存在顯著正相關關系(p<0.05)(表4),莖N/P與莖C/P間顯著負相關(p<0.05),莖N/P與莖C/N間極顯著正相關(p<0.01),莖N/P與葉N/P間存在顯著正相關關系(p<0.05)。馬河山黃芪莖C/P與莖C/N,莖C/P與果實C/N間存在顯著負相關關系(p<0.05),葉片C/N與葉片N/P間存在顯著正相關關系(p<0.05)。
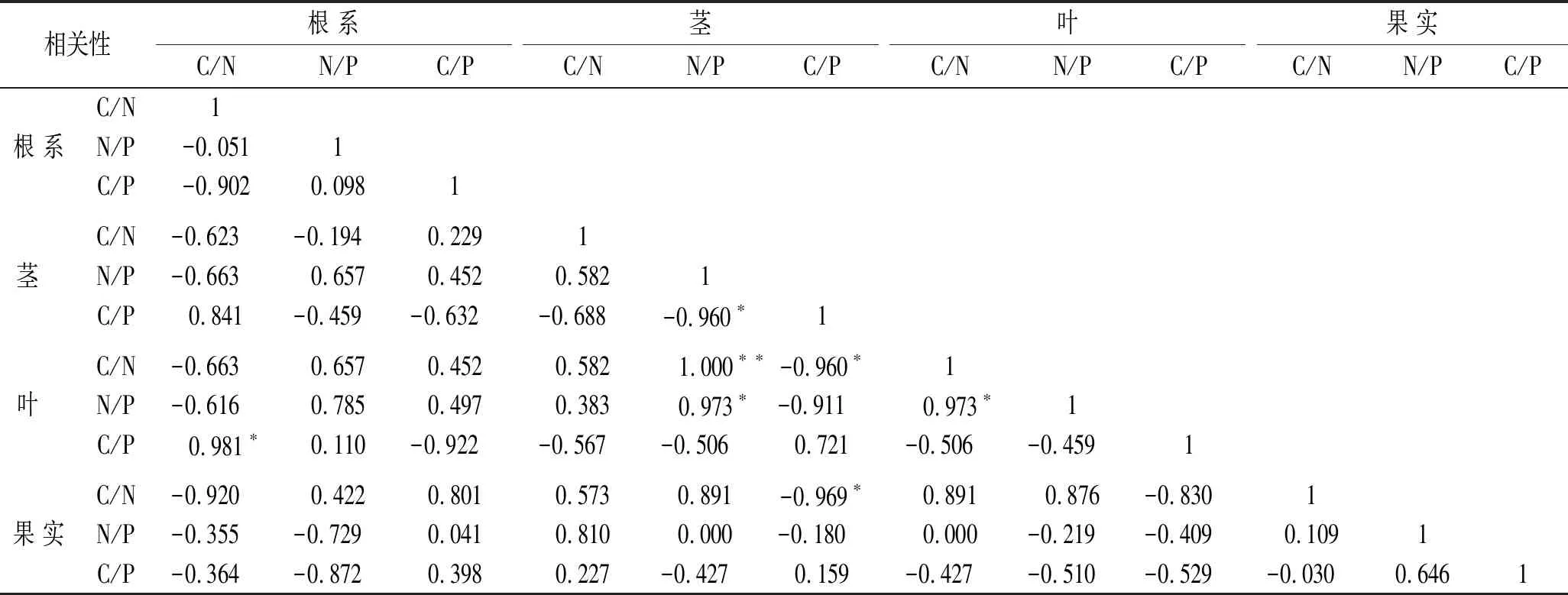
表4 馬河山黃芪不同器官生態化學計量比間相關性分析
3 討 論
3.1 冰川退縮跡地植被演替初期關鍵種的C,N,P元素特征
海螺溝冰川屬于季風海洋性冰川,與歐洲、北美等處于高緯度的冰川相比,海螺溝冰川退縮區具有氣溫高、降水量大的特點,水熱條件較好,為早期先鋒植物的定居提供了良好的環境基礎。Chapin等[19]通過對阿拉斯加Glacier Bay冰川原生演替研究發現,先鋒植物在冰川退縮第10 a后才開始侵入原生裸地,且最初形成的先鋒群落沒有喬木幼苗成分,到15—20 a才有柳樹和赤楊(Alnusrubra)進入群落。而貢嘎山海螺溝冰川退縮跡地,在第4 a就有種子植物黃芪、柳葉菜等生長發育,隨著演替的進行有喬灌木幼苗伴生其中,植被演替速度加快。
冰川退縮跡地植被演替初期,柳葉菜和馬河山黃芪常作為先鋒物種快速侵入,隨著演替的推進,馬河山黃芪的固氮作用不斷得到發揮,在演替到第6年,柳葉菜群落優勢地位逐漸被馬河山黃芪群落替代。植物不同器官的C,N,P含量及差異反映了植物對元素的吸收、運輸和利用特征[20]。葉片與根系是植物地上部分和地下部分重要的兩個營養器官,葉片是植物進行光合作用和制造有機物的重要器官[21],根系是植物體吸收、存儲、運輸養分水分[22]的主要器官。對比分析植物不同器官的C,N,P含量及其相關性有助于深入理解植物不同器官對C,N,P等營養物質的利用與分配規律[23]。冰川退縮跡地演替初期的柳葉菜和馬河山黃芪兩種關鍵種均為多年生草本植物,相比于一年生草本植物,多年生草本植物通常具有較長的葉壽命和較高的養分含量[24]。本研究的柳葉菜和馬河山黃芪不同器官C含量平均值(409.63 g/kg)均低于全球植物C平均值[25],表明該區域植物不同器官C含量較低。柳葉菜不同器官N含量值變化為7.48~12.98 g/kg,表現為:葉片>果實>根系>莖,低于我國[19]和全球[26]植物葉片N含量。這可能是由于柳葉菜處于冰川退縮跡地演替初期,退縮跡地土壤養分較貧瘠有關。柳葉菜不同器官P含量均值為0.81~3.52 g/kg,與我國和全球尺度植物葉片P含量的研究較為接近[19]。這可能是由于柳葉菜不同器官的P元素主要來源于退縮跡地的母巖風化,由于冰川退縮跡地成土母質風化速度慢,使得冰川退縮跡地表層土壤P含量相對較低且變幅小(表5)。且2個關鍵種不同器官的CNP分配策略相似,均優先保障果實和葉片的NPK含量以繁衍后代和維持自身生長代謝,不斷推動冰川退縮跡地植被演替的進行。植被演替年限稍長于柳葉菜的馬河山黃芪莖P含量與葉片P含量,葉片P含量與果實N含量間存在顯著正相關關系,這有助于深入理解不同植物的養分利用及其對環境變化的響應。

