大豆害蟲黑毛薊馬研究進展
2021-07-11 07:09:46趙一瑾許嘉嘉史樹森
中國農學通報 2021年18期
關鍵詞:大豆
趙一瑾,高 宇,王 迪,許嘉嘉,史樹森
(吉林農業大學植物保護學院,長春130118)
0 引言
薊馬蟲體微小、生活隱蔽、不易被發現,是一個龐大而獨特的類群。這類害蟲在國內各個大豆產區普遍發生,目前已知3科12屬21種[1]。由于不合理地施用化學農藥,一些種類的種群數量在局部地區呈快速上升趨勢,發生面積逐年擴大,成為大豆高產優質的重要制約因素之一。黑毛薊馬(Thripsnigropilosus)隸屬纓翅目(Thysanoptera)、薊馬科(Thripidae),異名Thrips nigropilosalaevior、Thripslactucae、Thripsumbratus、Thripspilosissima,又稱為豆黃薊馬等。該害蟲曾在黑龍江省多個地區大發生,1985年在密山農場造成較大損失,造成補種甚至部分地塊毀種;1989年在克山農場0.8萬hm2大豆田大發生;1990年在牡丹江、佳木斯等地農場3.37萬hm2大豆田大發生[2];1992年在克拜地區發生較重[3];1999—2000年在訥河市大發生,超過0.8萬hm2大豆田受害嚴重[4-5];在綏化、哈爾濱等地也有發生為害[6-8]。此外,在肯尼亞嚴重為害除蟲菊(Chrysanthemumcinerariaefolium)[9],在日本為害菊花(Chrysanthemummorifolium)和魁蒿(Arternisia princeps)[10],在英國為害長葉車前(Plantagolanceolata)和沿海車前(Plantagomaritima)[10],在美國南卡羅來納州為害小白菊(Tanacetumparthenium)、在夏威夷州為害黃瓜(Cucumissativus)和萵苣(Lactucasativa)[11]、在阿拉巴馬州發現其為害棉花,在新西蘭為害翼薊(Cirsiumvulgare)。在中國北京溫室內蒲公英(Taraxacummongolicum)上也嚴重發生,造成了不同程度的經濟損失[12]。近幾年來,隨著國際貿易往來的增多,生物入侵事件頻頻發生,為了監測和預防黑毛薊馬暴發成災,避免可能造成的危害,筆者對其分布與寄主、形態特征、危害特點、發生規律、生活習性、發生與生態因子的關系以及綜合防控等方面進行介紹,以供深入研究參考。
