飼料糖和脂水平對團頭魴生長性能及血漿代謝物的影響
2021-07-17 03:25:10李紅燕巫麗云許文婕金俊琰楊云霞朱曉鳴劉昊昆解綬啟
水生生物學報 2021年4期
李紅燕 巫麗云 董 博 許文婕 金俊琰 楊云霞 朱曉鳴 韓 冬 劉昊昆 解綬啟
(1. 中國科學院水生生物研究所淡水生態(tài)與生物技術國家重點實驗室, 武漢 430072;2. 中國科學院大學現(xiàn)代農(nóng)業(yè)科學學院, 北京 100049)
飼料蛋白質是維持魚體組織生長和生命功能的重要物質, 也是價格最高的飼料原料[1]。在所有的蛋白源中, 魚粉因其氨基酸組成平衡, 可消化性強和含有較少的抗營養(yǎng)因子, 被認為是水產(chǎn)飼料的優(yōu)質蛋白源[2]。然而魚粉的生產(chǎn)依賴于海洋魚類資源, 水產(chǎn)飼料行業(yè)的可持續(xù)性發(fā)展需要進一步降低海洋資源的使用[3], 因此增加飼料中非蛋白能源物質的比例是目前研究的熱點。糖類和脂肪作為重要的非蛋白能源物質, 被認為具有節(jié)約蛋白的作用,在水產(chǎn)行業(yè)領域中的重要性與日俱增[4,5]。
糖類來源廣泛、價格低廉, 是飼料的主要能量來源[5,6]。糖類有利于飼料的制粒加工, 減少蛋白質作為能量的消耗, 并且可以為許多生物合成過程提供代謝底物[6,7]。然而, 飼料糖類含量過高則會導致許多的負面效應, 如生長受阻、脂肪的異常沉積和免疫機能下降等[8—10]。飼料脂肪為魚類生長發(fā)育提供能量, 同時為機體提供必需脂肪酸和一些脂溶性維生素, 對于魚類生命活動具有重要意義[11,12]。脂肪可以被大多數(shù)魚類較好的利用, 但飼料脂肪含量過剩或短缺都會對魚類生長和免疫造成直接的影響[13,14]。此外, 糖類和脂肪利用相互影響, 密不可分。脂類物質可以通過糖異生途徑轉化為葡萄糖, 糖也可以為脂肪合成提供底物從而轉化為脂肪儲存在組織中[15]。因此, 飼料中適宜的糖和脂肪水平對水產(chǎn)養(yǎng)殖的發(fā)展具有重要意義。在飼料糖和脂相對水平對魚類影響的研究中, 飼料糖脂比在不同種類如長吻(Leiocassis longirostrisGünther)[16]、黃顙魚(Pelteobagrus fulvidraco)[17]、大菱鲆(Scophthalmus maximus)[18]、草魚(Ctenopharyngodon idella)[8]和羅非魚(Oreochromis niloticus)[19]等魚類中均有開展。以上研究內容旨在探究不同魚類生長或生理狀態(tài)最佳條件下的最適糖脂比, 其確定可為飼料配比提供理論依據(jù)。
