不同開(kāi)口餌料對(duì)黑斑原仔魚(yú)生長(zhǎng)和消化酶活性的影響
2021-07-24 07:49:50王萬(wàn)良王金林曾本和周建設(shè)
水產(chǎn)學(xué)雜志 2021年3期
關(guān)鍵詞:差異
王萬(wàn)良,王金林,曾本和,周建設(shè)
(西藏自治區(qū)農(nóng)牧科學(xué)院水產(chǎn)科學(xué)研究所,西藏 拉薩 850000)
1 材料與方法
1.1 材料
1.2 方法
實(shí)驗(yàn)在平列槽中進(jìn)行,每個(gè)平列槽(280 cm×50 cm×25 cm)用網(wǎng)一分為三,隔離規(guī)格為60 cm×50 cm×25 cm,確保環(huán)境一致,共設(shè)置11 個(gè)處理,每個(gè)處理3 個(gè)平行,每個(gè)平行仔魚(yú)300 尾,共計(jì)9 900尾。每個(gè)處理分別投喂微粒子飼料(Ⅰ組)、冰凍搖蚊幼蟲(chóng)(Ⅱ組)、攪碎的豬肝(Ⅲ組)、微粒子飼料+螺旋藻(9∶1)(Ⅳ組)、搖蚊幼蟲(chóng)+螺旋藻(9∶1)(Ⅴ組)、豬肝+螺旋藻(9∶1)(Ⅵ組)、微粒子飼料+螺旋藻+搖蚊幼蟲(chóng)(4∶1∶5)(Ⅶ組)、微粒子飼料+螺旋藻+豬肝(4∶1∶5)(Ⅷ組)、微粒子飼料+螺旋藻+輪蟲(chóng)(4∶1∶5)(Ⅸ組)、微粒子飼料+蒼蠅幼蟲(chóng)(1∶1)(Ⅹ組)和人工配合飼料(Ⅺ組)。
表1 黑斑原魚(yú)苗人工配合飼料配方Tab.1 Formula of artificial compound diet for Glyptosternum maculatum
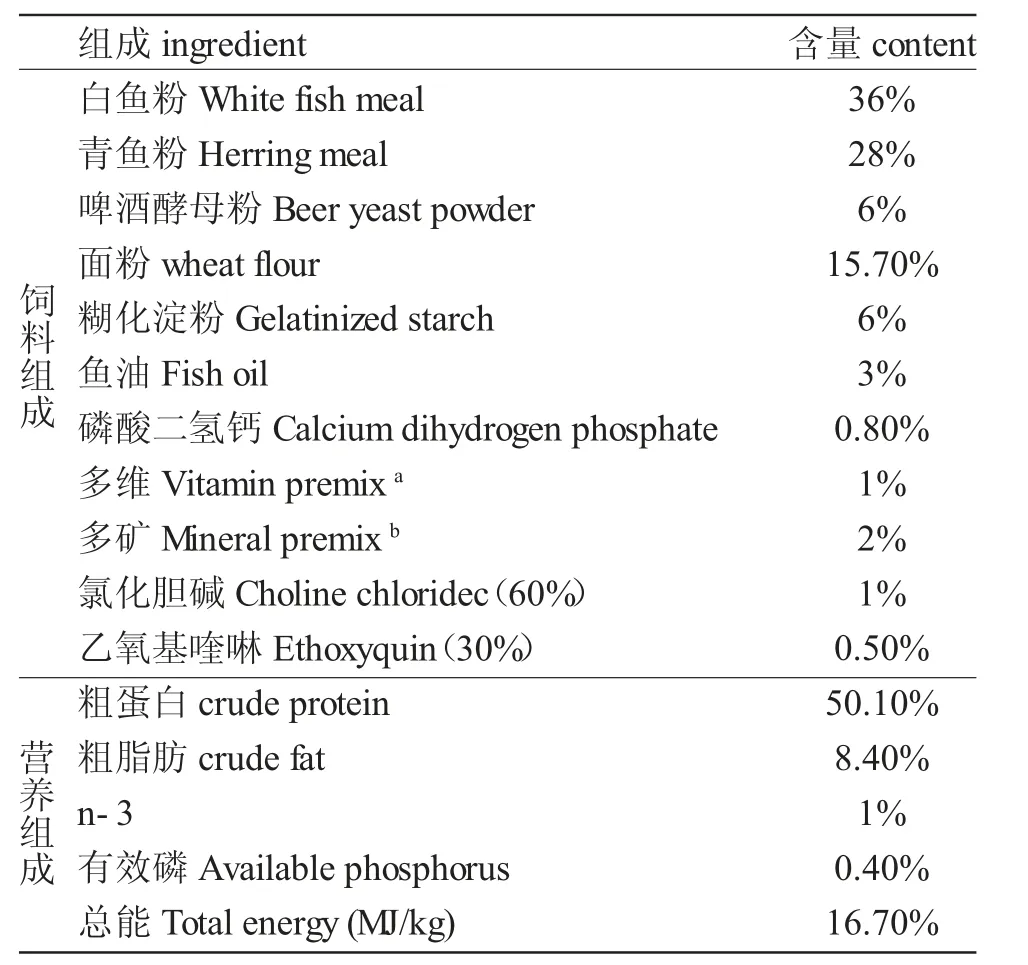
表1 黑斑原魚(yú)苗人工配合飼料配方Tab.1 Formula of artificial compound diet for Glyptosternum maculatum
注:飼料營(yíng)養(yǎng)成分為實(shí)測(cè)值a.多維中(mg/kg 飼料):鹽酸硫胺素(98%)25,維生素B2(80%)45,鹽酸吡哆醇(99%)20,維生素B12(1%)10,維生素K(51%)10,肌醇(98%)800,泛酸鈣(98%)60,煙酸(99%)200,葉酸(98%)20,生物素(2%)60,維生素A(500 000 IU/g)32,維生素D(500 000 IU/g)5,維生素E(50%)240,維生素C(35%)2000,抗氧化劑(克氧靈,100%)3,稻殼粉1470.2;b.