高世代回交玉米矮稈種質的轉錄組分析
2021-06-30 02:23:53王亞麗陳煜東王益軍
江蘇農業學報 2021年2期
王亞麗 陳煜東 王益軍
摘要:?株高影響株型建成,與抗倒性、收獲指數以及最終產量相關。對矮稈材料的研究有助于豐富對株高調控機理的認識。在前期研究中,對玉米矮稈材料D11的農藝與生理特征進行了分析。本研究基于高世代回交群體,通過轉錄組測序,對矮稈材料D11的基因表達調控進行解析。與株高正常植株相比,矮稈材料中共檢測到2 537個差異表達的基因,其中1 120個基因表達上調、1 417個基因表達下調。功能注釋、GO富集、通路富集的結果表明,差異表達基因參與細胞延伸、細胞骨架功能、微管組織、葉綠體功能、植物激素合成代謝等。差異表達基因最顯著富集的通路與糖酵解相關。共表達分析結果表明,編碼果糖-1,6-雙磷酸酶的基因與多個差異表達基因存在功能關聯。該研究結果揭示了玉米矮稈材料中的基因表達調控,為后續玉米株高調控節點的發掘提供了參考。
關鍵詞:?矮稈;高世代回交群體;轉錄組測序;共表達;玉米
中圖分類號:?S513.035.3??文獻標識碼:?A??文章編號:?1000-4440(2021)02-0280-09
Abstract:?Plant height affects plant type and is related to lodging resistance, harvest index and final yield. The research on dwarf plants helps to improve the understanding of plant height regulation mechanism. In previous studies, the agronomic and physiological characteristics of maize dwarf material D11 were analyzed. In this study, based on the advanced backcross population, the gene expression regulation of D11 was analyzed by transcriptome sequencing. A total of 2 537 differentially expressed genes (DEGs) were identified,including 1 120 up-regulated and 1 417 down-regulated DEGs. Functional annotation, GO enrichment and pathway enrichment analysis results indicated that DEGs were involved in cell expansion, cytoskeletal function, microtubule organization, chloroplast function, phytohormone homeostasis. The pathway most significantly enriched by DEGs was related to glycolysis. The gene encoding fructose-1,6-bisphosphatase was functionally related to several DEGs. The results of this study reveal the gene expression regulation in maize dwarf materials, and provide a reference for the subsequent exploration of maize plant height regulatory modes.