團頭魴(Megalobrama amblycephala)是我國重要的草食性淡水魚類, 具有生長快速、肉質鮮美和存活率高等優(yōu)點[20,21]。團頭魴在我國消費需求量大, 2019年的年產(chǎn)量已達78.32×107kg[22]。關于團頭魴糖和脂需求方面的研究較多, 研究結果表明,團頭魴幼魚最適飼料糖和脂需求分別為31%—34%和8%[23,24]。但相關研究較多為單一性的不同含量或不同來源的糖或脂肪的研究[23—27]。有關團頭魴糖脂比的研究均是按照糖/脂的比值按一定的比例逐漸增加, 來探究最適生長或生理狀態(tài)的糖/脂比值。研究表明高糖飼料促進了肝臟糖酵解過程, 在以特定生長率為評判指標時, 團頭魴最適糖脂比為3.58(糖和脂含量分別為291.70和81.40 g/kg)[28]。在以應激和抗氧化反應為評判指標時, 糖脂比在5.64(糖和脂含量分別為320.87和58.30 g/kg)時血漿皮質醇和乳酸等應激生物標志物含量最低[9]。但是文章中并沒有對脂肪代謝及糖和脂肪的交互作用進行研究。本研究以適合的糖和脂含量(中糖中脂)為對照, 探究在高糖低脂和低糖高脂這兩種相差較大的糖和脂的配比模式下, 糖和脂交互作用對團頭魴生長性能、飼料利用、營養(yǎng)沉積及相關生理代謝物的影響。
1 材料與方法
1.1 飼料配方與制作
本實驗制備了3種等氮飼料(蛋白質含量32%),分別包含不同糖和脂水平: 高糖低脂(HCLL, 45%糖和2%脂)、中糖中脂(MCML, 30%糖和8%脂)及低糖高脂(LCHL, 15%糖和14%脂)飼料。實驗飼料以玉米淀粉為碳水化合物源, 魚油和豆油等量混合為脂肪源, 飼料配方如表1所示。對所有飼料原料都進行研磨, 均勻混合后過40目篩網(wǎng)后加水調制,用制粒機(SLP-45, 漁業(yè)機械設備研究所, 中國上海)制備成飼料顆粒。以上各顆粒飼料于烘箱50℃烘干后于4℃冰柜中保存?zhèn)溆谩F渲? 飼料的脂肪酸組成見表2。
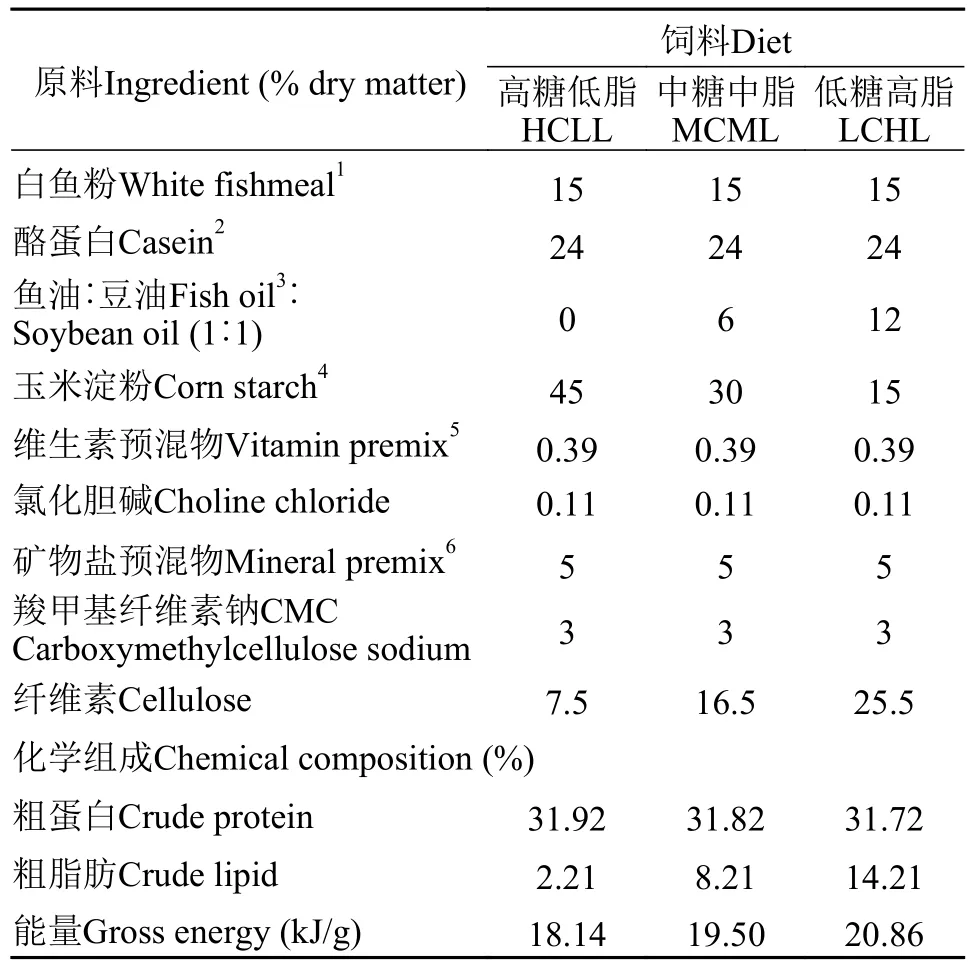
表1 飼料配方和化學組成(% 風干重)Tab. 1 Formulation and chemical composition of experimental diets
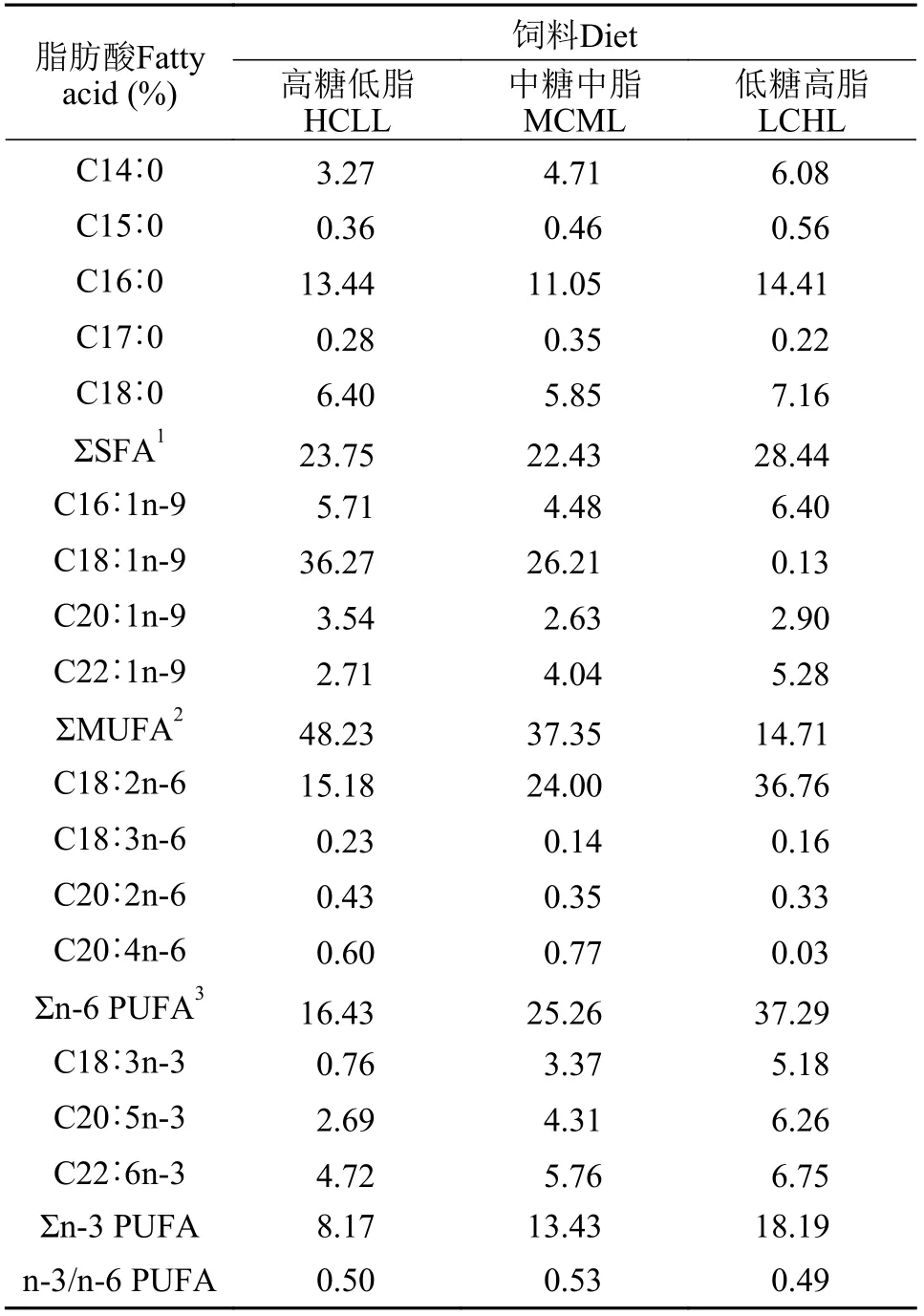
