叢枝菌根真菌影響植物病害的研究進展
2021-07-28 07:26:00段廷玉
草業科學 2021年6期
陳 濤,段廷玉
(蘭州大學草地農業生態系統國家重點實驗室 / 蘭州大學草地農業科技學院 / 農業農村部草牧業創新重點實驗室,甘肅 蘭州 730020)
叢枝菌根(arbuscular mycorrhizas,AM)真菌是土壤微生物系統中的重要成員,是世界上分布最為廣泛的菌根真菌[1],能與90%以上的維管植物或80%的陸生植物建立互惠共生關系[2]。AM 真菌菌絲侵染植物根系產生叢枝和泡囊,通過菌絲橋連接植物根系與AM 真菌形成共生關系,這種共生關系的建立基于雙向的養分交換,因此對共生生物雙方都有利[3]。AM 真菌向寄主植物提供水和養分,從而交換寄主植物中的碳水化合物[4],如AM 真菌可以為寄主植物提供氮(氨)[5]以及其他的一些金屬和鹽類[6-7]。研究表明,AM 真菌侵染后,植物的出苗率、葉面積、莖稈質量、果實產量以及果實的營養含量顯著提高,土壤施肥頻率和施肥量減少,土壤的肥力利用率極大地提高[8-9]。
AM 真菌和植物形成的共生體除了能夠有效吸收營養物質外,還能夠通過調節植物激素水平提高寄主植物對干旱、鹽堿、重金屬等非生物逆境脅迫下的抵抗能力[10]以及增強寄主植物對于害蟲、雜草和病原菌等生物脅迫的抵抗能力[11-15]。我國農作物病蟲害種類眾多,常見農業病害775 種[16],草地植物病害2 831 種[17]。引起植物發病的生物因素主要有真菌、病毒、細菌和線蟲等。植物–微生物互作,是當前生物學和生態學研究的熱點領域,也是病蟲害綠色防控的重要技術。早在1968 年,Safir[18]首次發現了摩西球囊霉(Glomus mosseae)能夠減少紅根腐菌(Pyrenochaeta terestris)對洋蔥(Allium cepa)根部的侵染,減輕洋蔥受紅根腐菌的危害,提高洋蔥的抗病性。自此,越來越多的研究關注了AM 真菌對植物病原菌的防控作用。諸多報道指出,AM 真菌與植物所形成的共生體,能夠有效抵御病菌的危害,提高宿主植物的抗病性。AM 真菌通過改變植物的次生代謝能力,來提高植物的防御系統[19-20],比如,在植物遭受病蟲害以及逆境時,AM 真菌能夠加快多酚氧化酶(polyphenol oxidase,PPO)、過氧化物酶(peroxidase,POD)、過氧化氫酶(catalase,CAT)等的代謝過程,促進相關代謝產物的合成,提高植物的防御能力[21-23]。