1 分布與寄主
該蟲在北半球廣泛分布、寄主范圍廣,現已知國內分布于黑龍江、吉林、江蘇、廣東、四川、貴州;國外分布于朝鮮、日本、土耳其、以色列、立陶宛、俄羅斯、捷克、斯洛伐克、波蘭、阿爾巴尼亞、羅馬尼亞、匈牙利、奧地利、荷蘭、瑞典、丹麥、西班牙、法國、芬蘭、瑞士、德國、英國、美國、埃及、斐濟、澳大利亞、新西蘭,中亞、外高加索地區和北非地區等[12-14]。已報道的寄主植物有菊科、豆科、茄科、葫蘆科等,在中國主要為害大豆,還為害煙草(Nicotianatabacum)、除蟲菊、菊花、小白菊、蒲公英、黃瓜、馬藍(Baphicacanthuscusia)、鐵杉(Tsuga chinensis)、蓍 (Achilleamillefolium)、萵苣、三葉草(Trifoliumspp.)、歐洲薊(Cirsiumvulgare)、蠶豆(Vicia faba)、長葉車前、沿海車前、甜瓜(Cucumissativae)、辣椒(Capsicumannuum)、羅勒(Ocimumbasilicum)、山艾(Salviaofficianalis)等[9,12-17]。
2 形態特征
成蟲:雌蟲體長1~1.2 mm,體黃色,各腹節間褐色,觸角節Ⅰ很淡,Ⅱ、Ⅲ黃,Ⅳ~Ⅷ黃棕色,足淡黃色,鬃和翅鬃黑色。胸部和腹部兩端常有不規則灰棕斑。觸角7節,第7節極小。單眼前后有橫線紋。單眼間鬃顯著粗于其他頭背鬃,在前、后單眼中途,位于前、后單眼外緣連線上。單眼后鬃距后單眼近,在復眼后鬃前。復眼后鬃圍眼排列。前胸背片布滿橫紋,背片鬃24根;中胸盾片布滿橫紋,鬃較細;后胸盾片前中部網紋較大,兩側有縱線紋。前翅略黃色,前緣鬃21根,前脈基部鬃6~8根,端鬃2根,后脈鬃11根。腹部節Ⅱ-Ⅷ背片和腹片線紋輕微。節Ⅱ背片側緣縱列鬃3根。節Ⅴ背片微梳小于Ⅵ-Ⅷ節微梳。節Ⅷ背片后緣梳毛細,中部稀疏,兩側缺。背側片和腹片無附屬鬃。節Ⅶ腹片后緣中對鬃在后緣之前。雄蟲與雌蟲相似,體長0.6 mm,短翅型,腹部腹片節Ⅲ-Ⅶ有啞鈴形橫腺域。本種個體(翅長、體色、鬃)變異大,雌蟲有長翅,半長翅、短翅的不同類型,雄蟲罕見,通常是短翅的(圖1)[9]。
