七種黃精屬植物的光合特性
2021-07-09 03:09:16馬英姿周詩雨
經濟林研究 2021年2期
關鍵詞:植物
劉 熙,宋 榮,馬英姿,周詩雨,劉 智
(1.中南林業科技大學 生命科學與技術學院,湖南 長沙 410004;2.湖南省農業環境生態研究所,湖南 長沙 410125)
黃精為百合科Liliaceae 黃精屬Polygonatum[1]。多年生草本植物的總稱,中國約有31 種,在國內分布較廣泛,該屬植物有肉質根狀莖,形似于圓柱形、結節處有時不規則,一般生長在灌叢、林下和陰涼處,具有保健、營養、滋補、治療腎虛、精血不足、肺虛、干咳等藥用功效,同時在抗衰老、調節自身免疫、降低機體血脂、增強機體記憶反應能力、抗惡性腫瘤、抗菌等方面也具有潛在的生物醫學價值[2-3]。目前,黃精種苗資源在很大程度上依賴于野生黃精種質資源,隨著現代社會對黃精的需求日益增加,我國黃精野生資源面臨枯竭和生態環境嚴重破壞的風險,林下栽培黃精可以減少黃精野生資源的消耗,并滿足市場對黃精的需求,同時,選育出優良的黃精種質資源已成為亟待解決的關鍵問題[4]。
農作物的產量取決于光合作用過程中光合產物的積累和分布,如何高效地利用太陽能增強植物的光合作用,目前仍然是現代農業發展中的一個根本性問題。選育高光效種質就是一種提高作物產量和品質的有效方法,選擇高光效種質的重要指標是其光合特性[5-6]。植物的光合特性主要包括光合作用日變化、光合速率、光飽和點和光補償點[7]。光合作用的日變化曲線可以直接反映各種植物之間的內在節律差異,還能夠反映各種植物之間環境適應性的差異,因此可以代表各種植物的生理特性和其它環境因素對它們的綜合影響[8-9]。本研究對湖南省內主要種植的多花黃精P.cyrtonema、黃精P.sibiricum、滇黃精P.kingianum、點花黃精P.punctatum、湖北黃精P.zanlanscianense、卷葉黃精P.cirrhifolium、長梗黃精P.filipes的光合特性進行了系統的測定,旨在對黃精屬植物的生理生態習性有更為精準的掌握,探討其對環境適應能力的差異,為選擇黃精屬植物的優良種質和科學高效地栽培利用提供理論依據。
1 材料與方法
1.1 試驗地與試驗材料
試驗地位于湖南省長沙市農業環境生態研究所基地。屬亞熱帶季風性濕潤氣候,年平均氣溫為17.5 ℃,年平均最高氣溫為21 ℃,年平均最低氣溫為14 ℃[10]。
本試驗材料均采自湖南省安化縣及湖南省農科院的長沙縣高橋基地3年生黃精屬植物栽培種,于2019年4月黃精出苗期整株采集移植。7 種黃精屬植物植株均經過湖南中醫藥大學王智老師鑒定,分別為點花黃精、多花黃精、滇黃精、湖北黃精、黃精、卷葉黃精和長梗黃精,栽培于林下環境中。