多礦中(mg/kg 飼料):CoCl2·6H2O(1%)50,CuSO4·5H2O(25%)10,F(xiàn)eSO4·H2O(30%)80,ZnSO4·H2O(34.5%)50,MnSO4·H2O(31.8%)45,MgSO4·7H2O(15%)1200,亞硒酸鈉(1%)20,碘酸鈣(1%)60,沸石粉8485Notes:The diet nutrient contents are measured a.Vitamin premix provides(mg/kg diet):Thiamin(98%)25,Riboflavin(80%)45,Pyridoxine-HCl(99%)20,Vitamin B12(1%)10,VitaminK(51%)10,Inositol(98%)800,Pantothenicacid(98%)60,Niacin acid(99%)200,Folic acid(98%)20,Biotin(2%)60,Retinolacetate(500,000 IU/g)32,Cholecalciferol(500,000 IU/g)5,Alphatocopherol(50%)240,Ascorbic acid(35%)2000,Anti-oxidants(Oxygenlinggrams,100%)3,and ricehuskpowder(100%)1470.b.Mimeral premix provides(mg/kg diet):CoCl2·6H2O(1%)50,CuSO4·5H2O(25%)10,FeSO4·H2O(30%)80,ZnSO4·H2O(34.5%)50,MnSO4·H2O(31.8%)45,MgSO4·7H2O(15%)1200,Sodium selenite(1%)20,Calcium iodine(1%)60,and Zoelite 8485
實(shí)驗(yàn)期間,保持水溫12~13℃,溶氧6 mg/L,每天下午5:50~6:00 投喂1 次,投喂時(shí)關(guān)閉進(jìn)水閥門(mén),每個(gè)平列槽用遮光板遮蓋,次日7:00 清污。每周下午13:00~14:00 之間定期在平列槽靜水條件下用1%鹽水浸泡10 min。
