哲羅鮭性別特異性標記篩選
2021-07-17 03:25:06佟廣香唐國盤張永泉尹家勝匡友誼
水生生物學報 2021年4期
佟廣香 唐國盤 徐 偉 張永泉 尹家勝 匡友誼
(1. 中國水產科學研究院黑龍江水產研究所, 黑龍江省冷水性魚類種質資源及增養殖重點開放實驗室, 哈爾濱 150070;2. 河南牧業經濟學院動物科技學院, 鄭州 450046)
哲羅鮭Hucho taimen(Pallas)隸屬鮭形目(Salmoniformes)、鮭科(Salmonidae)、哲羅魚屬(Hucho),是我國珍稀名貴冷水性魚類之一, 肉質細嫩, 味道鮮美[1]。1998年哲羅鮭被列入中國瀕危動物紅皮書[2,3]。近年來哲羅鮭的資源量不斷下降, 已經很難見到野生群體[4—8], 2004年被列入中國物種紅色名錄。
哲羅鮭是鮭科魚類中個體最大的魚類, 生長迅速, 抗病力強, 營養價值高, 養殖條件與虹鱒類似, 是優秀的養殖對象[5,9]。在養殖條件下, 哲羅鮭需要4—5年才能初次性成熟, 性成熟前, 無法通過外形來判斷其性別; 在性成熟后, 由于生活環境不同, 婚姻色存在差異, 也很難通過婚姻色鑒定雌、雄[1,10,11]。哲羅鮭雌魚懷卵量大, 魚卵可用于加工魚子醬, 因此單性養殖和性別控制育種具有較高的應用價值。怎樣快速、準確并在幼魚期鑒定哲羅鮭遺傳性別, 指導養殖過程中雌、雄比例,有目的的養殖, 節約養殖成本, 成為制約哲羅鮭產業發展的重要因素之一。本研究根據虹鱒Y染色體特異性序列, 從哲羅鮭基因組草圖中篩選出性別特異性標記, 并建立準確的性別鑒定方法, 為哲羅鮭的性別分化研究、性別控制育種和單性養殖等奠定基礎。
1 材料與方法
1.1 樣本采集及DNA提取
本研究材料來源于中國水產科學研究院黑龍江水產研究所渤海冷水性魚試驗站。繁殖時鑒定雌、雄, 采集雌、雄魚各48尾鰭條貼在濾紙上陰干。取0.2—0.4 cm2的鰭條樣本, 用100 μL裂解液[裂解液成分: 蛋白酶K 0.5 mg/mL、1 mol/L Tris(pH8.0)10 mmol/L、KCl 50 mmol/L、0.3%Tween 20、0.3% NP 40]在PCR板內裂解樣本, 裂解程序為55℃ 2—4h, 98℃ 10min。樣本裂解后采用Vortex儀混勻, 混勻后1000—2000 r/min離心1—2min, 取上清液作為PCR擴增模板。