1.2 方法
光合指標測定時間為2020年7月中旬。測定時黃精屬植物均為4年生植株結實期,連續數日選擇晴朗天氣測定其光響應曲線、CO2響應曲線和日變化,每種黃精屬植物測定3 株,每株測定3片葉,選擇中上部位同一水平、成熟無病蟲害的葉片進行測定。
1.2.1 葉片葉綠素含量的測定
葉綠素含量用丙酮-乙醇混合法測定,丙酮∶乙醇(無水)=5∶5。取新鮮葉片0.1 g,切碎,放入試管中,加入25 mL 溶液,密封,在暗處放置6~8 h,然后在470、663、645 nm 進行比色,得到A值,根據Arnon法公式計算各提取物中葉綠素含量,重復測定3 次,計算其平均值[11-12]。
1.2.2 光合速率相關指標測定
以便攜式光合作用測試儀Li-6400 為測量儀器,于晴天測定日變化,共測量7 次,時間段分別 為7:00、9:00、11:00、13:00、15:00、17:00、18:00。每個種隨機選取3 株植物,每株選取3 片中上部位同一水平、成熟無病蟲害的葉片進行測定。測定7 種黃精屬植物葉片的凈光合速率(Pn)、胞間CO2濃度(Ci)、氣孔導度(Gs)、蒸騰速率(Tr)、光合有效輻射(PAR)、相對濕度(RH)和大氣溫度(Tair),水分利用效率(WUE)=Pn/Tr,對每片葉測定3 次,測定數據取平均值。
1.2.3 光響應曲線的測定
選取7 種黃精屬植物中間部位生長狀況較好的成熟葉片,在晴天用Li-6400 測定其光響應曲線。儀器葉片室采用紅、藍光光源,光響應曲線測量參數設定如下:參比室內CO2濃度約為400 μmol·mol-1,葉室溫度范圍控制在(30±0.5) ℃,相對濕度范圍控制在(40±5)%,光響應曲線的光合有效輻射強度設定在2 000、1 800、1 500、1 200、1 000、800、600、400、200、100、50、0 μmol·m-2·s-1共12 個光照強度梯度。儀器開機時,預先設定估計的飽和光強和最佳CO2濃度,光誘導黃精葉片30 min,等到儀器測得的凈光合速率(Pn)參數穩定在小數點后1 位數時記錄數據。7 種黃精屬植物均隨機選定3 株生長良好的植株,每株選擇3 片中上部位同一水平、成熟無病蟲害的葉片,盡量保持葉片在植株上同一位置測量。后續采取葉子飄直角雙曲線修正模型擬合光響應曲線,得到飽和光強(Isat)、初始斜率(α)、最大凈光合速率(Pnmax)、暗呼吸速率(Rd)和光補償點(Ic)等光合參數[13-14]。
1.2.4 CO2 響應曲線的測定