每天統(tǒng)計(jì)死亡仔魚(yú)尾數(shù)。每7 d 測(cè)定1 次生長(zhǎng),每個(gè)平行隨機(jī)選取5 尾仔魚(yú),經(jīng)麻醉后用游標(biāo)卡尺逐尾測(cè)量全長(zhǎng)(吻端至尾鰭末端),輕輕吸干體表水分后用萬(wàn)分之一天平稱(chēng)量體質(zhì)量,整個(gè)實(shí)驗(yàn)持續(xù)6周共42 d。死亡率(%)=死亡尾數(shù)/總尾數(shù)×100。由于仔魚(yú)個(gè)體較小,實(shí)驗(yàn)結(jié)束時(shí)不同實(shí)驗(yàn)組各取6尾,按照重量(g)體積(mL)比1∶9 的量加入預(yù)冷的0.65%生理鹽水,整魚(yú)剪碎后在4℃下以3 500 r/min離心10 min,取組織勻漿上清液,即為粗酶提取液,相關(guān)消化酶活力參照Engrola 等[12]方法測(cè)定。
數(shù)據(jù)采用SPSS17.0 軟件進(jìn)行統(tǒng)計(jì)分析,差異顯著性測(cè)驗(yàn)采用單因素方差分析(One-way ANOVA)和Duncan's 多重比較。
2 結(jié)果與分析
2.1 不同開(kāi)口餌料對(duì)黑斑原仔魚(yú)死亡率的影響
由表2 可知,養(yǎng)殖7 d 時(shí),Ⅱ組魚(yú)死亡率最低,顯著低于Ⅵ組(P<0.05),與其他各組差異不顯著(P>0.05);養(yǎng)殖14 d 時(shí),Ⅸ組魚(yú)死亡率最高,顯著高于Ⅱ組、Ⅲ組、Ⅳ組、Ⅴ組、Ⅶ組和Ⅺ組(P<0.05),與其他各組差異不顯著(P>0.05);養(yǎng)殖21 d 時(shí),Ⅹ組魚(yú)死亡率最高,與Ⅰ組、Ⅲ組、Ⅵ組、Ⅷ組、Ⅸ組、Ⅺ組差異不顯著(P>0.05),顯著高于其他各組(P<0.05);養(yǎng)殖28 d 時(shí),Ⅵ組魚(yú)死亡率最高,與Ⅷ差異不顯著(P>0.05),顯著高于其他各組(P<0.05);養(yǎng)殖35 d 時(shí),Ⅷ組魚(yú)死亡率最高,顯著高于其他各組;養(yǎng)殖42 d 時(shí),Ⅱ組魚(yú)死亡率最低,與Ⅴ組差異不顯著(P>0.05),但顯著低于其他各組(P<0.05),Ⅵ組死亡率最高,顯著高于其他各組(P<0.05)。試驗(yàn)最終死亡率由低到高依次為Ⅱ組<Ⅴ組<Ⅳ組<Ⅰ組<Ⅶ組<Ⅹ組<Ⅸ<Ⅺ組<Ⅷ組<Ⅲ組<Ⅵ組,即搖蚊幼蟲(chóng)<(搖蚊幼蟲(chóng)+螺旋藻)<(微粒子飼料+螺旋藻)<微粒子飼料<(微粒子飼料+螺旋藻+搖蚊幼蟲(chóng))<(微粒子飼料+蒼蠅幼蟲(chóng))<(微粒子飼料+螺旋藻+輪蟲(chóng))<人工配合飼料<(微粒子飼料+螺旋藻+豬肝)<豬肝<(豬肝+螺藻)。