1.2 序列分析及引物設計
從NCBI下載虹鱒Y染色體序列(EU081756.1),采用BLAT程序在哲羅鮭基因組草圖中搜索出同源序列。用Primer 3在同源序列上設計性別特異性引物, 并在哲羅鮭線粒體12S rRNA序列上設計參照引物。參照引物用于消除非特異性擴增、樣本降解及加樣失誤等因素的影響。引物序列及退火溫度見表1。

表1 引物列表Tab. 1 Primer list
1.3 PCR擴增
PCR擴增反應體系為: 2×PCR Dream Taq master mix(Thermo Fisher, USA)10 μL, 樣本DNA裂解上清液2 μL, 10 μmol/L的上下游引物各0.1—1 μL,超純水補足20 μL。PCR程序設置為95℃變性3min;30個循環, 每個循環設置為95℃ 30s, 退火(退火溫度見表1)30s, 72℃ 30s; 最后72℃延伸5min。PCR產物分別用2%瓊脂糖凝膠電泳和8%的非變性聚丙烯酰胺電泳檢測。將特異性引物的PCR擴增產物純化、回收, 克隆至pMD18-T載體, 處理后, 轉入大腸桿菌感受態中, 篩選含有陽性插入片段的菌落進行測序。
1.4 引物濃度優化
本研究利用哲羅鮭基因組中與虹鱒Y染色體同源的序列設計性別鑒定引物, 在理想情況下, 雌性樣本中無擴增條帶, 而實際檢測過程中樣本DNA降解、加樣失誤、PCR擴增失敗等原因也會導致無擴增條帶。為消除此類問題的影響, 本研究引入了線粒體12S rRNA作為參照, 將性別特異性引物和12S rRNA引物在同一個PCR反應體系內擴增, 在參照引物有條帶的前提下, 用特異性引物有無條帶鑒定雌、雄。由于不同引物擴增效率存在差異, 因此需要對性別特異性引物和12S rRNA引物濃度比例進行優化。優化時設置4個濃度梯度, 在20 μL擴增體系內10 μmol/L的性別特異性引物上下游各1 μL,而10 μmol/L的12S rRNA參照引物上下游體積分別為1、0.5、0.25和0.1 μL。
2 結果
2.1 引物篩選
用雌、雄魚各3尾樣本初步篩選3對性別特異性引物和12S rRNA參照引物。由圖1可知, 在性別特異性引物中, 引物ST1在雌、雄魚樣本中均能擴增出133 bp條帶; 引物ST2在雌魚樣本中無擴增條帶, 在雄魚樣本中能擴增出153 bp條帶; 引物ST3在雌、雄魚樣本均能擴增出182 bp條帶。12S rRNA參照引物在雌、雄魚中均能擴增出251 bp條帶。此結果表明, 引物ST2可能是哲羅鮭雄魚特異性標記。