表2 飼料脂肪酸組成(% 總脂肪酸)Tab. 2 Fatty acid composition of the experimental diets (% of total fatty acids)
1.2 飼養(yǎng)管理
實驗所用團頭魴幼魚來自中國科學院水生生物研究所(中國武漢)。在正式養(yǎng)殖實驗前, 將制備的3種飼料等量均勻混合投喂進行為期2周的馴養(yǎng)以適應實驗系統(tǒng)環(huán)境。實驗開始前對實驗魚饑餓24h后撈出, 隨機挑選規(guī)格均勻和體質健康的團頭魴幼魚[初重(6.20±0.01) g]分為3個處理組。其中每缸30尾魚, 每個處理組3個平行。實驗在9個圓柱形玻璃纖維缸(水體積為167 L)組成的室內循環(huán)水系統(tǒng)中進行, 生長實驗持續(xù)56d, 每天飽食投喂3次(08:30、13:30和18:30)并記錄攝食量和殘餌。實驗期間每天進行水質監(jiān)測管理, 其中水溫為25—27℃,溶氧(7.09±0.12) mg/L, 氨氮(0.18±0.02) mg/L, pH為6.5—7.0。光照強度為2.21—2.95 μmol/(s·m2)。光照周期為12L∶12D(8:00—20:00光亮)。
1.3 樣品采集
在養(yǎng)殖實驗結束后, 在最后一次投喂后8h進行取樣。實驗魚用麻醉劑MS-222 (60 mg/L, Sigma,美國) 麻醉后稱總重后取樣。每缸4尾魚稱重后保存于–20℃冰箱中用于體成分分析。每缸3尾魚進行體長和體重測量, 并解剖肝臟和內臟團稱重以計算肝體比和臟體比。另外, 每缸隨機取2尾魚, 用肝素鈉抗凝劑潤過的無菌注射器進行尾部靜脈采血。血液3000×g10min離心后得到血漿, 保存于–80℃。同時, 在冰上取出肝臟組織, 置于液氮中速凍并于–80℃保存。
1.4 樣品分析
本實驗所有飼料和魚樣樣品基本組成的測定均參照AOAC (2003) 標準方法[29]進行檢測。干物質通過在烘箱(電熱恒溫干燥箱, 精宏, 中國上海)中105℃烘干至恒重, 通過失重法測定; 灰分在馬弗爐(馬弗爐, 中國湖北)中550℃煅燒3h, 采用失重法測定; 粗蛋白采用凱氏定氮儀(2300, Kjeltec Analyzer Unit)測定; 粗脂肪采用索氏抽提儀(Soxtec System HT6, Tecator, Hoganas, Sweden)進行抽提測定; 飼料脂肪酸含量參照Folch等[30]方法采取氯仿甲醇 =2∶1混合試劑提取總脂, 進行脂肪酸甲酯化處理后采用氣相色譜儀測定脂肪酸組成和含量 (7890A-5975C, Agilents Technologies Inc., Santa Clara, CA,USA)。