一般來說,與單純的植物個體相比,植物–AM 真菌共生體的防御性有較大的提高,和植株防御相關的酶活性也更高,例如PPO、POD、CAT、超氧化物歧化酶(superoxide dismutase,SOD)等[24-26]。
以AM 真菌為代表的共生微生物是潛在的重要生防劑及促生菌肥。20 世紀以來,世界農業生產力顯著提高,主要體現在化肥和農藥的使用上[27],在1990–2010 年間,農業生產水平趨于穩定[28],但由于長期使用化肥農藥,造成了病菌對藥劑產生了抗性,同時造成了土壤板結、耕地質量下降以及水體污染等問題,亟需開發新的技術,減少化肥、農藥的使用。迄今為止,AM 真菌提高植物抗病能力的研究已經取得了很大的進展,每年都有相關成果發表,涉及的植物、微生物物種更加廣泛,研究更貼近生產,更關注AM 真菌與其他微生物及環境等多因素的互作,更注重顯微技術、生物信息技術及轉錄組等新技術的綜合應用,研究日新月異。因此,有必要對該領域相關進展進行及時總結,進一步推動此方面的研究。為讀者更全面、高效地了解該領域研究進展提供便捷,同時為進一步開展此方面的研究提供理論依據。
1 研究方法
本文基于Web of Science 數據庫和CNKI 數據庫,在題目中查詢arbuscular 及其分別與diseases,rust,smut,downy mildew,podery mildew,root rot 和blight 等組合關鍵詞以及對應的中文組合關鍵詞,歸納了2018 年1 月-2020 年4 月有關AM 真菌調控植物病害的相關文獻,分析了近年來AM 真菌調控植物病害涉及的植物、病原菌及AM 真菌,總結歸納了AM 真菌對植物病害發生中宿主、環境及病原菌的影響,尤其是AM 真菌與其他生物、非生物因子協同作用對植物病害的影響;并從生理生化及分子機制歸納了AM 真菌調控植物病害發生的新進展。
2 叢枝菌根真菌調控植物病害總體概況
AM 真菌對提高宿主植物抵抗病害的作用受到病原菌、植物、AM 真菌和環境條件4 個因素的調控。植物是病原菌與AM 真菌作用的主體平臺,病原菌如何致使植物患病?早期的研究表明,病原菌接觸寄主植物后,在合適的條件下會產生入侵結構,然后侵入植物體內,在植物體內定殖擴展,通過產生各種致病效應因子如胞外酶、真菌毒素等,使植物產生病害癥狀[29],而這一過程又離不開適宜的環境條件。