Key words:?dwarf plant;advanced backcross population;transcriptome sequencing;co-expression;maize
株高是重要的農藝性狀,影響抗倒性、光合效率、收獲指數和最終產量。“綠色革命”期間,產量的提高主要得益于株型與農藝措施的改良。改良的高產品種具有半矮稈性狀、較高的氮肥同化效率、優良的抗倒性[1]。控制高產品種半矮稈性狀的基因已被克隆。水稻的半矮稈基因semidwarf1 (sd1)編碼負責赤霉素合成的酶GA 20-oxidase2 (GA20ox2)[2-3]。小麥株高降低是由于Reduced height (Rht)基因的變異所導致,Rht編碼DELLA蛋白,參與赤霉素信號轉導[4]。“綠色革命”期間,半矮稈性狀主要是由于赤霉素合成、信號轉導通路的改變所導致。除了赤霉素,其他植物激素,例如生長素、油菜素內酯、獨腳金內酯等植物激素也與株高調控相關。生長素轉運異常導致玉米和高粱植株矮化[5]。玉米嚴重矮化表型是由于編碼油菜素內酯合成酶brassinosteroid C-6 oxidase基因的突變所導致[6]。油菜素內酯信號通路的改變同樣能導致玉米植株矮化[7]。植物激素獨腳金內酯誘導豌豆幼莖伸長[8]。除了植物激素,表觀調控因子也影響株高。通過全基因組關聯分析,調控玉米株高的表觀因子被發掘[9]。
玉米株高是由微效多基因控制的性狀[10]。通過連鎖分析、全基因組關聯分析,已有大量調控玉米株高的QTL/靶點被發掘[10-11]。其中,一些玉米株高QTL已被圖位克隆,例如qPH3.1、qph1[12-13]。QTL克隆是發掘玉米株高調控靶點的重要方法。通過玉米矮稈突變體研究,亦能為株高調控機制的解析提供重要參考。在前期研究中,實驗室鑒定到1份玉米矮稈材料Dwarf11 (D11),矮稈材料D11的農藝性狀、生理特征已被初步分析[14]。基于高通量測序的轉錄組技術,被用來研究植物調控網絡。轉錄組技術可用于SNP發掘、可變剪輯預測、融合轉錄本鑒定、基因表達分析等[15]。本研究采用轉錄組測序技術,結合熒光定量PCR驗證、基因功能注釋、通路富集分析、共表達網絡構建等,對高世代回交D11矮稈群體的基因表達調控進行了分析,所得結果有助于解析矮稈基因D11介導的調控網絡,也為后續玉米株高調控靶點的解析提供參考。
1?材料與方法
1.1?試驗材料
矮稈材料D11與玉米自交系Mo17雜交,獲得F1代。通過連續5個世代的回交,獲得高世代回交群體BC5。在高世代回交群體BC5中,與株高正常植株相比,矮稈材料株高顯著降低。本研究采用高世代回交群體BC5中的株高正常植株與矮稈植株作為試驗材料。
1.2?RNA提取、文庫構建與高通量測序
在高世代回交群體BC5中,取三葉期幼莖,采用RNeasy Plant Mini Kit (QIAGEN)試劑盒進行RNA提取。提取RNA的質量與濃度用NanoDrop 2000 spectrophotometer、Agilent 2100 Bioanalyzer進行測定。mRNA通過帶有oligo(dT)的磁珠進行吸附。吸附的mRNA片段,反轉錄成cDNA。cDNA末端修復,加腺嘌呤,連接接頭。連接產物進一步純化,用于PCR擴增。回收400~500 bp大小的產物,采用RNA Seq Library Preparation Kit進行文庫構建。測序前,文庫質量采用ABI StepOnePlus Real-Time PCR System進行qPCR檢測。合格的文庫基于125 bp成對模式,在Illumina Hiseq2500 platform進行高通量測序。
1.3?差異表達基因發掘