肝糖原含量根據(jù)南京建成生物工程研究所糖原測試盒(A043-1-1)說明書進行, 該方法根據(jù)蒽酮法原理, 利用糖原在濃硫酸作用下可脫水生成醛糖衍生物, 后者再與蒽酮作用形成藍色復合物, 與同法處理的標準葡萄糖溶液比色定量測得。肝臟甘油三酯(TG)含量采用GPO-PAP方法, 由南京建成采用商品試劑盒(A100-1-1)檢測(南京建成生物工程研究所, 中國南京)。
血漿葡萄糖、甘油三酯、游離脂肪酸(NEFA)和膽固醇含量采用商品試劑盒進行測定(Fujifilm,Wako Pure Chemical, Corporation, Osaka, Japan)。血漿低密度脂蛋白膽固醇(LDL-C)和高密度脂蛋白膽固醇(HDL-C)分別采用南京建成商品試劑盒A113-1-1和A112-1-1檢測(南京建成生物工程研究所,中國南京)。
1.5 數(shù)據(jù)統(tǒng)計分析
本實驗所有數(shù)據(jù)使用統(tǒng)計軟件SPSS 18.0進行統(tǒng)計分析。實驗結果采用均值±標準誤(Mean±SEM,n=6), 進行單因素方差分析(One-way ANOVA)。若各實驗組間差異顯著, 則同質性檢驗后進行Duncan’s多重比較。以P<0.05表示統(tǒng)計學差異顯著。
2 結果
2.1 生長與飼料利用
如表3所示, 攝食不同糖和脂水平飼料, 團頭魴幼魚攝食率FR各組間無顯著性差異(P>0.05), 但魚均攝食量FI在HCLL組顯著低于MCML組(P<0.05)。團頭魴攝食HCLL飼料組特定生長率SGR顯著低于MCML和LCHL組(P<0.05)。團頭魴飼料效率隨飼料糖水平減少脂水平增多而升高, 在高糖低脂HCLL組最低。攝食不同糖和脂水平飼料對團頭魴肥滿度CF、肝體比HSI、臟體比VSI均無顯著性差異(P>0.05)。
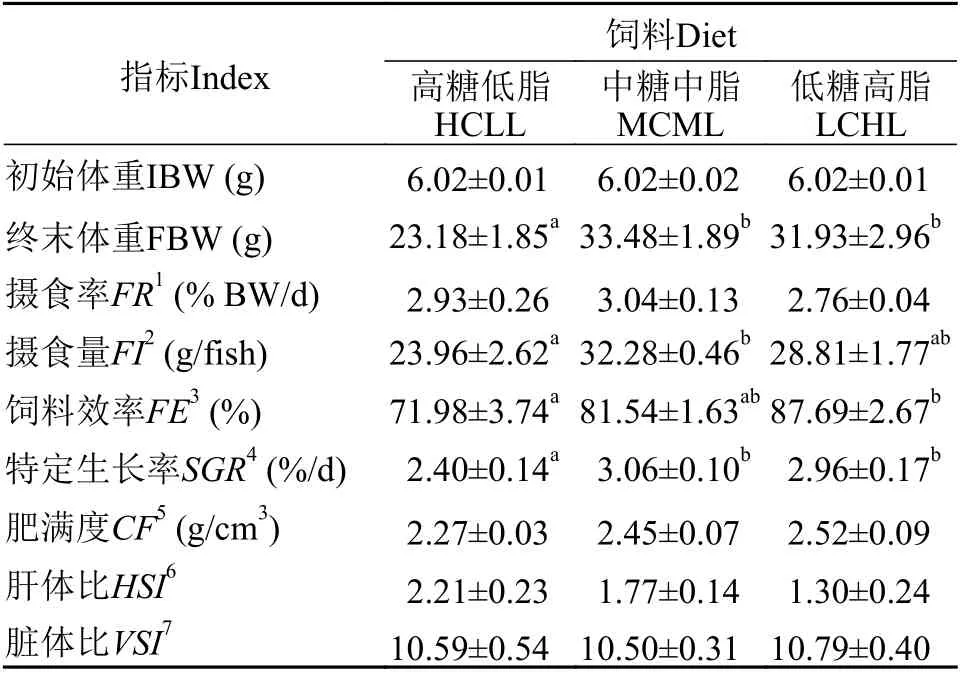
表3 不同飼料糖和脂水平對團頭魴幼魚生長性能、飼料利用和形體指數(shù)的影響Tab. 3 Effects of dietary carbohydrate and lipid levels on growth performance, feed utilization, and morphological indices in blunt snout bream juveniles