表5 冰川退縮跡地植被演替初期柳葉菜和馬河山黃芪不同器官N,P含量及N/P與其他研究結果比較
3.2 冰川退縮跡地植被演替初期關鍵種不同器官的化學計量特征
生態化學計量學是基于元素動態平衡原理和生長速率理論,可反映植物營養狀況、養分需求、養分利用效率、內穩態特征以及養分限制等[1]。不同植物化學計量特征及養分分配策略存在差異,能夠在一定程度上反映植物的適應能力[27]。C/N和C/P代表植物吸收養分且同化C的能力,即植物對養分的吸收利用率及固碳效率,N/P代表對生產力起限制作用的指示劑,能影響植物群落的結構和功能[28]。生態化學計量比能有有效反應植物養分循環與限制作用。可利用C/N,N/P,C/P生態化學計量分析揭示生態系統中養分的可獲得性以及C,N,P元素的循環、平衡機制與相互制約關系。N,P作為陸地生態系統中植物生長的主要限制元素,N/P可反映生態系統中植物受N,P養分的供應與限制特征[29]。當植被葉片N/P<14時,主要是受到N的限制,當N/P>16時,主要是受到了P的限制,當14 冰川退縮跡地演替初期柳葉菜和馬河山黃芪兩種關鍵種不同器官C,N,P含量及其化學計量比存在相關關系。C,N表現為極顯著正相關,說明地上器官葉片與地下器官細根養分分配具有一致性,功能聯系密切。柳葉菜根系、莖、葉片C/N間表現為相互促進、協同增長的正相關關系,柳葉菜果實N/P與葉片N/P間存在顯著正相關關系(p<0.01)。馬河山黃芪莖P含量與葉片P含量間存在極顯著正相關關系(p<0.01),葉片P含量與果實N含量間存在顯著正相關關系(p<0.05),根系C/N與葉片C/P間,莖N/P與莖C/N間,莖N/P與葉N/P間,葉片C/N與葉片N/P間存在極顯著正相關關系(p<0.01),馬河山黃芪莖N/P與莖C/P間存在顯著負相關關系(p<0.05),莖C/P與莖C/N,與果實C/N間存在顯著負相關關系(p<0.05)。說明植物不同器官在進行C,N,P分配過程中,能量如果更多地用于生長,則會減少同化產物貯存。 (1) 柳葉菜根系有機碳含量較馬河山黃芪根系有機碳含量高。馬河山黃芪為豆科固氮植物,其根系全氮含量顯著高于柳葉菜根系全氮含量。柳葉菜不同器官的全磷含量較馬河山黃芪不同器官的全磷含量高。 (2) 柳葉菜不同器官的C,N,P含量均低于全球平均水平和我國平均水平,馬河山黃芪不同器官的N含量略高于全球和我國平均水平,C,P含量均低于全球平均水平,2種關鍵種的各養分分配策略相似,均優先保障果實和葉片的N,P,K含量以繁衍后代和維持自身生長代謝推動冰川退縮跡地植被演替的進行。 (3) 冰川退縮跡地植被演替初期的柳葉菜不同器官的C/N高于馬河山黃芪,高C/N值反映出柳葉菜具有較高的碳同化效率、較慢的生長速率及較低的營養需求,即資源保護策略。馬河山黃芪高N/P和低C/P值反映了其具有高生長率或繁殖輸出,即生長競爭策略,與柳葉菜相比,馬河山黃芪具有快速生長的特性,以生長競爭優勢即生長競爭策略戰勝柳葉菜,從而促進冰川退縮跡地植被原生演替的向前推進。3.3 冰川退縮跡地植被演替初期關鍵種不同器官化學計量比的相關關系
4 結 論
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