若蟲:與成蟲相似,無翅,1齡若蟲體長0.3~0.4 mm,初孵無色,復眼紅色,幾小時后體色變黃;2齡若蟲體長0.6~1.0 mm,體黃色;3齡若蟲體白色,體長約1.0 mm,呈透明狀,翅芽出現,長達腹部第2節,觸角伸向外前方,分節不明顯,呈鞘囊狀,行動遲緩,稱前蛹;4齡若蟲體白色,單眼出現,翅芽長達腹部第5節,翅脈可見,不行動,稱擬蛹(圖1)[9]。
卵:腎形,長約0.25 mm,寬約0.14 mm,近無色(圖1)[9]。
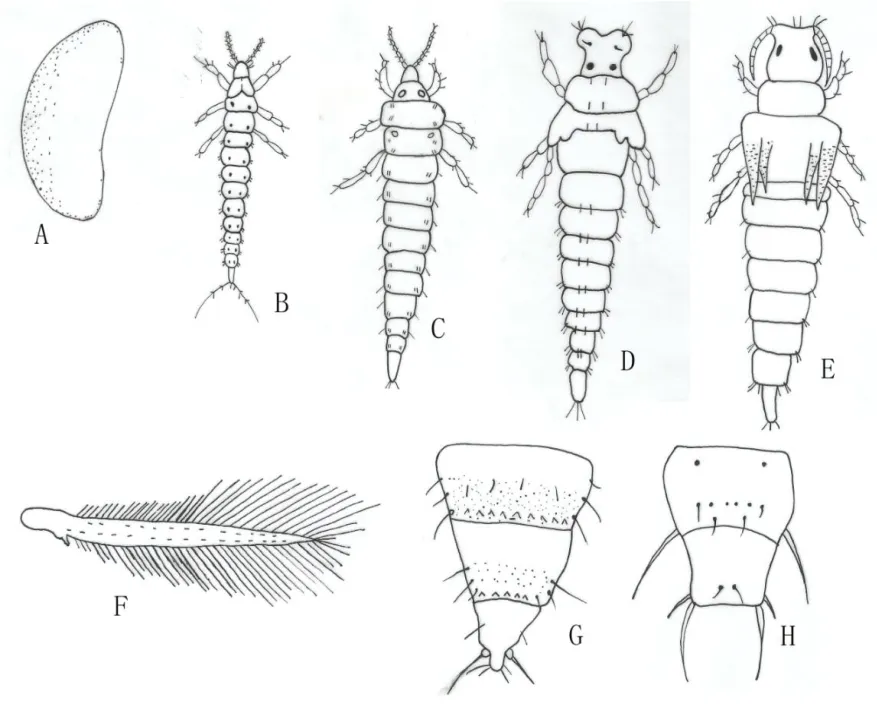
圖1 黑毛薊馬形態圖
3 危害特點
周弘春等[5]觀察發現成蟲和1~2齡若蟲為害大豆葉片和嫩莢,較少為害花器,葉片表皮被害部位表面發白并逐漸枯死變褐;幼嫩新葉受害表現為皺縮、卷曲、葉色褪綠,嚴重時葉面出現灰白色斑點,葉片局部枯死導致植株矮小,生長點被害,植株出現多頭現象或停止生長,生長長勢逐漸減弱甚至正株死掉。生長后期花器被害,造成落花、落莢,對產量影響很大。楊興勇[9]觀察發現成蟲和若蟲均為害大豆的芽、葉片和花,特別是在大豆幼苗期危害生長點和開花期為害花器較為嚴重。幼苗期生長點受害后變黃凋萎,嚴重時不能繼續延伸,開花期大多數在花內取食,搗散花粉,破壞柱頭,吸收花器營養,造成花的營養不良或引起不孕,形成癟莢或落花落莢,受害葉片主要是嫩葉,致葉片形成不規則的成片枯點,初呈淡黃色,漸漸隨著組織的完全枯死而呈黃褐色[3]。此外,已證實黑毛薊馬不是傳播斑萎病植物病毒的介體昆蟲。
4 發生規律
在黑龍江密山地區1年發生5~6代,在訥河地區干旱年份發生6~8代,世代重疊明顯(見表1)[5]。以成蟲在枯枝落葉及薊等雜草葉鞘內越冬,大多數越冬成蟲是短翅型[18]。雄蟲生活力較弱不能越冬,越冬成蟲均為雌性[19-20]。翌年5月中旬越冬成蟲陸續出現,先在小薊(Cirsiumsetosum)上取食為害,5月下旬轉移到剛出苗的大豆上為害,甚至在尚未張開的子葉間亦有薊馬出現,6月初豆苗上數量漸增,6月中旬越冬成蟲出現量達到高峰,然后薊馬成蟲和若蟲混合發生。6月下旬到7月下旬達為害盛期,大豆受害最嚴重。7月中旬以后,大豆植株長大,受害相對減輕。9月中旬大豆成熟,遷回到小薊上越冬[2,4-5]。