選取7 種黃精屬植物中部生長條件較好的成熟葉片,在晴天用Li-6400 測定其CO2響應曲線。該儀器配有CO2注射器,可為葉片室提供穩定可調的CO2氣源,測定中使用CO2小鋼瓶提供測定所需要輸出的穩定CO2濃度。在測定過程中,葉室內的光合有效輻射強度控制在1 000 μmol·m-2·s-1,參比室內的CO2濃度分別設定在0、50、100、200、400、600、800、1 000、1 200、1 500、1 800、2 000 μmol·m-2·s-1,共有12 個CO2的濃度梯度,重復測定3 次。
1.3 數據處理與分析
數據整理采用Excel 2020 軟件,數據分析采用SPSS 20.0 軟件,多重比較采用Duncan 方法,顯著性水平P≤0.05,極顯著水平P≤0.01。利用光合計算軟件對實測的光合數據進行擬合,得到各項目的光合計算參數,進而對7 個黃精種間的光合參數進行分析[15]。
2 結果與分析
2.1 7 種黃精屬植物的凈光合速率日變化
7 種黃精屬植物均不存在午休現象,總體走勢相近,7 種黃精屬植物中點花黃精與黃精的凈光合速率高于其余5 種,其凈光合速率日變化呈明顯的單峰曲線。上午隨著外界光照輻射強度的增強黃精屬植物的凈光合速率迅速增加,下午隨著外界光照輻射強度的減弱而隨之降低。
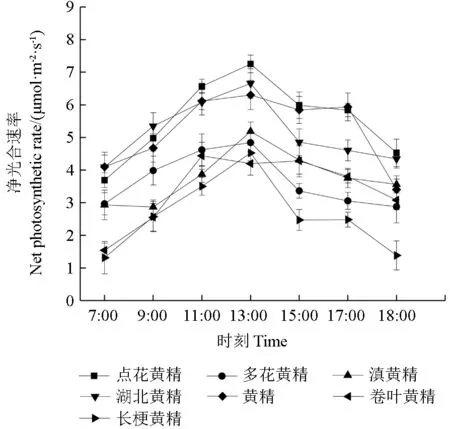
圖1 7 種黃精屬植物的凈光合速率日變化Fig.1 Daily changes of net photosynthetic rate of seven Polygonatum plants
2.2 7 種黃精屬植物的葉片葉綠素含量差異
由表1可以看出,7 個黃精屬植物種間的葉片葉綠素含量差異極顯著(F=23.3,P<0.01)。多重比較發現,黃精的葉片葉綠素含量最高,為1.83 mg·g-1,顯著高于其它種,卷葉黃精的葉片葉綠素含量最低,為1.07 mg·g-1,顯著低于其它種,其余黃精屬植物葉片的葉綠素含量在1.25~1.54 mg·g-1之間。
2.3 7 種黃精屬植物的水分利用率差異
表1所示7 個黃精屬植物種間的水分利用效率差異極顯著(F=11.33,P<0.01),其中點花黃精(5.13 μmol·mol-1)和長梗黃精(5.05 μmol·mol-1)的水分利用效率顯著高于其它種,具有更好的抗旱性能,黃精(3.47 μmol·mol-1)、卷葉黃精(3.42 μmol·mol-1)和多花黃精(2.50 μmol·mol-1)亦顯著高于湖北黃精(1.94 μmol·mol-1)和滇黃精(1.83 μmol·mol-1),其中滇黃精的水分利用率最低,其抗旱性較差。
2.4 7 種黃精屬植物種間的氣孔導度差異