2.2 魚體組成與營養(yǎng)沉積
如表4所示, 團頭魴幼魚攝食不同糖和脂水平飼料后, 其全魚粗蛋白、粗脂肪、水分及灰分含量均無顯著性差異(P>0.05)。隨飼料糖含量增加脂含量減少, 魚體蛋白沉積率降低, 反之, 脂肪沉積率增加。
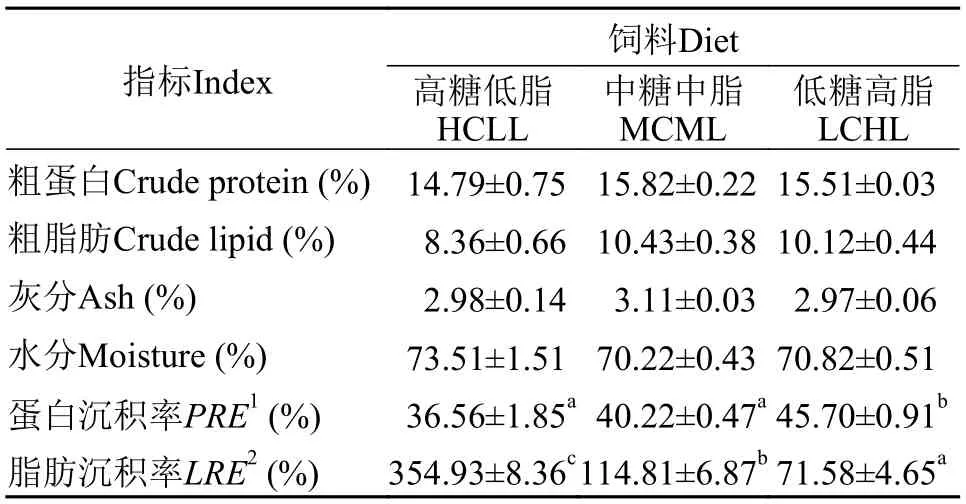
表4 不同飼料糖和脂水平對團頭魴幼魚體生化組成和營養(yǎng)沉積的影響Tab. 4 Effects of dietary carbohydrate and lipid levels on body chemical composition and nutrient retention efficiency in blunt snout bream juveniles
2.3 肝臟糖原和甘油三酯含量
如圖1所示, 不同糖和脂水平飼料組間, 團頭魴肝糖原和甘油三酯含量均無顯著性差異(P>0.05)。
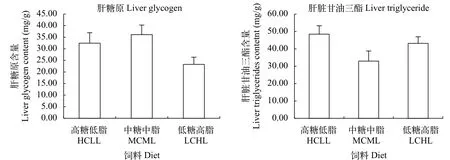
圖1 不同飼料糖和脂水平對團頭魴肝臟糖原含量和甘油三酯含量的影響Fig. 1 Effects of dietary carbohydrate and lipid levels on glycogen and triglycerides content in the liver of blunt snout bream
2.4 血漿代謝物水平
如表5所示, 血漿葡萄糖水平、甘油三酯、游離脂肪酸、總膽固醇及高密度脂蛋白膽固醇含量在各飼料組間均無顯著性差異(P>0.05)。血漿低密度脂蛋白膽固醇含量在LCHL組顯著低于其他飼料組(P<0.05)。
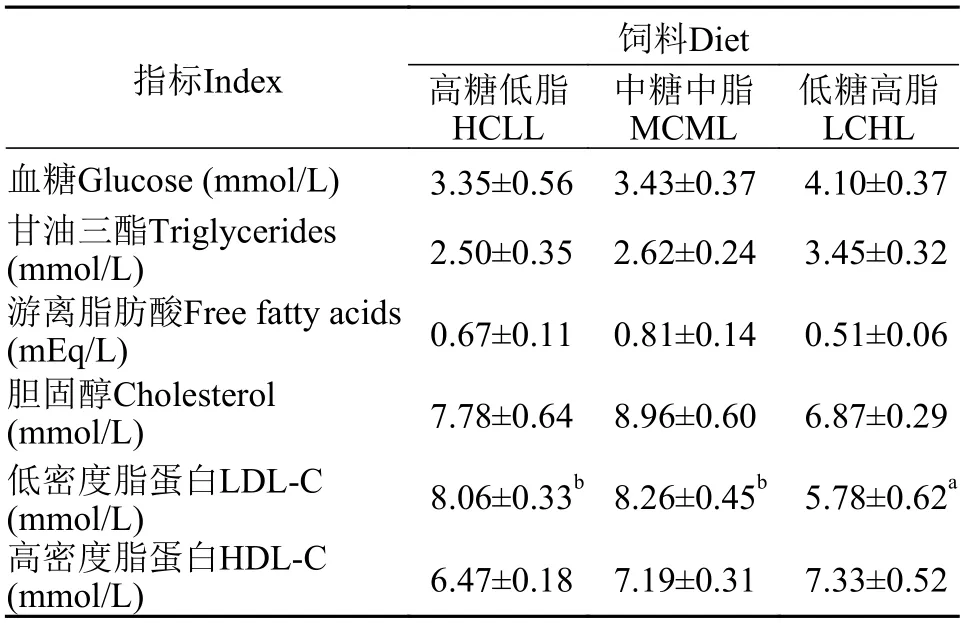
表5 不同飼料糖和脂水平對團頭魴幼血漿生化組成的影響Tab. 5 Effects of dietary carbohydrate and lipid levels on plasma metabolites in blunt snout bream juveniles
3 討論
3.1 飼料糖和脂水平對團頭魴生長和飼料利用的影響