表2 不同開(kāi)口餌料對(duì)黑斑原仔魚(yú)死亡率的影響Tab.2 The effect of different weaning diets on the mortality of larval Glyptosternum maculatum
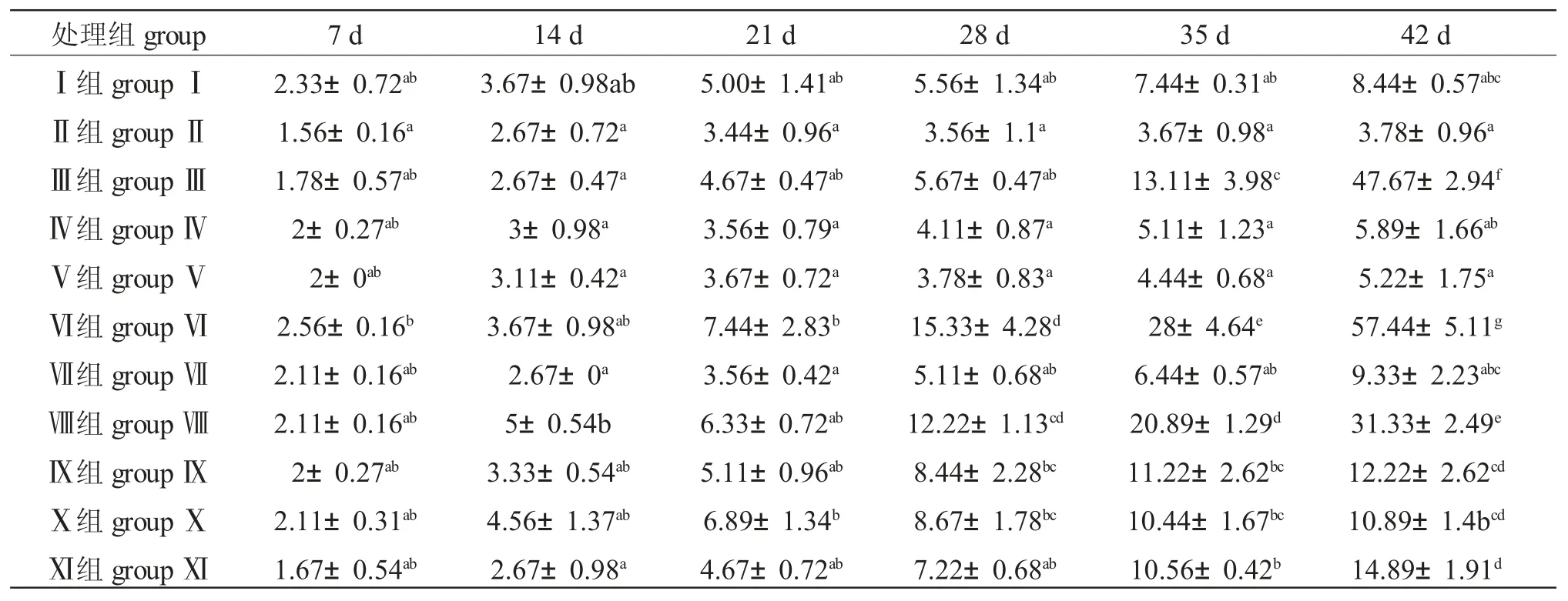
表2 不同開(kāi)口餌料對(duì)黑斑原仔魚(yú)死亡率的影響Tab.2 The effect of different weaning diets on the mortality of larval Glyptosternum maculatum
注:同列中標(biāo)有不同小寫(xiě)字母者表示組間差異顯著(P<0.05),標(biāo)有相同小寫(xiě)字母者表示組間差異不顯著(P>0.05),下同Note:means with different letters in the same column are significant differences among the groups(P <0.05),and the means with the same letter in the same column are not significant differences among the groups(P >0.05),et sequentia
2.2 不同開(kāi)口餌料對(duì)黑斑原體質(zhì)量的影響
由圖1 可知,開(kāi)口7 d、14 d 和21 d 時(shí),各試驗(yàn)組魚(yú)體質(zhì)量變化均呈遞增趨勢(shì)。開(kāi)口7 d 時(shí),Ⅹ組魚(yú)體質(zhì)量最快,顯著高于Ⅰ組和Ⅲ組(P<0.05),與其他各組差異不顯著(P>0.05);開(kāi)口14 d 時(shí),Ⅳ組魚(yú)體質(zhì)量增長(zhǎng)最慢,顯著低于Ⅴ組和Ⅹ組(P<0.05),與其他各組差異不顯著(P>0.05);開(kāi)口21 d 時(shí),Ⅴ組體質(zhì)量增長(zhǎng)最快,顯著高于Ⅲ組、Ⅵ組、Ⅷ組和Ⅺ組(P<0.05),與其他各組差異不顯著(P>0.05);開(kāi)口28 d 時(shí),Ⅲ組、Ⅵ組、Ⅷ組體質(zhì)量呈負(fù)增長(zhǎng),Ⅲ組增長(zhǎng)最慢,顯著低于其他各組(P<0.05);開(kāi)口35 d時(shí),Ⅱ組體質(zhì)量增長(zhǎng)最快,顯著高于其他各組(P<0.05),與Ⅴ組、Ⅶ組、Ⅹ組差異不顯著(P>0.05);開(kāi)口42 d 時(shí),Ⅶ組生長(zhǎng)最快,與Ⅱ組、Ⅴ組差異不顯著(P>0.05),但顯著高于其他各組(P<0.05)。