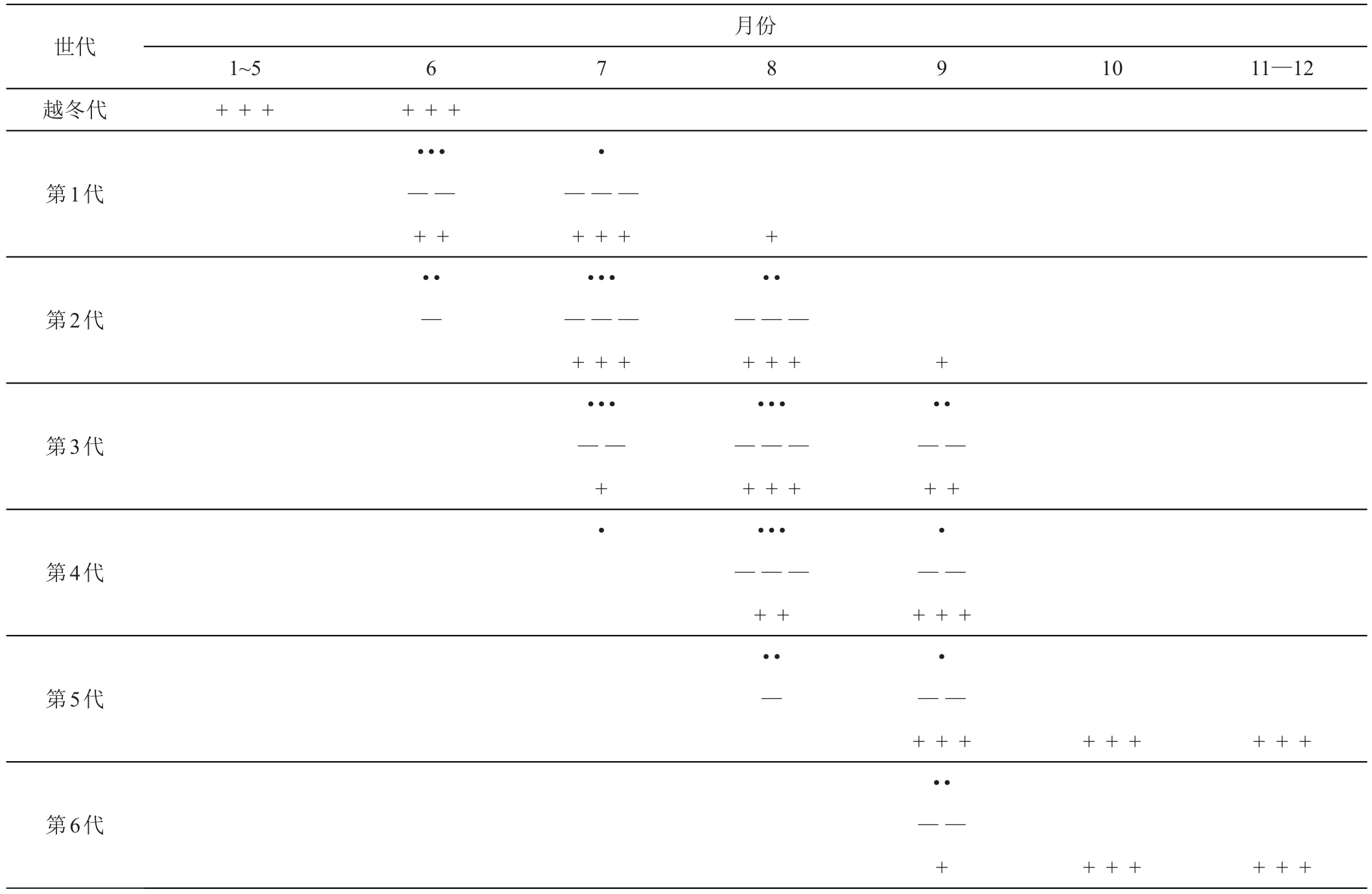
表1 黑毛薊馬的年生活史表
5 活動習性
初孵若蟲一般只在孵出的葉片上爬行,喜群集取食,2齡若蟲較活潑,常常爬行到其他葉片上尋找幼嫩葉片取食。成蟲行動活潑,能彈跳。雌成蟲很少起飛,但爬行很快,雄成蟲很活潑并起飛頻繁,可借助風力進行稍遠距離的遷移,一次只能飛行數米遠,高度不超過豆株上部1 m。從始發期到盛發期,約需4~5天,薊馬即可蔓延至全田。成、若蟲畏強光、趨嫩,每日10:00前和16:00后為活動盛期,中午強光照射則潛伏于葉背或剛長的嫩葉上棲息,在葉上往往依附于葉脈或凹坑的邊緣。黑毛薊馬可行孤雌生殖也可行兩性生殖,雌雄蟲的交尾行為一般分為4個步驟(圖2)[21]。越冬代雌成蟲行孤雌生殖繁殖出第1代。在第1代成蟲中出現雄蟲,此時田間雌雄性比為2:1。以后各代雄蟲數量變化不大,到8月份田間雌雄性比為1.5:1。雌成蟲具有鋸狀產卵器,可將卵產于植物組織內,產卵部位多為幼嫩葉片的近葉脈處。卵散產成小片集中,一處產幾粒到十幾粒。中等溫度和短日照光周期會抑制雌蟲的繁殖[20]。
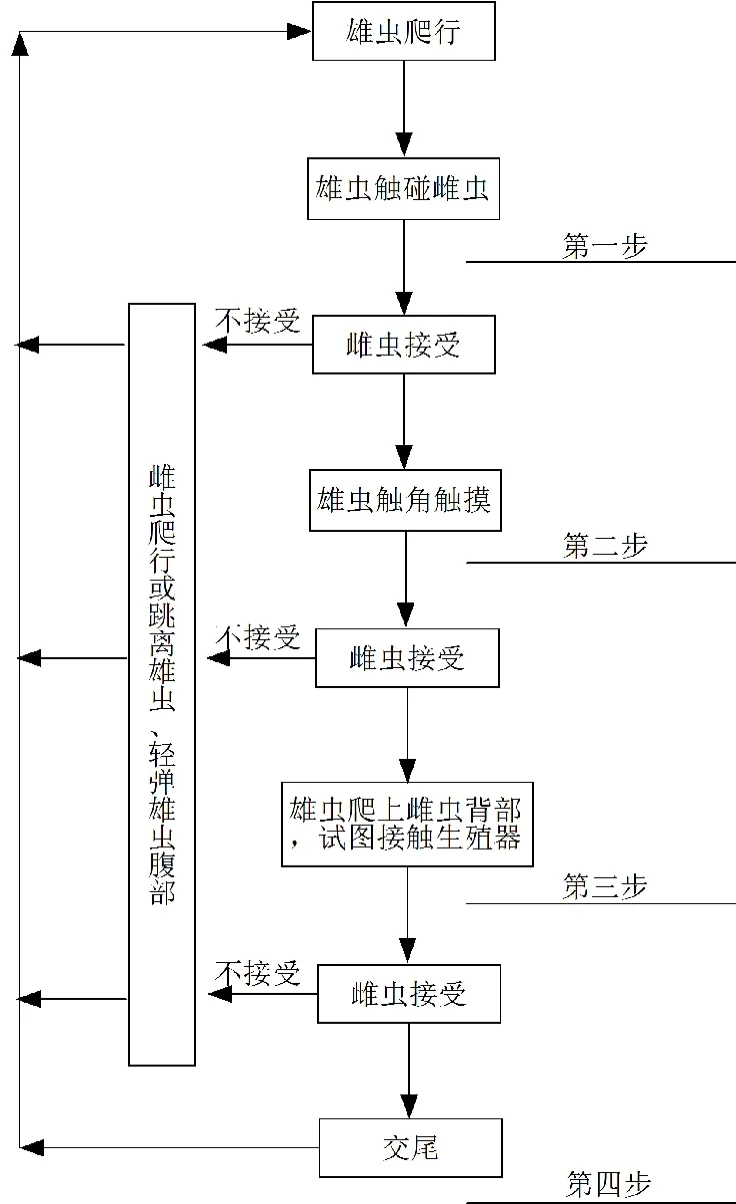
圖2 黑毛薊馬的交尾行為示意圖
6 發生與生態因子的關系
該蟲的發生與危害常受氣候的影響,一般溫暖干旱有利于大發生,低濕多雨對其發生不利。研究發現,當月降水量超過40 mm時,薊馬種群數量下降明顯[9]。其發生多少還與栽培條件有關,早播田比晚播田發生重,崗地比低洼地勢發生重,早熟和小粒型品種易感蟲。林志偉等[22]調查了不同時期的蟲口數量,Iwao回歸分析、Taylor冪法則分析和頻次擬合度分析表明,該蟲呈聚集分布(負二項分布),個體間存在相互吸引作用,分布基本成份是個體群,種群聚集度對種群密度有依賴性,有蟲株率和蟲口密度存在相關關系。