對轉錄組測序獲得的原始序列,首先進行質量控制。接頭序列、低質量reads(含有超過50%低質量堿基的reads、未知堿基數超過5%的reads、測序質量低于10的低質量堿基)被剔除。采用SOAPaligner/soap2,過濾后的reads比對到玉米參考基因組[16]。比對結果質量控制后,采用Cufflinks進行轉錄本組裝[17]。通過計算每1×106 reads中來自于某基因每1 000堿基長度的reads數 (RPKM),對基因表達量進行度量。本研究中,表達變化≥2、發現錯誤率(FDR)≤0.05的基因為差異表達基因。
1.4?差異表達基因生物信息學分析
差異表達基因功能注釋通過BLAST 進行。玉米基因功能注釋使用maizeGDB數據庫 (https://www.maizegdb.org/)。水稻、擬南芥中差異表達基因的同源基因通過BLASTP獲得,所用數據庫分別為RGAP (http://rice.plantbiology.msu.edu/index.shtml)與Araport (https://www.araport.org/)。Gene ontology (GO)富集分析:首先基于Gene Ontology Consortium (http://www.geneontology.org/) 數據庫,將差異表達基因比對到GO term[18]。確定每個GO term的差異表達基因數,超幾何分布檢驗用來發掘顯著富集的GO term。通過Bonferroni校正,獲得校正P值。校正P值小于0.05為差異表達基因顯著富集的GO term。通路富集分析采用KEGG 數據庫(http://www.genome.jp/kegg/)進行[19]。校正P值小于0.05為差異表達基因顯著富集的通路。
1.5?熒光定量PCR分析
RNA樣品用RNase-free DNase I (QIAGEN) 處理,采用M-MLV reverse transcriptase (Promega)反轉錄,獲得cDNA。采用SYBR qPCR master mix (TaKaRa),在7500 Real-Time PCR System (ABI)上進行熒光定量PCR。每個樣品重復3次。18S rRNA基因作為內參基因。用2-△△Ct方法分析基因的相對表達量[20]。
2?結果與分析
2.1?高世代群體矮稈材料表型及基因表達分析
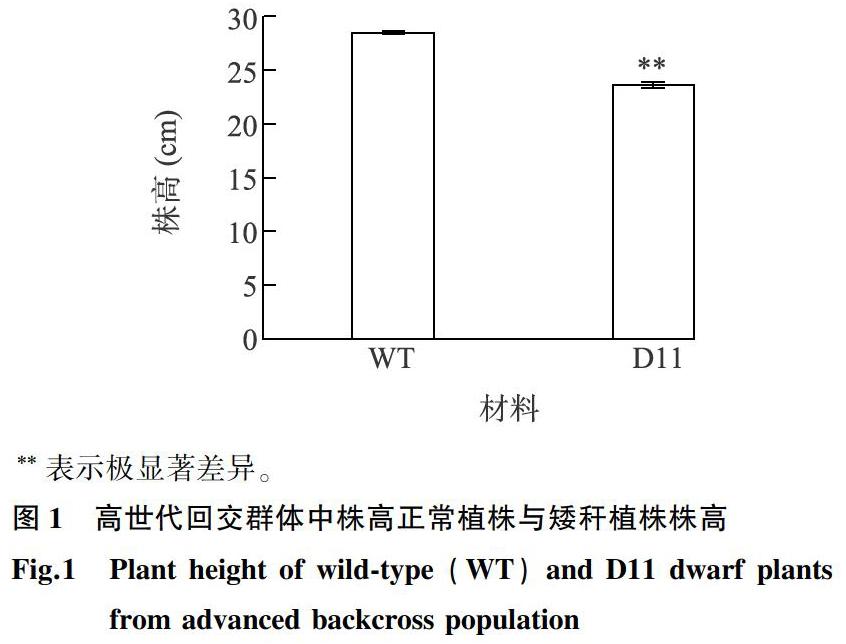
玉米矮稈材料D11與自交系Mo17雜交。通過連續5個世代的回交,獲得高世代回交群體BC5。在高世代回交群體BC5中,與株高正常植株相比,矮稈材料株高極顯著降低(圖1)。
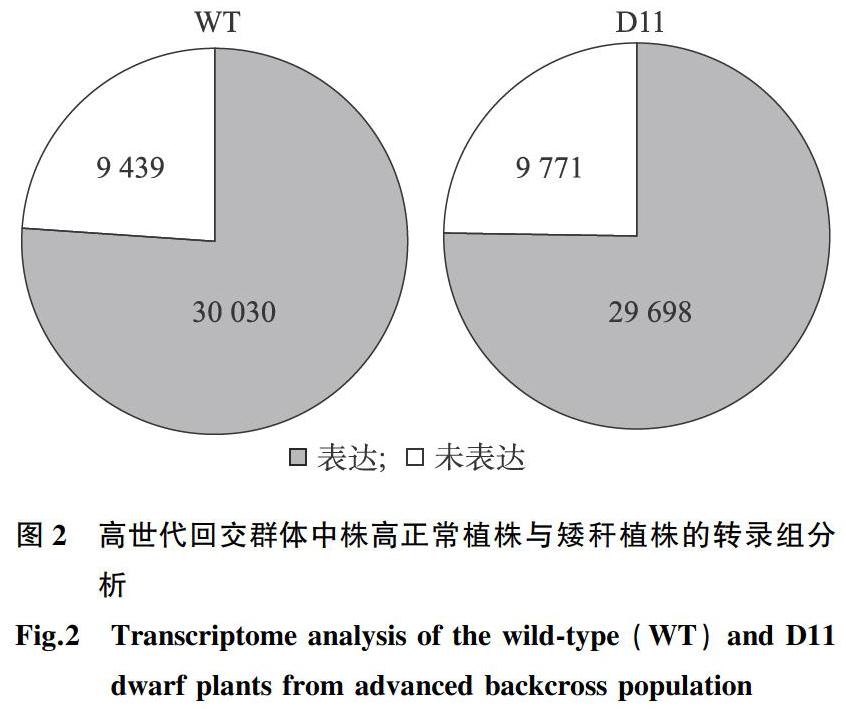
為了發掘矮稈材料中差異表達基因,取高世代回交群體中株高正常植株與矮稈植株,進行轉錄組測序。通過對原始測序數據質量控制、過濾,Clean reads比對到玉米參考基因組,對株高正常植株與矮稈植株中的基因表達進行分析。結果表明,在株高正常植株中檢測到超過30 000個基因的表達。在矮稈植株中檢測到近30 000個基因的表達(圖2)。
2.2?轉錄組測序發掘差異表達基因
為了發掘矮稈材料中差異表達基因,比對的reads通過計算RPKM值對基因的表達量進行度量。通過比較株高正常植株和矮稈植株的基因表達量,進行差異表達基因發掘。本研究中,基因表達變化≥2、發現錯誤率≤0.05的基因被認為是差異表達基因。根據上述設定的閾值,共發掘到2 537個差異表達基因。與株高正常植株相比,其中1 120個基因在矮稈材料中表達量上升、1 417個基因在矮稈材料中表達量下降。
2.3?轉錄組測序揭示的差異表達基因的驗證
基于轉錄組測序揭示的差異表達基因,設計引物(表1),進行熒光定量PCR分析,對差異表達基因進行驗證。