飼料中非蛋白能源物質糖和脂肪的適宜添加水平對于維持魚類生長和生命過程十分重要, 但過高或過低的添加水平都會對魚體造成負面的生理響應[6,8,31]。高糖飼料導致的生長抑制效應在多種魚類中已經(jīng)得到證實, 如虹鱒(Salmo gairdneri)[32]、大西洋鮭(Salmo salarL.)[33]和草魚(Ctenopharngon idellus)[34]等。在團頭魴糖脂比的研究中, 團頭魴攝食量隨飼料糖脂比升高(CHO/L從1.62到10.46)而增多, 但更高的糖脂比(CHO/L: 24.20)卻導致攝食量的顯著下降, 從而抑制了團頭魴的生長[28]。與此一致, 在本實驗中團頭魴高糖低脂HCLL組(CHO/L:22.5)魚均攝食量FI顯著低于適宜糖脂中糖中脂MCML組, 且高糖低脂HCLL飼料顯著降低了團頭魴幼魚的飼料利用效率, 造成特定生長率下降。由此可見, 高糖造成了團頭魴生長性能受到抑制的負面效應。另外, 研究表明飼料高小麥淀粉水平造成草魚對蛋白和糖的表觀消化率顯著低于低淀粉水平組[34], 表明高糖可能造成魚類對營養(yǎng)物質消吸收的減弱, 也可能是造成生長抑制的原因之一。然而,魚類對飼料中糖和脂肪的利用受到兩者水平的相互影響, 飼料的脂肪水平也會影響糖的利用[15,35],前期已有研究驗證了團頭魴對于糖和脂的利用存在交互影響[36]。并且, 飼料脂肪含量過低可能不足以為魚體提供生長必需的能量和某些生物合成過程所需的脂肪酸, 進而導致魚體的代謝紊亂。在本實驗中, 3種糖脂水平飼料的脂肪源均采用魚油和豆油等比例混合, 脂肪酸種類均衡(表2), 滿足團頭魴對于脂肪酸種類的需求。且已有研究報道團頭魴的最適脂肪需求為8%[23,24], 在本實驗中2%脂肪水平的高糖低脂HCLL組團頭魴攝食量FI和相應的總脂肪攝入量均低于適宜糖脂即中糖中脂MCML組, 不足以維持團頭魴生長所必需的脂肪需求。綜上, 高糖和低脂的交互作用因素導致團頭魴攝食量FI和飼料效率FE的降低, 從而抑制了團頭魴的生長性能。另外, 不同魚類利用糖和脂供能的效率可能不同, 如虹鱒[37]和歐洲海鱸(Dicentrarchus labraxL.)[38]對飼料脂肪的利用優(yōu)于糖類, 而尼羅羅非魚(Oreochromis niloticus)[39]和非洲鯰(Clarias gariepinus)[40]則相反。在本實驗3個飼料組中, 隨著飼料糖水平降低、脂水平升高, 團頭魴的飼料效率呈升高趨勢。這表明團頭魴對脂肪有著更好的利用, 從某種程度上來說其對高糖缺乏耐受[24,28]。另外, 中糖中脂組特定生長率最高, 表明適宜水平的糖和脂的配伍才可以達到團頭魴生長性能的最大化, 同時也驗證了魚類飼料糖和脂等營養(yǎng)物質最適需求研究的必要性。