圖1 4對引物PCR產物瓊脂糖凝膠電泳圖譜Fig. 1 Electrophoresis of PCR products using four pairs of primers
2.2 PCR引物濃度優化
為提高檢測的準確率, 節約檢測成本和時間,本研究采用雙重PCR進行性別鑒定。由于不同引物的擴增效率存在差異, 因此需要對性別特異性引物和12S rRNA參照引物的濃度比例進行優化。在20 μL擴增體系內10 μmol/L的引物ST2固定為1 μL,而10 μmol/L的12S rRNA參照引物體積分別為1(圖2A)、0.5(圖2B)、0.25(圖2C)和0.1 μL(圖2D)。由圖2可知, 引物ST2和12S rRNA的4組比例均能擴增出條帶, 且隨著12S rRNA引物濃度的降低, 12S rRNA擴增條帶亮度呈下降趨勢, 引物ST2擴增條帶亮度呈現遞增趨勢。當引物ST2為1 μL, 12S rRNA為0.25 μL時(圖2C), 二者條帶亮度達到平衡, 條帶均比較清晰。因此在哲羅鮭性別鑒定中引物ST2和12S rRNA引物濃度比例設置為1∶0.25。
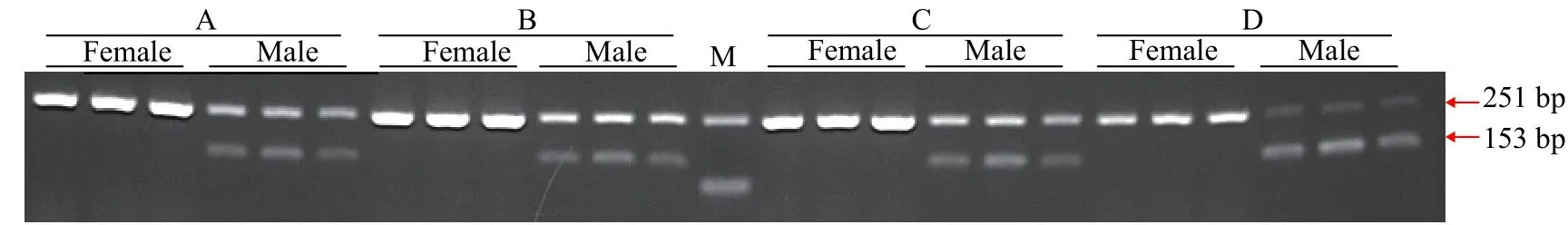
圖2 引物ST2和12S rRNA雙重PCR擴增瓊脂糖電泳圖譜Fig. 2 Electrophoresis of duplex PCR products using ST2 and 12S rRNA primers
2.3 性別特異性標記鑒定
為驗證引物ST2的有效性, 進一步用雌、雄魚各48尾樣本評估引物ST2的準確性。48尾雌、雄魚樣本擴增產物瓊脂糖凝膠電泳圖譜見圖3, 聚丙烯凝膠電泳圖譜見圖4。引物ST2在雌魚中均無擴增條帶, 在雄魚中均能夠擴增出153 bp的單一條帶;12S rRNA參照引物在雌、雄樣本中均能擴增出251 bp條帶。12S rRNA參照引物能夠擴增出條帶表明雌魚樣本條帶缺失并不是由于PCR擴增失敗、加樣失誤和DNA降解等原因造成的, 而是由雌、雄樣本間基因組DNA差異造成的。鑒于以上結果, 引物ST2可以作為哲羅鮭雄性特異性標記, 用于性別鑒定。引物ST2在檢測的48尾雌、雄魚樣本中, 性別鑒定準確率可達100%。