采用2-△△Ct方法,對基因的表達量進行相對定量。差異表達基因熒光定量PCR與轉錄組測序表達量的決定系數(R2)為0.72(圖3)。結果表明,轉錄組測序獲得的差異表達基因可靠,由轉錄組測序獲得的差異表達基因可用于后續分析。
2.4?差異表達基因功能
對轉錄組測序揭示的差異表達基因,首先在maizeGDB數據庫中進行功能注釋。此外,發掘水稻、擬南芥中差異表達基因的同源基因。玉米數據庫中沒有功能注釋的差異表達基因,通過其水稻、擬南芥中的同源基因進行功能注釋。功能注釋的結果表明,差異表達基因參與多個生物學過程。基于矮稈材料表型,本研究集中分析可能與矮稈表型相關的差異表達基因。與株高正常植株相比,矮稈材料中與細胞伸長相關的ARF GTPase家族成員基因差異表達,與微管組織、細胞骨架、木質素合成、維管束組織發育相關的基因差異表達。矮稈材料葉片變寬、微卷,通過轉錄組測序,檢測到與葉片發育相關的差異表達基因。例如,生長調節因子、MYB轉錄因子等。此外,矮稈材料葉緣白化。在矮稈材料中,與葉綠素合成、葉綠體功能相關的基因差異表達,例如編碼參與葉綠素合成Mg-螯合酶基因、編碼葉綠素a/b結合蛋白的基因、調控葉綠素降解的NAC轉錄因子基因等。差異表達基因還參與生長素、油菜素內酯、細胞分裂素等植物激素的合成與信號轉導過程(表2)。
本研究所用矮稈材料對外源赤霉素施用敏感。轉錄組測序的結果表明,與株高正常植株相比,矮稈材料中與赤霉素合成代謝、信號通路相關的基因差異表達。與赤霉素合成相關的4個基因差異表達,包括上調的GA20ox5基因,下調的Dwarf1、dwarf3、KO1基因。與赤霉素代謝相關的2個GA2ox基因均下調表達。與赤霉素信號轉導相關的6個基因,包括赤霉素受體基因、F-box蛋白基因等,其中2個基因表達上調、4個基因表達下調。與赤霉素通路調控相關的5個基因,在矮稈材料中均下調表達(表3)。
2.5?差異表達基因GO與通路富集分析
基于Gene Ontology Consortium resource數據庫,對差異表達基因進行GO分析。設定了較嚴格的閾值(校正P值<0.05)用來發掘差異表達基因顯著富集的GO term。在生物學過程中,差異表達基因富集在碳水化合物合成代謝、光合作用等過程。在分子功能方面,差異表達基因最顯著富集的GO term與結構組分相關。為了進一步分析差異表達基因參與的生物學通路,采用KEGG數據庫,對差異表達基因進行KEGG通路分析。與GO富集分析類似,校正P值<0.05作為顯著富集KEGG通路篩選的標準。KEGG通路富集的結果表明,差異表達基因最顯著富集的通路與糖酵解相關。差異表達基因可能參與糖的合成與代謝。此外,差異表達基因顯著富集的通路與RNA轉運、氨基酸合成、光合作用等過程相關(表4)。
2.6?差異表達基因共表達分析
共表達分析有助于發掘基因調控網絡的關鍵節點,為后續基因調控網絡的深入解析提供參考。本研究對轉錄組測序揭示的差異表達基因進行了共表達分析。結果表明,Class I-like SAM-binding甲基轉移酶基因(ID: GRMZM2G064759)下調表達。在基因共表達網絡中,該基因與10個差異表達基因互作,包括DUF947結構域蛋白基因(ID: GRMZM2G051197)、寡肽轉運類蛋白基因(ID: GRMZM2G434935)等。果糖-1,6-雙磷酸酶編碼蛋白基因(ID: GRMZM2G306732)上調表達,是共表達調控網絡的關鍵調節因子。該基因與多個差異表達基因功能相關,包括編碼纖維素-1,7-雙磷酸酶的shbp1基因、編碼磷酸核酮糖激酶基因(ID: GRMZM2G026024)等。基因共表達分析的結果表明,轉錄組測序揭示的差異表達基因可能存在一定的功能關聯。
3?討論