3.2 高糖低脂未造成團頭魴肝臟脂肪異常沉積或形體指數(shù)的改變
一般來說, 攝入過量的碳水化合物, 其消化產(chǎn)生的單糖會通過糖酵解途徑轉化為丙酮酸, 進而通過氧化為機體供能或者合成脂肪在機體中儲存起來[41]。因此, 高糖飼料通常造成許多魚類肝體比增加, 肝臟脂肪代謝的紊亂甚至是脂肪肝的生成[8,42]。在本實驗中, 3種糖和脂水平飼料組間團頭魴幼魚肝糖原含量和甘油三酯含量無顯著性差異, 且肝體比各組間也無顯著性差異。因此, 高糖低脂飼料并未造成團頭魴肝臟脂肪或糖原的異常沉積或脂肪肝的產(chǎn)生。與本實驗結果一致, 在團頭魴不同糖脂比的研究中, 飼料糖脂比24.2時即糖41.6%、脂1.7%含量時肝體比HSI和肝臟脂肪含量與其他組間無顯著性差異[28]。這一結果的產(chǎn)生可能歸因于以下幾點: 首先, 肝臟不是團頭魴脂肪沉積的主要場所[28,36];其次, 在劉梅珍等[43]對團頭魴的研究中, 2%—11%的不同脂肪水平飼料都未誘發(fā)團頭魴的肝臟脂肪沉積, 所以本實驗高糖低脂HCLL飼料中2%的脂肪含量對于誘發(fā)團頭魴的脂肪沉積較低。綜合以上因素, 高糖低脂HCLL飼料對團頭魴形體指數(shù)、肝糖原和脂肪沉積無顯著性影響。
3.3 飼料糖和脂水平與團頭魴魚體營養(yǎng)沉積
在一些魚類中, 飼料糖水平可能比脂水平有著更重要的脂肪合成誘因效果, 高糖飼料可以促進脂肪合成相關酶活性[8,42]。并且, 在某些魚類中魚體脂肪含量隨飼料碳水化合物水平升高而升高[38]。在本實驗中, 各飼料組團頭魴體粗脂肪含量無顯著性差異。但高糖低脂HCLL組脂肪沉積率LRE顯著高于其他兩飼料組, 表明機體內存在由糖向脂的轉化過程。而表觀上來講, 較高的脂肪沉積率卻未造成魚體脂肪生化組成高于其他兩飼料組, 這一現(xiàn)象產(chǎn)生的原因可能是高糖低脂HCLL組食源性脂肪攝入量少于其他兩飼料組, 高糖轉化為的脂肪用于機體生長而未在魚體內過度沉積。同時, 脂肪作為重要的能源物質, 被認為具有節(jié)約蛋白的作用[4]。隨著飼料脂肪含量升高, 團頭魴更多的利用脂肪來為機體供能, 從而減少了蛋白作為能量物質用于生長活動的消耗, 促進了低糖高脂組的蛋白沉積。與之一致的是, 團頭魴氮沉積率隨飼料糖脂比增加(CHO/L: 2.45—24.0)而減少[28]。由此可知, 盡管團頭魴作為草食性魚類, 但隨飼料脂肪增多可以更好地儲存蛋白質, 表明團頭魴對脂肪的利用較好。
3.4 團頭魴脂質轉運與脂質穩(wěn)態(tài)調節(jié)