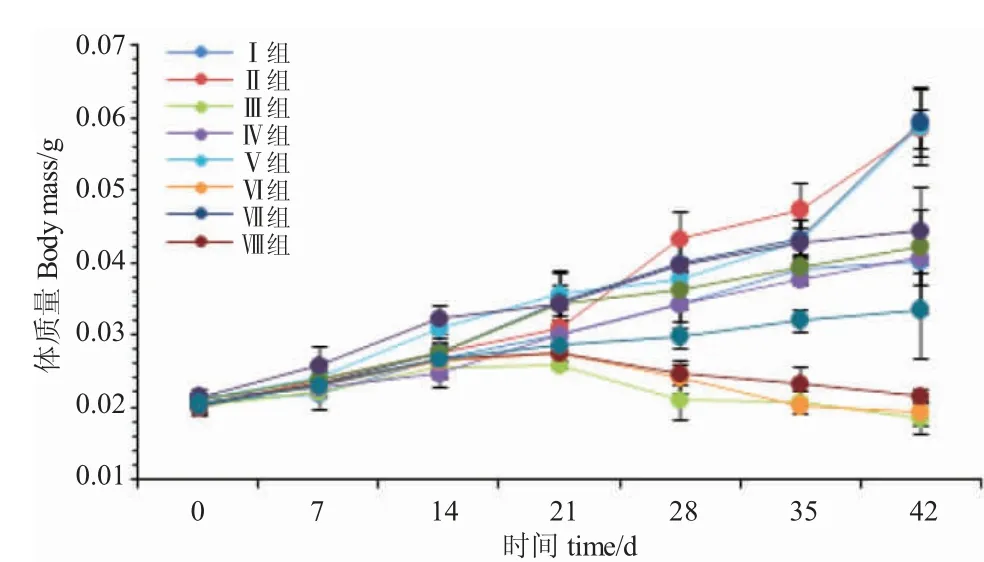
圖1 攝食不同開(kāi)口餌料的黑斑原仔魚(yú)體質(zhì)量的增長(zhǎng)Fig.1 Growth of body weight of the larval Glyptosternum maculatum fed different diets
2.3 不同開(kāi)口餌料對(duì)黑斑原全長(zhǎng)的影響
由圖2 可知,開(kāi)口7 d、14 d 和21 d 時(shí),各處理組魚(yú)的全長(zhǎng)均呈現(xiàn)遞增趨勢(shì)。開(kāi)口7 d 時(shí),Ⅰ組魚(yú)全長(zhǎng)增長(zhǎng)最慢,顯著低于其他各組(P<0.05),與其他各組間差異不顯著(P>0.05);開(kāi)口14 d 時(shí),Ⅴ組魚(yú)全長(zhǎng)增長(zhǎng)最快,與Ⅹ組差異不顯著(P>0.05),但顯著高于其他各組(P<0.05);開(kāi)口21 d 時(shí),Ⅰ組魚(yú)全長(zhǎng)增長(zhǎng)最慢,與Ⅲ組、Ⅷ組和Ⅹ組差異不顯著(P>0.05),顯著低于其他各組(P<0.05);開(kāi)口28 d 時(shí),Ⅲ組和Ⅵ組魚(yú)全長(zhǎng)開(kāi)始呈現(xiàn)負(fù)增長(zhǎng),二者差異顯著(P<0.05),其他各組均呈現(xiàn)正增長(zhǎng),Ⅳ組增長(zhǎng)最快,與Ⅱ組、Ⅴ組、Ⅶ組、Ⅷ組和Ⅹ組差異不顯著(P>0.05),但顯著高于Ⅰ組、Ⅺ組(P<0.05);開(kāi)口35 d時(shí),Ⅶ組魚(yú)全長(zhǎng)增長(zhǎng)最快,與Ⅰ組、Ⅱ組、Ⅳ組、Ⅴ組差異不顯著(P>0.05),顯著高于其他各組(P<0.05);開(kāi)口42 d 時(shí),Ⅶ組魚(yú)全長(zhǎng)增長(zhǎng)最快,與Ⅱ組、Ⅴ組差異不顯著(P>0.05),顯著高于其他各組(P<0.05),Ⅲ組魚(yú)全長(zhǎng)增長(zhǎng)最慢,顯著低于其他各組(P<0.05)。
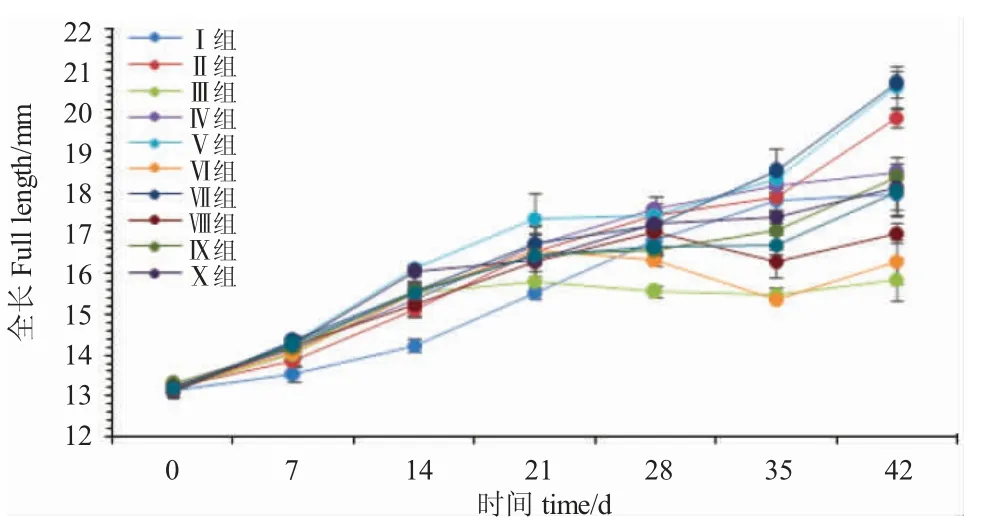
圖2 攝食不同開(kāi)口餌料的黑斑原仔魚(yú)全長(zhǎng)的增長(zhǎng)Fig.2 The growth of the larval Glyptosternum maculatum fed different weaning diets