本研究所用矮稈材料是田間自然突變所產生,表現為植株矮化。在前期研究中,對該矮稈材料的農藝、生理特征進行了分析。結果表明,外源赤霉素施用可以使該矮稈材料的株高變高,表明該矮稈材料屬于赤霉素敏感型矮稈種質[14]。本研究基于高世代回交種質,通過轉錄組測序,對矮稈材料的基因表達調控進行了解析。采用高世代回交種質,有利于降低遺傳背景對基因表達的影響,提高轉錄組測序結果的可靠性。在高世代回交群體中,與株高正常植株相比,矮稈材料中共檢測到2 537個基因差異表達,其中1 120個基因上調表達、1 417個基因下調表達。差異表達基因功能注釋、GO富集、通路富集的結果表明,差異表達基因參與多個生物學過程,包括細胞延伸、微管組成、木質素合成、葉綠素合成、葉綠體功能、植物激素合成與信號轉導等。供試矮稈材料的矮化可能與細胞延伸的缺陷相關。葉綠素合成、葉綠體功能的異常,可能與矮稈材料的葉緣白化相關。木質素合成的變化與矮稈材料莖稈質地的改變相關[14]。不同生物學過程存在一定的關聯性。例如,植物激素赤霉素、生長素、油菜素內酯、細胞分裂素、獨腳金內酯等互作,協同調控了多個生物學過程[21-25]。此外,已有證據表明,植物激素赤霉素影響葉綠體的產生[26]。本研究轉錄組測序結果表明,供試矮稈材料中存在著復雜的正向、反向、交叉基因調控網絡,進而影響矮稈材料的表型。
通過比較不同矮稈材料的轉錄組數據,可以發掘共有的、特異的差異表達基因,為矮稈基因介導的調控網絡的解析提供科學依據。通過檢索數據庫,未能獲得d1、d3、d5、br2、na2、bv1、D8、D9等玉米矮稈材料的轉錄組數據。通過將本研究獲得的轉錄組數據與數據庫中玉米矮稈材料的轉錄組數據進行比較分析,有助于發掘本研究矮稈材料中特異的差異表達基因,特異差異表達基因的發掘可以為供試矮稈材料基因調控網絡的解析提供重要參考。
參考文獻:
[1]?KHUSH G S. Green revolution: the way forward[J]. Nature Reviews Genetics,2001,2(10): 815-822.
[2]?MONNA L,KITAZAWA N,YOSHINO R, et al. Positional cloning of rice semidwarfing gene,sd-1: rice ‘green revolution gene encodes a mutant enzyme involved in gibberellin synthesis[J]. DNA Research,2002,9(1): 11-17.
[3]?SPIELMEYER W,ELLIS M H,CHANDLER P M. Semidwarf (sd-1), ‘green revolution rice,contains a defective gibberellin 20-oxidase gene[J]. Proceedings of the National Academy of Sciences of the United States of America,2002,99(13): 9043-9048.
[4]?PENG J,RICHARDS D E,HARTLEY N M,et al. ‘Green revolution genes encode mutant gibberellin response modulators[J]. Nature,1999,400(6741): 256-261.
[5]?MULTANI D S,BRIGGS S P,CHAMBERLIN M A,et al. Loss of an MDR transporter in compact stalks of maize br2 and sorghum dw3 mutants[J]. Science,2003,302(5642): 81-84.