AM 真菌提高宿主植物的抗病性是一個復雜綜合的過程,既有可能是在局部產生抗病效應,也有可能是提高植株整體的抗病能力[30]。據此,侯劭煒等[31]提出土傳病害的“根系–根際–植株”三級防御體系,AM 真菌在根系競爭生態位點和建立機械屏障,在根際調節根系分泌物以及促進根際促生菌的定殖,在植株中促進養分吸收并誘導宿主植物的系統性防御體系,拓寬了AM 真菌協助宿主植物防御體系的范圍。
根據文獻統計,2018 年以來AM 真菌–病原菌相關文獻約計80 篇,所涉及病原菌以真菌為主。表1歸納了所查閱文獻植物病害的病原,包括真菌和細菌兩類,共10 目/科,13 屬,28 種病原微生物。其中真菌所導致的病害占據所統計病害的85%以上,這與真菌病害在所有病害中所占比重及重要性相符[32]。已經記載的植物病原真菌有8 000 種以上,真菌可以引起3 萬余種植物病害,歷史上大流行的植物病害多數為真菌引致[33]。
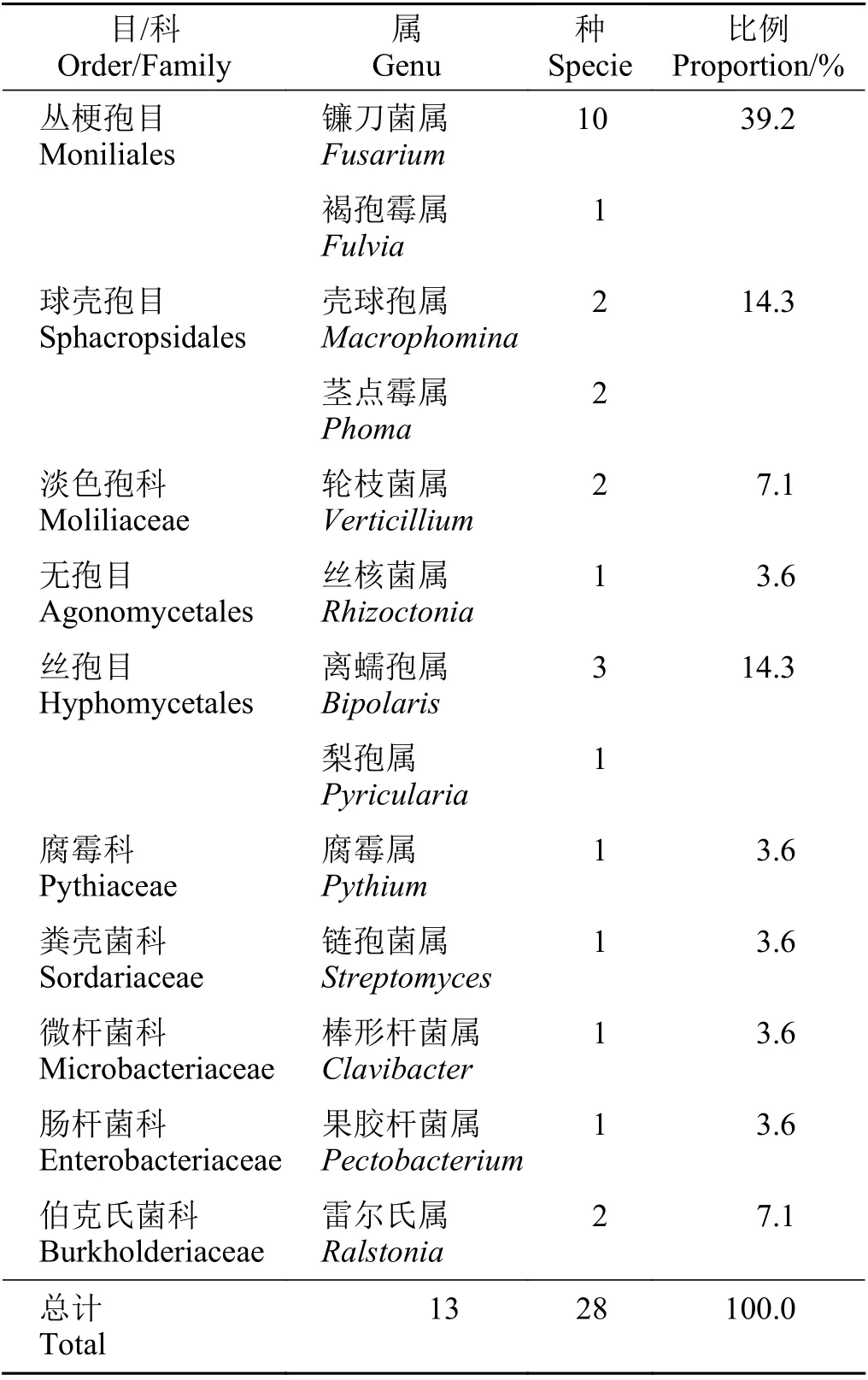
表1 AM 真菌-植物病害涉及病原類群Table 1 Pathogen groups involved in AM fungal-plant diseases
表2 統計歸納了AM 真菌與植物病害互作中的植物類群,其中茄科(Solanaceae)和豆科(Leguminosae)植物所占比例最多,共占所統計植物總數近50%。總的來說,番茄(Solanum lycopersicum)、紫花苜蓿(Medicago sativa)、多年生黑麥草(Lolium perenne)的病害研究較多,所研究的植物主要集中于栽培牧草和農作物,其他物種的研究相對較少。
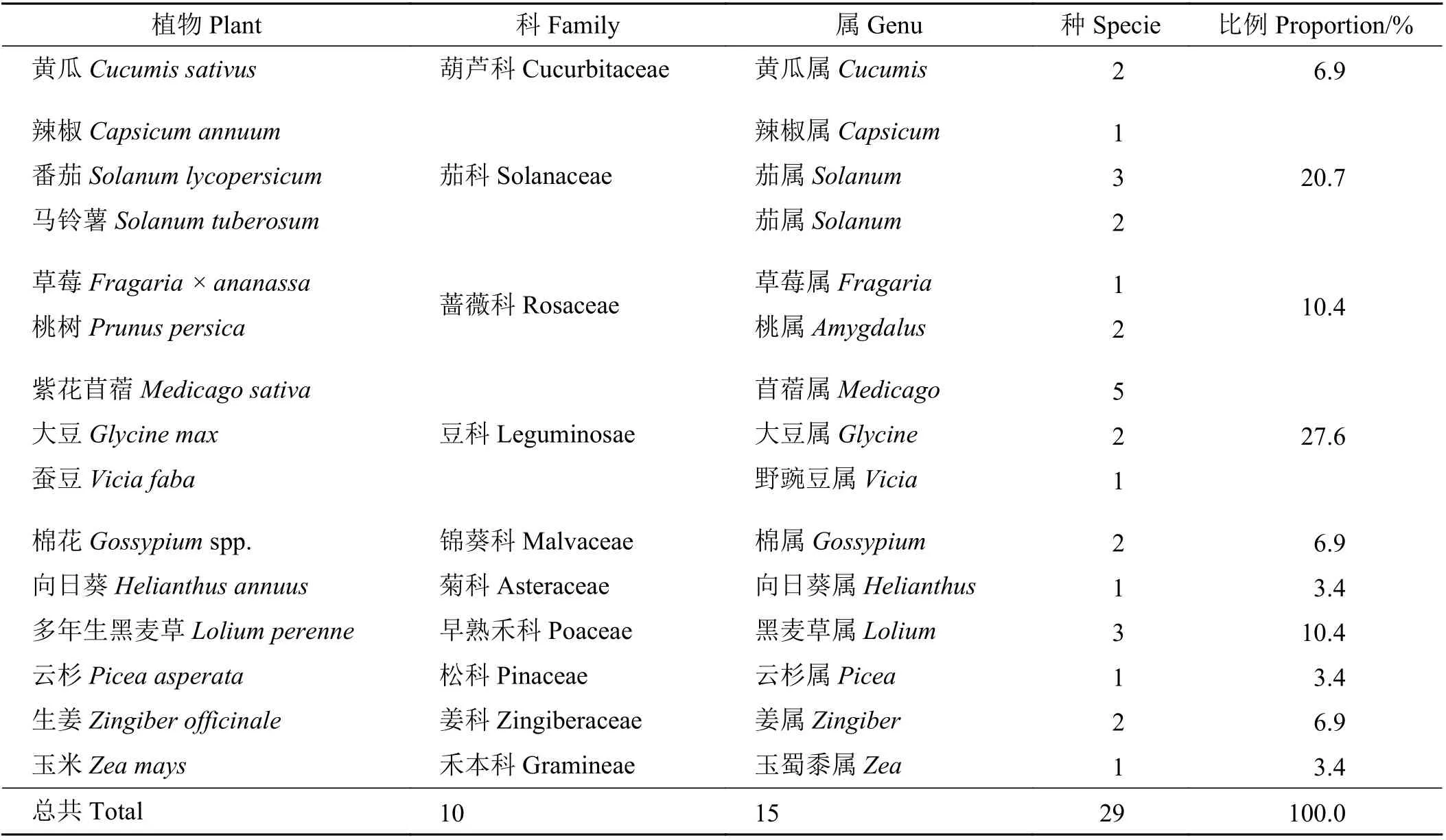
表2 AM 真菌-植物病害涉及植物類群Table 2 AM fungi-plant diseases involving plant groups
AM 真菌在自然界中分布十分廣泛,且分類變化較大,目前全世界已報道300 多種AM 真菌[34-35]。表3 統計了近年來植物病害互作的AM 真菌類群。其中球囊霉目(Glomerales)的AM 真菌占到全部統計AM 真菌的90%以上,多集中在球囊霉科(Glomeraceae)的球囊霉屬(Glomus)和斗管囊霉屬(Funneliformis)[36],諸多研究表明,球囊霉科是分布最為廣泛、研究最為深入的AM 真菌,具有廣泛的環境適應性[37]。