2.4 不同開(kāi)口餌料對(duì)黑斑原仔魚(yú)消化酶活性的影響
表3 不同開(kāi)口餌料對(duì)黑斑原仔魚(yú)消化酶變化Tab.3 Changes of digestive enzymes of different diets to the larva of the Glyptosternum maculatum
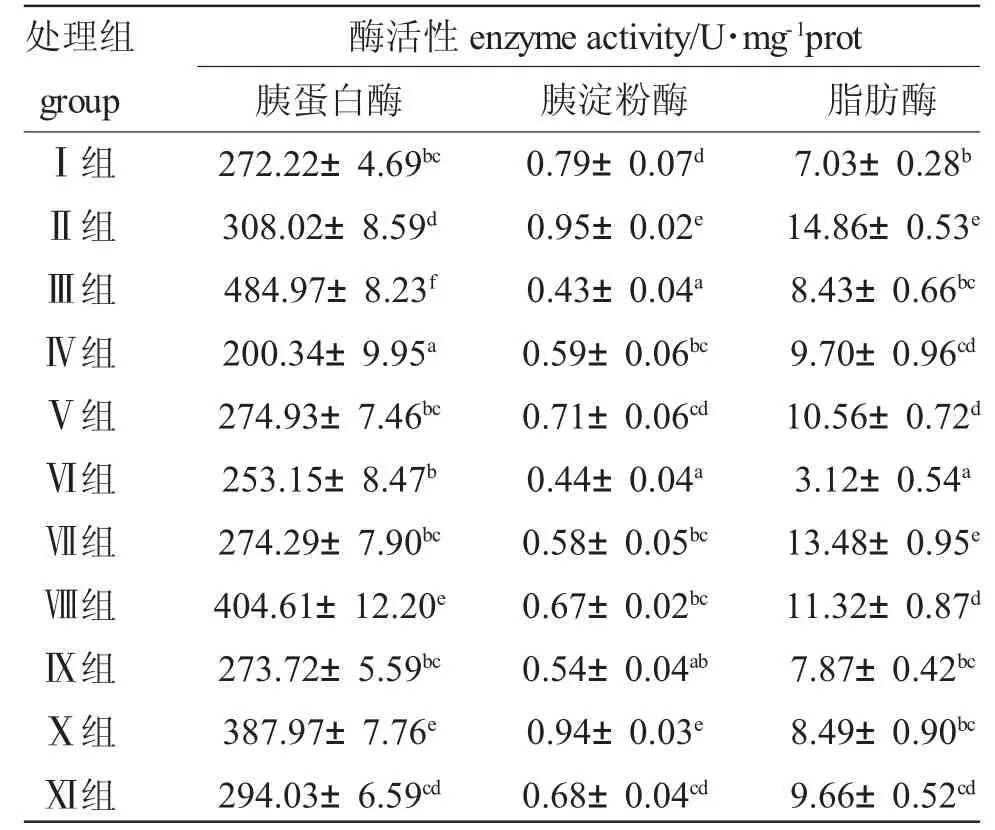
表3 不同開(kāi)口餌料對(duì)黑斑原仔魚(yú)消化酶變化Tab.3 Changes of digestive enzymes of different diets to the larva of the Glyptosternum maculatum
3 討論
3.1 不同開(kāi)口餌料對(duì)黑斑原仔魚(yú)存活和生長(zhǎng)的影響
早期生活期是魚(yú)類(lèi)在整個(gè)生命過(guò)程中對(duì)外界環(huán)境最敏感的時(shí)期,也是死亡率最高的重要階段[13,14]。開(kāi)口餌料對(duì)魚(yú)類(lèi)早期生活階段的存活和后期生長(zhǎng)具有直接的影響,與仔魚(yú)開(kāi)口攝食后,對(duì)餌料的種類(lèi)、大小、密度及營(yíng)養(yǎng)等密切相關(guān)。開(kāi)口餌料的選擇已成為魚(yú)類(lèi)苗種培育最主要的研究?jī)?nèi)容之一[15-18]。在自然水體中,黑斑原屬于雜食性魚(yú)類(lèi),口裂寬大、下位,具有發(fā)達(dá)的口腔齒,成魚(yú)主要以小型魚(yú)類(lèi)和藻類(lèi)為主。本研究結(jié)果表明,黑斑原仔魚(yú)對(duì)開(kāi)口餌料的選擇性較低,除了豬肝餌料組之外,不管是單一餌料,還是組合餌料對(duì)其能夠開(kāi)口投喂,經(jīng)過(guò)42 d 養(yǎng)殖死亡率均在15%以?xún)?nèi),且投喂搖蚊幼蟲(chóng)單一餌料組死亡率最低,僅為3.78%;從生長(zhǎng)來(lái)看,投喂組合餌料(微粒子飼料+螺旋藻+搖蚊幼蟲(chóng))終末體質(zhì)量和體全長(zhǎng)增長(zhǎng)均最快,死亡率為9.33%,可見(jiàn),就單一餌料而言,天然餌料進(jìn)行黑斑原仔魚(yú)開(kāi)口優(yōu)于微粒子飼料和人工配合飼料,這與羅非魚(yú)Oreochroms mossambcus[19]、納氏鱘Acipenser naccarii[20]、四川裂腹魚(yú)Schizothorax kozlovi[21]和半刺厚唇魚(yú)Acrossocheilius hemispinus[22]進(jìn)行開(kāi)口餌料研究所得的結(jié)果相一致。