表1顯示7 種黃精屬植物種間的氣孔導度間差異不顯著(F=0.925,P>0.05),在7 種黃精屬植物中滇黃精的氣孔導度最高,為0.122 mmol·m-2·s-1,湖北黃精的氣孔導度最低,為0.737 mmol·m-2·s-1,所有種均分布于0.737~0.122 mmol·m-2·s-1之間,種間均不存在顯著性差異。

表1 7 種黃精屬植物光合指標參數的比較?Table 1 Comparison of photosynthetic index parameters of seven Polygonatum plants
2.5 7 種黃精屬植物種間的胞間CO2 濃度差異
表1中7 種黃精屬植物種間的胞間CO2濃度不存在顯著差異(F=0.638,P>0.05),其中點花黃精的胞間CO2濃度最高(352.6 μmol·mol-1),長梗黃精的胞間CO2濃度最低,為312.6 μmol·mol-1,種間不存在顯著差異。
2.6 7 種黃精屬植物種間的蒸騰速率差異
表1所示為7 種黃精屬植物種間的葉片蒸騰速率差異顯著(F=21.0,P<0.01),其中湖北黃精和滇黃精的葉片蒸騰速率最高,分別為2.92和2.43 mol·m-2·s-1,顯著高于其它種,兩者水分利用率最低;長梗黃精的葉片蒸騰速率最低,為0.62 mol·m-2·s-1,顯著低于其它種,其水分利用率最高,試驗結果表明黃精的水分利用率與葉片蒸騰速率存在顯著的負相關關系。
2.7 7 種黃精屬植物的光響應曲線及其相關參數
從圖2可以看出,7 種黃精屬植物的葉片Pn值對光合有效輻射的響應曲線存在差異。7 種黃精屬植物的Pn值在光照強度為0~500 μmol·m-2·s-1范圍內會隨著PAR 的增強,Pn也隨之迅速增加。PAR 在500~1 000 μmol·m-2·s-1范圍內,Pn上升趨勢緩慢。在PAR 在1 000~2 000 μmol·m-2·s-1范圍內,7 種黃精屬植物的Pn的變化趨勢相近,均呈下降趨勢,點花黃精和卷葉黃精比較穩定,下降速度較慢,其余5 種黃精屬植物保持持續下降趨勢,下降速度較快,表明點花黃精和卷葉黃精對光照的利用區間較廣。
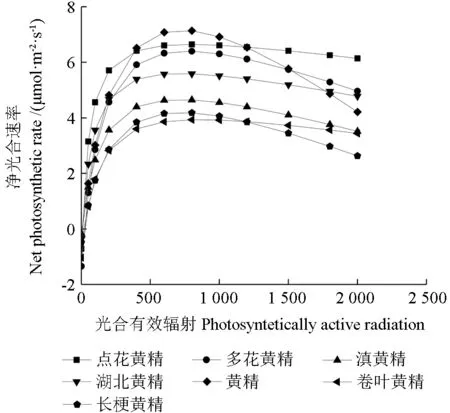
圖2 7 種黃精屬植物的光響應曲線Fig.2 Light response curves of seven species of Polygonatum
根據表2的多重比較發現多花黃精的暗呼吸速率最高,卷葉黃精的光補償點最高。7 種黃精屬植物最大凈光合速率在4.03~7.15 μmol·m-2·s-1之間,黃精的最高為7.15 μmol·m-2·s-1,卷葉黃精的最低為7.99 μmol·m-2·s-1;光飽和點在709.44~861.85 μmol·m-2·s-1之間,其中卷葉黃精最高,表明其利用強光的能力最強,湖北黃精的最低。光補償點在3.66~13.72 μmol·m-2·s-1之間,其中卷葉黃精的光補償點最高為13.72 μmol·m-2·s-1,滇黃精的最低為3.66 μmol·m-2·s-1,表明滇黃精更適應弱光環境。