但考慮到AM 真菌的多樣性與AM 共生體抗病能力密切相關[38]。不同的AM 真菌對同一種植物病害或同一種AM 真菌對不同植物所起的作用也不同,具有高度的多樣性[39-40]。因此,在農業生產中,選擇合適的AM 真菌種類搭配才能更有效地提高作物的抗病能力。應加強其他AM 真菌與植物–病原菌互作的研究,為充分挖掘和利用AM 真菌生防、促生功能提供依據。

表3 AM 真菌-植物病害涉及AM 真菌物種Table 3 AM fungi-plant diseases involving AM fungal species
綜合近年來該領域的研究發現,無論是葫蘆科、茄科等蔬菜,還是大豆、棉花等經濟作物,亦或是紫花苜蓿等重要牧草,研究涉及病原多為土傳病原真菌,如引致根腐病的尖孢鐮刀菌(Fusarium oxysporum)、煙色織孢霉(Microdochium tabacinum)及大麗輪枝菌(Verticillium dahliae),這一方面體現了土傳病害防控的重要性,另一方面也與土傳病害病原菌與AM 真菌共同存在于土壤中,存在直接的競爭作用有關。盡管如此,一些學者也嘗試開展了AM 真菌對非土傳病害的研究,如Campo 等[41]報道了AM 真菌可提高水稻(Oryza sativa)對稻瘟病抗性,鑒于非土傳病害的重要性和危害性,今后應加強此方面的研究。
3 AM 真菌對植物病害發生的影響
AM 真菌對植物病害產生的影響具有高度的多樣性,總的來說,可分為三類:一是通過提高植物自身系統抗性影響病害的產生,二是影響植物生長環境降低病原菌的生存范圍,三是影響與AM 真菌協同的因素使植物自身或者生存環境產生有益變化來提高抗病能力。這些影響效果大部分對植物抗病具有正向作用,少部分具有負向作用。
3.1 AM 真菌對植物本身的影響
AM 真菌能夠提高植物的養分吸收能力、促進生長、提高根系活力,進而抵抗病菌侵染。這一直是AM 真菌–植物–病原菌互作的基本內容,近2 年多的文獻亦是如此,如尖孢鐮刀菌引起的紫花苜蓿根腐病中,接種摩西管柄囊霉(Funneliformis mosseae)的紫花苜蓿莖葉干重相比不接種的對照提高23.4 倍,根系干重提高15.0 倍,根系總氮和總磷含量增加54.3 和51.3 倍[42]。