然而,豬肝不適宜作為黑斑原仔魚(yú)開(kāi)口餌料,主要原因可能是平列槽中的水體有限,攪碎的豬肝容易影響水體水質(zhì)和攝食,造成死亡率較高和生長(zhǎng)緩慢;而投喂(微粒子飼料+螺旋藻+搖蚊幼蟲(chóng))餌料生長(zhǎng)最快的主要原因可能是組合餌料營(yíng)養(yǎng)成分較為全面,對(duì)黑斑原仔魚(yú)生長(zhǎng)和存活均具有顯著促進(jìn)作用。
3.2 不同開(kāi)口餌料對(duì)黑斑原仔魚(yú)消化酶變化
通常,魚(yú)類(lèi)在早期發(fā)育過(guò)程中,消化系統(tǒng)具有一定的可塑性,消化酶活性的強(qiáng)弱對(duì)魚(yú)類(lèi)消化吸收具有重要的影響,與其早期存活和生長(zhǎng)關(guān)系很大,通過(guò)調(diào)節(jié)自身消化酶活性實(shí)現(xiàn)機(jī)體內(nèi)儲(chǔ)存營(yíng)養(yǎng)物質(zhì)的有效利用,維持正常的新陳代謝[22-24]。本次實(shí)驗(yàn)中,采用不同餌料對(duì)黑斑原進(jìn)行投喂,消化酶也產(chǎn)生了相應(yīng)的變化。
目前有關(guān)魚(yú)類(lèi)蛋白酶活性的研究結(jié)果尚不一致。有的認(rèn)為,肉食性魚(yú)類(lèi)蛋白酶活性高于草食性魚(yú)類(lèi),但鯉(雜食性)蛋白酶活性又高于鰻鱺(肉食性),甚至還有報(bào)道稱(chēng),魚(yú)類(lèi)蛋白酶活性差異跟食性的變化不明顯[25,26]。而本實(shí)驗(yàn)結(jié)果表明,不同餌料對(duì)黑斑原仔魚(yú)蛋白酶活性有顯著影響,投喂肉食性餌料消化酶遠(yuǎn)高于人工配合餌料。魚(yú)類(lèi)的各個(gè)器官均分布有淀粉酶和脂肪酶,通常認(rèn)為淀粉酶活性與魚(yú)類(lèi)的種類(lèi)存在差異較大,脂肪酶的活性與魚(yú)攝食食物脂肪含量呈正相關(guān)[27]。趙月月等[28]采用不同餌料研究了對(duì)稀有鯽Gobiocypris raru 仔稚魚(yú)的消化酶活性,結(jié)果淀粉酶和脂肪酶變化均無(wú)顯著性差異。然而本研究結(jié)果表明,投喂不同開(kāi)口餌料對(duì)黑斑原仔魚(yú)淀粉酶和脂肪酶變化存在顯著差異,搖蚊幼蟲(chóng)試驗(yàn)組胰淀粉酶活性和脂肪酶活性最高,與上述結(jié)果不一致。