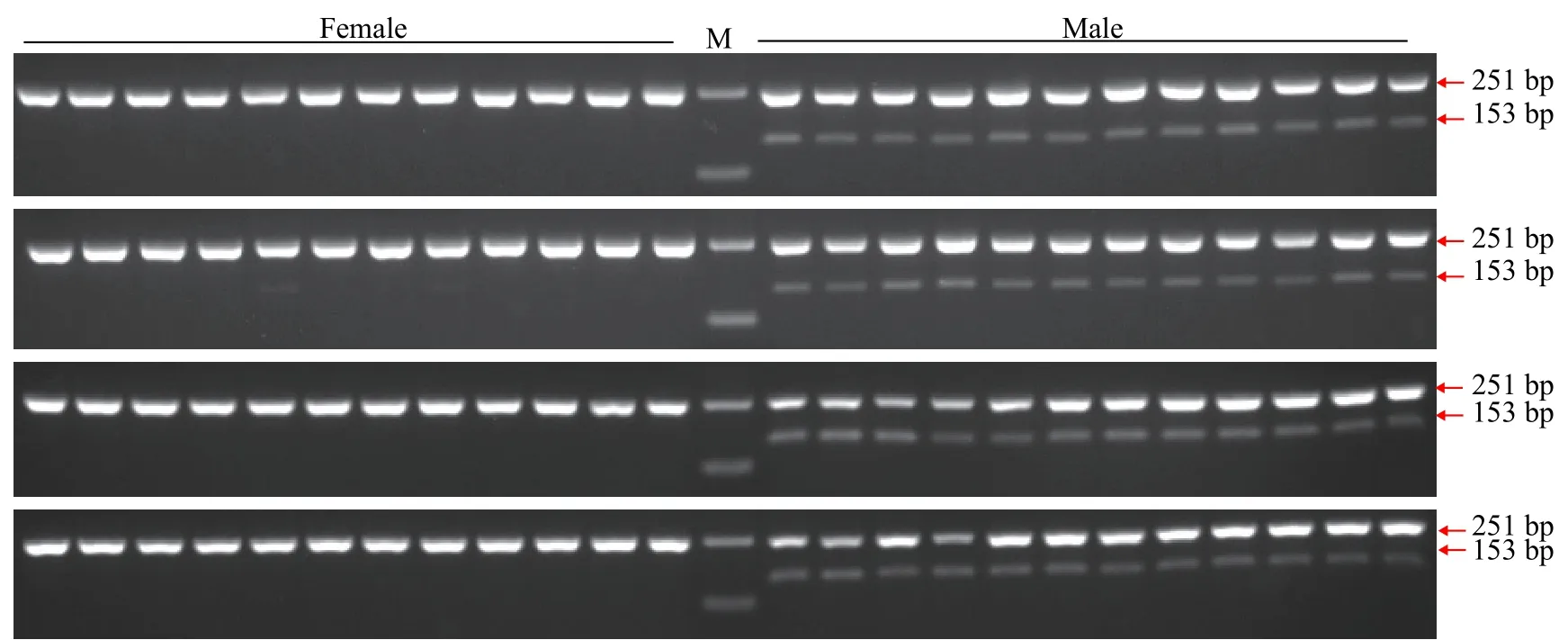
圖3 引物ST2和12S rRNA1組合雌、雄樣本的瓊脂糖電泳圖譜Fig. 3 Electrophoresis of duplex PCR products using ST2 and 12S rRNA primers in female and male samples
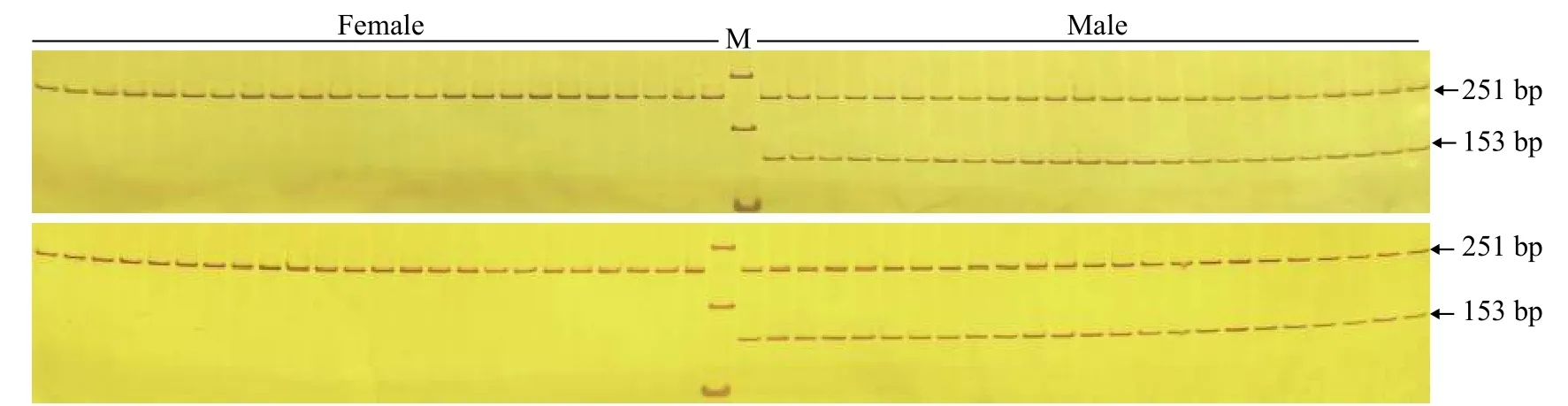
圖4 引物ST2和12S rRNA1組合雌、雄樣本的聚丙烯酰胺凝膠電泳圖譜Fig. 4 Non-denaturing PAGE electrophoresis of duplex PCR products using ST2 and 12S rRNA primers in female and male samples
2.4 雄性特異性基因篩選
將引物ST2的PCR擴增產物純化、回收, 克隆至pMD18-T載體, 處理后轉入大腸桿菌感受態中,篩選含有陽性插入片段的菌落進行測序。測序獲得153 bp序列(5′-TGTCAGGGTTGATTACAGTTCC TAAGGCATTTGCATTTTATCTCATGGTAGTGGTTG TGTCCTGCAGCCTCCCAACAGCCTTGTCGTT CTGTGGAGTTCATGTGGGATGTATATCAATCA TGGCTGGAGGTGTGATGAGGAATCAGACTG CAG-3′)。將該序列與哲羅鮭基因組(未發表)對比發現, 此序列與sdY基因位于同一Scaffold上, 并與sdY基因相鄰。除此之外, 還將該序列與哲羅鮭的近源物種多瑙河哲羅鮭(Hucho hucho)基因組進行了對比, 結果顯示此序列在多瑙河哲羅鮭基因組中存在2個拷貝, 分別位于QNTS01001159.1的ENSH HUG00000033926(lrp4-1)基因內和QNTS01000029.1的ENSHHUG00000034142(lrp4-2)內, 并與這2個基因7號外顯子匹配90 bp。
3 討論