Jamiolkowska 等[43]研究尖孢鐮刀菌引起辣椒(Capsicum annuum)幼苗根腐病過程中發現,接種AM 真菌對已感染病菌的辣椒幼苗具有一定的保護作用,幼苗的主根出現少量的病變壞死,側根強烈拉長。AM 真菌能夠提高植物的光合效率[44]、維持光合活性[45]。AM 真菌提高植物體內激素水平以及抗氧化酶的活性,降低有害物質的含量。尖孢鐮刀菌能夠引起黃瓜(Cucumissativus)的枯萎病,在黃瓜幼苗期接種地表球囊霉(G. versiforme)能夠保護幼苗,減少病原菌的入侵[46],試驗表明,雙接種地表球囊霉和摩西管柄囊霉相比單接種尖孢鐮刀菌,使黃瓜幼苗的吲哚乙酸(IAA)含量提高667%,赤霉素(GA3)含量提高223%,玉米素核苷(ZR)含量提高254%,茉莉酸(JA)濃度增加215%;脫落酸(ABA)含量降低53%[47]。Li 等[48]對苜蓿莖點霉(Phoma medicaginis)引起的紫花苜蓿春季黑莖病研究發現,接種根內球囊霉(G. intraradices)能夠使感病紫花苜蓿的過氧化物酶活性提高58.21%,多酚氧化酶活性提高49.64%,水楊酸(SA)濃度增加62.95%,茉莉酸濃度增加38.18%。AM 真菌對感病植株的影響因AM 真菌種類不同而異。AM 真菌對植物的生長發育具有顯著的促進效應,然而也有研究表明,AM 真菌會抑制植物的生長。Ravnskov等[39]對終極腐霉(Pythium ultimum)引起黃瓜猝倒病的研究發現,AM 共生體的生物防治策略均具有獨特的表型寄主植物對AM 真菌的反應,即AM 真菌–植物的生物防治效果取決于共生AM 真菌的基因型,接種摩西管柄囊霉完全抵消病原菌引起的生長抑制,促進植物生長;接種根內球囊霉一方面會抑制植物生長,另一方面又抵消了病菌引起的生長抑制,接種近明球囊霉(C. claroideum)不僅會抑制植物生長,亦未提高植物抗病性,不具有生物防治作用,這也體現了植物–微生物互作中的選擇策略及多樣性。在作物進化方面,AM 真菌對野生種更具有共生優勢。比如,與已馴化的栽培水稻相比,野生水稻還保留著馴化過程中失去的一些共生優勢,馴養過程會影響與水稻共生的AM真菌種群,甚至降低AM 真菌的環境適應性。因此,野生水稻具有更優的共生水平和更高效的抗病基因表達途徑[49]。
3.2 AM 真菌對植物生長環境的影響