3.3 小結(jié)
結(jié)合死亡率、生長(zhǎng)和消化酶變化總體分析,組合餌料(微粒子飼料+螺旋藻+搖蚊幼蟲(chóng))對(duì)黑斑原仔魚(yú)開(kāi)口效果較好。建議在標(biāo)準(zhǔn)化魚(yú)苗養(yǎng)殖中選擇微粒子飼料+螺旋藻+搖蚊幼蟲(chóng)開(kāi)口投喂。但對(duì)黑斑原整個(gè)幼魚(yú)階段和成魚(yú)期的餌料選擇和營(yíng)養(yǎng)需求還未知,有待進(jìn)一步研究。
猜你喜歡
英語(yǔ)世界(2023年10期)2023-11-17 09:19:16
汽車(chē)實(shí)用技術(shù)(2022年10期)2022-06-09 11:16:58
音樂(lè)探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級(jí)語(yǔ)數(shù)英綜合(2019年8期)2019-08-27 02:23:00
中國(guó)特種設(shè)備安全(2018年11期)2019-01-08 02:08:32
小學(xué)科學(xué)(學(xué)生版)(2018年7期)2018-08-13 09:33:04
中國(guó)非營(yíng)利評(píng)論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現(xiàn)代語(yǔ)文(2016年21期)2016-05-25 13:13:50

- 水產(chǎn)學(xué)雜志的其它文章
- 高原鰍屬魚(yú)類(lèi)線粒體DNA 和微衛(wèi)星標(biāo)記開(kāi)發(fā)研究進(jìn)展
- 流式細(xì)胞術(shù)在貝類(lèi)血細(xì)胞研究中的應(yīng)用進(jìn)展
- 鯉Cyprinus carpio、泥鰍Misgurnus anguillicaudatus與黃豆種子共生系統(tǒng)對(duì)水質(zhì)的影響
- 杭州西湖子湖湖區(qū)沉水植物群落恢復(fù)區(qū)魚(yú)類(lèi)調(diào)查與分析
- 水豐水庫(kù)入侵魚(yú)類(lèi)河鱸的年齡結(jié)構(gòu)與生長(zhǎng)特性
- 于橋水庫(kù)魚(yú)類(lèi)種類(lèi)組成及其多樣性