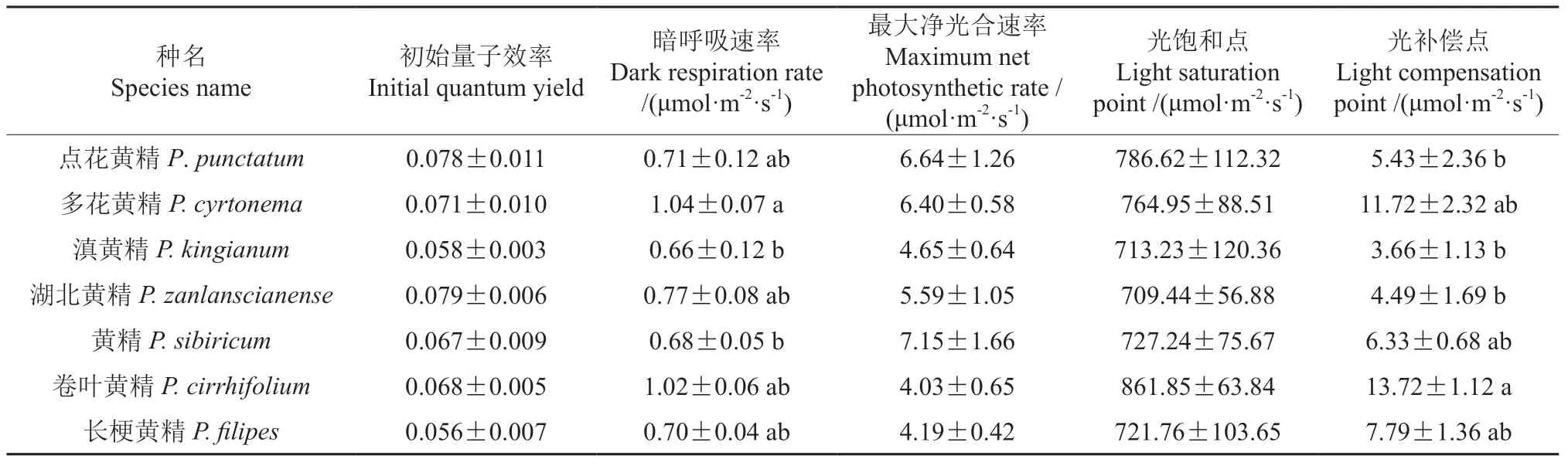
表2 光響應曲線相關參數Table 2 Related parameters of light response curve
2.8 7 種黃精屬植物CO2 響應曲線
從圖3可以看出,7 種黃精屬植物的葉片Pn值對CO2的響應曲線趨勢一致。CO2濃度在0~600 μmol·mol-1范圍內,隨著CO2濃度的增加,凈光合速率迅速增加。CO2濃度在600~1 000 μmol·mol-1范圍內,凈光合速率增加速率變慢。CO2濃度在1 000~2 000 μmol·mol-1范圍內,凈光合速率緩慢增加,不同種間表現略有差異。
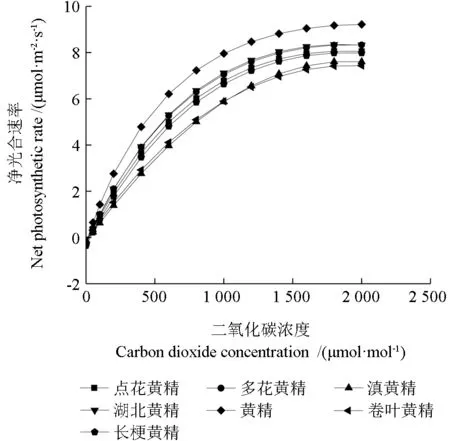
圖3 7 種黃精屬植物CO2 光響應曲線Fig.3 CO2 light response curves of seven species of Polygonatum
2.9 光合作用參數間的相關性
通過對7 種黃精屬植物的光合參數的平均值間做相關性分析可以看出(表3),黃精屬植物葉片凈光合速率分別與葉片的胞間CO2濃度和水分利用率存在顯著的正相關關系,相關系數分別為0.691 和0.905(P<0.05);葉片的氣孔導度和蒸騰速率間也存在顯著的正相關關系,相關系數為0.712(P<0.05);胞間CO2濃度與葉片蒸騰速率間存在顯著正相關關系(R2=0.64,P<0.05)。
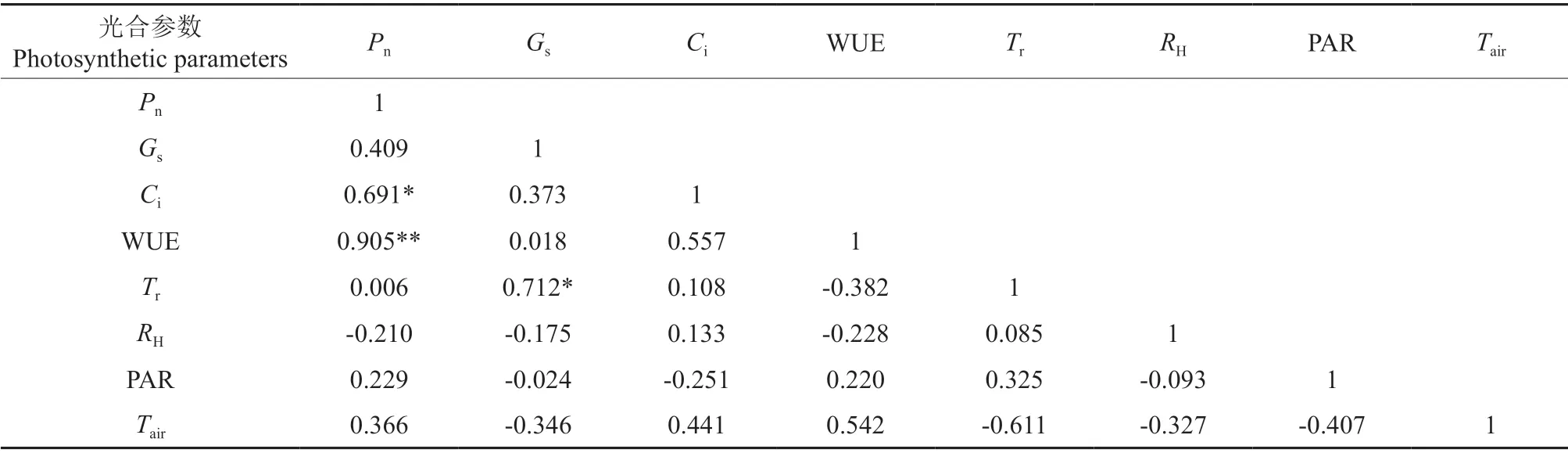
表3 光合作用參數間的相關性?Table 3 Correlation among photosynthesis parameters
2.10 7 種黃精屬植物的光合特性評價
光合速率的大小受內外多種因素的影響,試驗選取了7 種黃精屬植物光合特性的5 個主要指標:凈光合速率、氣孔導度、蒸騰速率、胞間CO2濃度和水分利用效率的平均值,在SPSS 數據處理系統中,選擇相似矩陣和最遠鄰元素聚類方法,聚類成3 類[16-17]。由聚類結果結合表1~2 中7種黃精屬植物光合速率的大小可以得出,7 種黃精可分為3 類:多花黃精、黃精和滇黃精分為一類,其綜合光合能力最好;湖北黃精、卷葉黃精和點花黃精同屬一類,綜合光合能力次之;長梗黃精是其中綜合光合能力最差的一類。
3 結論與討論