各世代歷期因發生時期不同而有差異。該蟲的第一個和最后一個世代歷期長,平均24天,其他3~4個世代平均歷期16.7天。在大豆上雌成蟲壽命平均17.5天,雄成蟲壽命平均5.6天。若蟲各齡歷期隨著季節溫度變化而異。6月平均溫度17.9℃,1~2齡為4天,3~4齡為8天;7月平均溫度23℃,1~2齡為6天,3~4齡為3天;8月平均溫度 18.2℃,1~2齡為6天,3~4齡為 5天[23]。卵、1~2齡若蟲、3~4齡若蟲及世代的發育起點溫度分別為10.5℃、13.7℃、133.3℃、11.7℃,有效積溫分別為77.9℃、32.1℃、34.7℃、165.7℃。
Anyango等[24]在室內恒溫下觀察該蟲在除蟲菊上的生活史以及溫度對其生活史的影響,發現成蟲具有性二型現象,雌雄比為11:2,在除蟲菊上成蟲壽命為1~21天,發育歷期隨著溫度的升高而降低;當以發育速率表示時,發現隨著溫度的升高,2齡和4齡若蟲的發育比其他階段更快。Nakao等[25]通過研究光周期敏感期是否是雌蟲翅型、生殖滯育和滯育誘導敏感期的決定因素,發現卵和幼蟲期是翅型發育的敏感階段,短日照是在1日齡幼蟲短翅型發育所必需的,從剛孵化前的卵到8日齡若蟲的任何時期內,長日照光周期部分或完全抑制了短翅型的發育,短日照光周期誘導生殖滯育,高溫抑制滯育誘導;卵期和若蟲期對光周期敏感,卵期和低齡若蟲期比高齡若蟲期對光周期更為敏感;短翅型進入滯育的雌性比例比長翅型和中間型更高。在實驗室飼養的種群密度越高,短翅型的比例也越高;隨著飼養密度的增加,從孵化到成蟲羽化期其死亡率和發育期也隨之增加,在所有翅型中,短翅型的發育期最長,短翅型個體的產卵期較長,頭較小,生殖力較低[26-27]。
7 綜合防控
大豆有害薊馬綜合治理一般應采取以農業防治為基礎,物理防治和生物防治為主,化學防治為輔的綜合治理策略[1]。例如,收獲后及時進行翻耕,冬春及時清除田內外雜草,消滅孳生及越冬場所,降低越冬蟲量,減少第二年該蟲基數。實行大豆與非寄主作物輪作,由于該蟲飛翔能力差,遠離上年田塊種植大豆,可減輕危害。加強田間水肥管理,干旱年有條件的地塊進行噴灌,可減輕薊馬危害。有條件的地區也可根據需要及時在田間使用有色黏板等物理方法誘殺成蟲,以達到降低種群密度減輕危害的目的[28]。天敵有捕食性薊馬、花蝽、蜘蛛、瓢蟲等,對薊馬種群具有一定的控制作用[29]。