魚類是低等脊椎動物, 進化相對原始, 由于大多數魚類的性染色體還處于未分化或者分化的早期階段, 因此很難通過性染色體的形態來辨別雌、雄。另外, 魚類胚胎發育過程中易受環境(特別是溫度)影響, 常導致魚類的表型性別(卵巢或精巢)和遺傳性別(基因型)不一致[12—16], 因此需要一種準確、快捷的方法來鑒定魚類的遺傳性別。分子標記是一種可以在DNA水平上直接反映基因組差異的方法, 與形態學標記[17,19]、細胞學標記和生物化學標記相比[20—23], 具有準確、快捷及數量豐富等優越性。目前多種鮭科魚類已經篩選出了性別相關的分子標記。如: Devlin等[24]篩選出大馬哈魚Y染色體上特異的DNA片段, 該片段可用來鑒定大馬哈魚遺傳性別; Yano等[25]發現了一種免疫相關基因進化成為虹鱒主要性別決定基因; Felip等[26]通過擴增片段長度多態性(Amplified fragment length polymorphism, AFLP)方法, 獲得了一些虹鱒性別特異分子標記。哲羅鮭也是鮭科魚類, 但性別相關的分子標記尚未見報道。本研究成功獲得了哲羅鮭雄性特異性的分子標記, 可以100%區分哲羅鮭的遺傳性別, 且僅用瓊脂糖凝膠電泳檢測就能獲得結果,方法簡單, 易于操作。
在鮭科魚類中, 基因sdY[25,27]是虹鱒、大西洋鮭和多瑙河哲羅鮭等15個不同物種的性別決定基因。本研究獲得的153 bp序列與sdY基因在哲羅鮭基因組草圖中位于同一Scaffold上, 與sdY基因相鄰。將這153 bp序列與多瑙河哲羅鮭基因組對比,發現此序列屬于lrp4基因的部分序列。lrp4基因是低密度脂蛋白受體4, 其主要功能是調節體內膽固醇平衡, 是膽固醇運輸的主要載體, 也有報道表明lrp4基因在小鼠胚胎發育E13和E14時期的睪丸中顯著高表達[28],lrp4是否為哲羅鮭Hucho taimen的性別決定基因, 還需要進一步實驗驗證。
哲羅鮭屬大型魚類, 商品魚養殖2—3年可上市,但需要4—5年才能初次性成熟。哲羅鮭在養殖過程中雌、雄魚生長差異不大, 但在人工繁殖過程中1尾雄魚的精液可以配多尾雌魚, 因此雌、雄魚的比例為2∶1就可以滿足繁殖需求, 養殖過多的雄性親魚造成不必要的浪費。同時哲羅鮭雌魚懷卵量大, 可用來生產魚籽醬, 單性別養殖具有重大意義。本研究建立的哲羅鮭遺傳性別鑒定方法, 能夠很好地解決生產需求, 可在養殖場簡易的實驗條件下, 幼魚期采集少量鰭條進行無損傷的早期性別鑒定, 實現有計劃有目的養殖, 使養殖者獲得最大的效益。本方法在哲羅鮭育種方面也具有很大的應用價值, 通過此方法可實現哲羅鮭的單性養殖及性別控制育種, 為哲羅鮭的全雌或全雄育種奠定基礎。