1)7 種黃精屬植物的葉片凈光合速率與胞間CO2濃度和葉片的水分利用率呈顯著性正相關。7 種黃精屬植物葉片葉綠素含量存在顯著性差異,但其凈光合速率與葉綠素含量之間并無顯著相關關系,這一點在其它作物上也有相似的報道[18]。植物葉片的凈光合速率是評價植物光合作用強弱的一個重要指標,影響葉片凈光合速率的因素有兩個方面,一方面是葉片本身,如葉齡、葉形、氣孔阻力、蒸騰速率、葉綠素含量及葉向等均對葉片光合速率產生影響;另一方面主要受外界環境,如溫度、光照強度、空氣濕度等的影響[19-20]。對凈光合速率和光合色素以及其它指標進行相關性分析,找出與凈光合速率相關的指標,可以為生產中提高植物的光合能力提供一定的理論依據[21-22]。
2)7 種黃精屬植物中,點花黃精和長梗黃精的水分利用效率相對較高,葉片蒸騰速率相對較低。水分利用效率是指單位面積水分消耗所能同化的光合產物量[23],是一個較為穩定的衡量碳固定和水分消耗比例的指標。研究表明,干旱脅迫下的小紫珠、日本莢蒾幼苗的光合速率都受到顯著的影響[24-25]。通過測定WUE 能夠闡明植物的氣體交換特性以及抗旱性機理,從而選擇出低耗水、生產效率高、抗性強的黃精屬植物[26]。本試驗條件下,7種黃精屬植物的WUE 種間差異顯著,表明不同種間水分利用能力差別較大,點花黃精和長梗黃精的水分利用效率較高,與其對應的葉片蒸騰速率較低,表明這兩個種的耐旱性相對更強。
3)7 種黃精屬植物的光飽和點接近陽生植物,光補償點接近陰生植物,CO2飽和點高于正常外界濃度。7 種黃精屬植物雖然喜陰,但是對于有效光合輻射有著比一般陰生植物更高的需求,種植中在提供陰涼環境的同時也應盡量滿足其光照需求[27]。從7 種黃精屬植物的CO2飽和點來看,均能適應較高濃度的CO2水平。所以,隨著大氣CO2濃度的增加,如果在其它條件滿足的條件下,將更有利用黃精的生長和產量的提高。
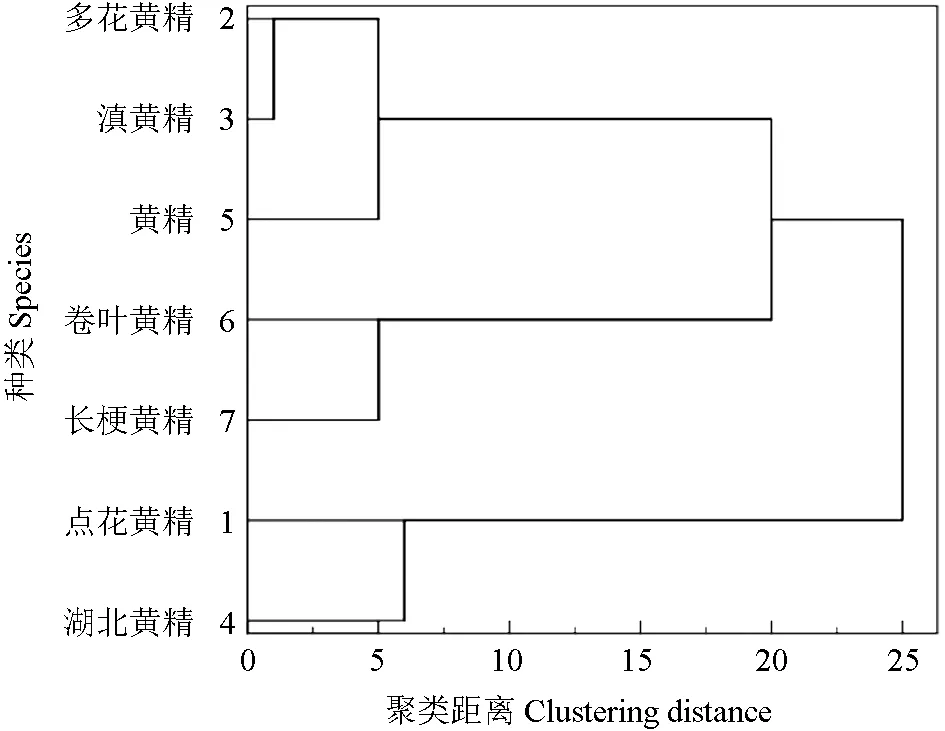
圖4 光合特性聚類分析圖Fig.4 Cluster analysis of photosynthetic characteristics
光合指標的聚類分析顯示,7 種黃精屬植物可聚成3 類,其中多花黃精、黃精和滇黃精歸為光合性能最優類。
多花黃精、黃精和滇黃精是《中國藥典》認定的藥用黃精正品,其種植歷史悠久,結合本研究結果,可以加大推廣多花黃精、黃精與滇黃精的種植面積,并加強保護多花黃精、黃精與滇黃精的野生種質資源。點花黃精、湖北黃精和卷葉黃精的光合性能次之,應結合產量、品質等綜合特性合理進行種植。長梗黃精的光合特性較差,應結合其它性狀綜合考慮是否適宜栽培。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