[6]?MAKAREVITCH I,THOMPSON A,MUEHLBAUER G J,et al. Brd1 gene in maize encodes a brassinosteroid C-6 oxidase[J]. PLoS One,2012,7(1): e30798.
[7]?KIR G,YE H,NELISSEN H,et al. RNA interference knockdown of Brassinosteroid Insensitive1 in maize reveals novel functions for brassinosteroid signaling in controlling plant architecture[J]. Plant Physiology,2015,169(1): 826-839.
[8]?DE SAINT GERMAIN A,LIGEROT Y,DUN EA,et al. Strigolactones stimulate internode elongation independently of gibberellins[J]. Plant Physiology,2013,163(2): 1012-1025.
[9]?WENG J,XIE C,HAO Z,et al. Genome-wide association study identifies candidate genes that affect plant height in Chinese elite maize (Zea mays L.) inbred lines[J]. PLoS One,2011,6(12): e29229.
[10]PEIFFER J A,ROMAY M C,GORE M A,et al. The genetic architecture of maize height[J]. Genetics,2014,196(4): 1337-1356.
[11]李清超,李永祥,楊釗釗,等. 基于多重相關RIL群體的玉米株高和穗位高QTL定位[J]. 作物學報,2013,39(9): 1521-1529.
[12]TENG F,ZHAI L,LIU R,et al. ZmGA3ox2,a candidate gene for a major QTL,qPH3.1,for plant height in maize[J]. The Plant Journal,2013,73(3): 405-416.
[13]XING A,GAO Y,YE L,et al. A rare SNP mutation in Brachytic2 moderately reduces plant height and increases yield potential in maize[J]. Journal of Experimental Botany,2015,66(13): 3791-3802.
[14]WANG Y,LU W,CHEN Y,et al. Revealing physiological and genetic properties of a dominant maize dwarf Dwarf11 (D11) by integrative analysis[J]. Molecular Breeding,2016,36(3): 31.
[15]CONESA A,MADRIGAL P,TARAZONA S,et al. A survey of best practices for RNA-seq data analysis[J]. Genome Biology,2016,17: 13.
[16]LI R,YU C,LI Y,et al. SOAP2: an improved ultrafast tool for short read alignment[J]. Bioinformatics,2009,25(15): 1966-1967.
[17]ROBERTS A,PIMENTEL H,TRAPNELL C,et al. Identification of novel transcripts in annotated genomes using RNA-Seq[J]. Bioinformatics,2011,27(17): 2325-2329.
[18]GENE ONTOLOGY CONSORTIUM. Creating the gene ontology resource: design and implementation[J]. Genome Research,2001,11(8): 1425-1433.
[19]KANEHISA M,GOTO S,SATO Y,et al. KEGG for integration and interpretation of large-scale molecular data sets[J]. Nucleic Acids Research,2012,40(Database issue): D109-D114.
[20]LIVAK K J,SCHMITTGEN T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method[J]. Methods,2001,25(4): 402-408.
[21]王江英,朱朋波,湯雪燕,等. 外源赤霉素誘導矮生山茶恨天高植株生長的轉錄組分析[J].江蘇農業學報,2020,36(1):47-56.
[22]員盎然,孫小娟,馬小雯,等. GA3與ABA對黑穗醋栗二次萌芽總酚含量及代謝酶活性的影響[J].南方農業學報,2019,50(6):1263-1270.
[23]劉達濰,劉詩琴,易鵬程,等.淹水脅迫下外源激素對甜瓜幼苗根系生理特性的影響[J].江蘇農業科學,2019,47(9):182-185.
[24]曹?帥,杜仲陽,劉?鵬,等. 堿脅迫對大豆光合特性及內源激素含量的影響[J].江蘇農業學報,2020,36(2):284-291.
[25]VANSTRAELEN M,BENKOVá E. Hormonal interactions in the regulation of plant development[J]. Annual Review of Cell and Developmental Biology,2012,28: 463-487.
[26]JIANG X,LI H,WANG T,et al. Gibberellin indirectly promotes chloroplast biogenesis as a means to maintain the chloroplast population of expanded cells[J]. The Plant Journal,2012,72(5): 768-780.
(責任編輯:陳海霞)