研究表明,黑毛薊馬在菊花上導致產量減少是由花數減少引起的,防治指標為16.64頭/葉[30-33]。但大豆上該蟲的防治指標尚無報道,應注意兼防兼治地下害蟲及其他苗期害蟲,可選用大豆種衣劑,隨種肥下地[34]。大豆苗期薊馬發生危害輕或不發生,盡量使用選擇性殺蟲劑,或減少施藥次數等來保護天敵,使天敵的種群維持在一定水平,以便控制薊馬種群數量急劇上升[2,5]。當該蟲發生量較大,非化學防治技術不足以控制種群數量時,可噴施1.8%阿維菌素EC和60 g/L乙基多殺菌素SC[12]。一般認為長期使用化學農藥容易導致薊馬產生抗藥性[35],也有學者認為關于黑毛薊馬近期很少受到關注,可能是由于廣泛使用化學農藥導致其生態位發生變化[10,36],所以該蟲對農藥是否具有較強的抗藥性,今后需予以澄清。
8 展望
現有研究初步明確了黑毛薊馬的危害特點、生活史、活動習性、發生與生態因子的關系等,還存在如下主要問題:大豆田間薊馬群落變化趨勢以及黑毛薊馬的種群消長情況尚不十分明確;各地區大豆黑毛薊馬生活史以及發生與生態因子的關系還需要深入研究;黑毛薊馬仍依賴于化學藥劑防治。建議今后開展下述研究工作:(1)研究新的氣候和種植制度下,大豆薊馬群落結構及演替趨勢;(2)繼續開展基礎生物學、生態學特性研究;(3)為了制定科學的綜合治理方案,進一步開展測報技術和綠色防控技術研究。
致謝:感謝吉林農大植保專業夏婷婷同學繪形態圖。
猜你喜歡
農業科技通訊(2023年1期)2023-02-12 07:09:18
今日農業(2022年16期)2022-11-09 23:18:44
中國化肥信息(2022年7期)2022-08-31 01:29:28
中國化肥信息(2022年5期)2022-08-30 01:58:26
今日農業(2021年13期)2021-11-26 11:50:54
今日農業(2021年20期)2021-11-26 01:23:56
今日農業(2021年14期)2021-10-14 08:35:34
下一代英才(酷炫少年)(2018年6期)2018-07-09 03:17:44
農產品市場周刊(2017年4期)2017-03-03 19:40:05
兒童故事畫報·智力大王(2015年10期)2016-01-27 01:01:35