肝臟是魚類中間代謝的主要器官, 且肝臟脂穩(wěn)態(tài)是一個復雜的過程, 包括脂的分泌、轉運和攝取等過程的綜合作用[13]。血漿生化指標可以作為機體代謝和生理狀態(tài)的反應, 如甘油三酯、膽固醇和脂蛋白等脂類物質水平可以反映機體對于脂質的動員情況[44,45]。在本實驗中, 團頭魴血漿葡萄糖、甘油三酯、游離脂肪酸和膽固醇的含量各飼料組間無顯著性差異, 表明團頭魴在不同糖和脂水平飼料攝入下可以維持血漿生理的穩(wěn)態(tài)。與哺乳動物類似, 魚類的脂類轉運通過脂蛋白經(jīng)內源性和外源性循環(huán)來調控。其中內源性途徑是指肝臟中極低密度脂蛋白(VLDL)向外分泌脂類并轉化為中間密度脂蛋白(IDL)和低密度脂蛋白, 而高密度脂蛋白負責脂質的逆向轉運[46]。攝食高脂飼料造成草魚[47]、大黃魚 (Larimichthys crocea)[46]和紅鰭東方鲀 (Takifugu rubripes)[48]等魚血漿低密度脂蛋白的顯著增高。在本實驗中低密度脂蛋白膽固醇在低糖高脂組顯著增加, 表明團頭魴通過低密度脂蛋白將較多攝入的脂質轉移到外周組織從而維持肝臟脂質的穩(wěn)態(tài)。
綜上所述, 團頭魴在飼料低糖高脂配比時生長性能、飼料利用和蛋白沉積均顯著優(yōu)于低糖高脂條件, 表明雖然是草食性魚類, 但團頭魴對脂肪的利用仍優(yōu)于糖類。且在飼料糖和脂的相對配比下,團頭魴血漿生理穩(wěn)態(tài), 未造成肝臟脂肪的異常沉積,肝體指數(shù)的增加或體脂肪的異常改變, 表明團頭魴具有較好的應對飼料糖和脂水平變化的自身代謝調控機制。
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
小讀者(2021年2期)2021-03-29 05:03:48
華人時刊(2019年13期)2019-11-17 14:59:54
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
人大建設(2019年12期)2019-05-21 02:55:32
文苑(2018年22期)2018-11-19 02:54:14
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
發(fā)明與創(chuàng)新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