AM 真菌可以通過影響植物根系周圍的土壤環境和微生物活性來影響植物病害。植物根系對磷的吸收速率遠高于土壤磷的擴散速率,從而在根系水平上形成一個貧化帶,限制了土壤磷向植物的供應[50],而可靠的磷源和維持細胞內磷穩態是生命所必需的,與AM 真菌共生是植物進化出的一個應對策略[51]。AM 真菌與植物形成共生體后,產生大量根外菌絲,形成菌絲網,根外菌絲及菌索的生長使植物根系擴展到更遠的土壤,吸取更大范圍的養分[52-53]。此外AM 真菌也能活化土壤磷,AM 真菌的菌絲網絡可以訪問和動員植物的不可利用磷,從而提高植物的磷獲取效率,提高植物利用土壤磷生產生物量或產量的能力[54-55]。AM 共生體對植物養分吸收的不同可能會對植物抗病性產生不同的影響,Ravnskov 和Larsen[56]分別研究了6 個與3 種AMF 共生的水稻品種在AM 共生中的功能兼容性發現,水稻植株的營養成分取決于水稻品種和AMF 品種的組合,在共生中,這種AM 共生體在營養成分上的功能相容性差異也可能影響植物對病原菌侵染的耐受性。Li 等[57]對尖孢鐮刀菌引起草莓(Fragaria ananassa)枯萎病的研究發現,接種摩西管柄囊霉能夠提升根系土壤的細菌多樣性進而促進根際細菌同病原菌競爭生態位,減少病原菌的侵染位點。
3.3 AM 真菌與其他因子協同作用
統計近年來的文獻研究發現,AM 真菌與其他因素協同作用影響植物病害,是當前AM 菌根研究的新熱點。AM 真菌可以與根際促生菌(plant-growthpromoting rhizobacteria,PGPR)[47,58]、各種環境脅迫[44]、土壤養分條件[59-61]、外生菌根真菌(ectomycorrhizal fungi,ECMF)[62]、植物促生酵母[63]、禾本科內生真菌[60,64]、根瘤菌[65-67]、田間耕作方式[68-69]等因素單一協同或者復合協同,不僅能夠加強植物對養分的吸收能力,促進植物生長,還能從整體上提高植物對病原菌的抵抗能力。AM 真菌與生物因素協同涉及的根際微生物包括:根際促生菌(PGPR)、外生菌根真菌、植物促生酵母、禾草內生真菌(Epichlo?)和根瘤菌。AM 真菌與根際促生菌共同作用,能夠使黃瓜的根系活力提高121%以上[47]。劉貴猛等[58]對青枯雷爾氏菌(Ralstonia solanacearum)引起生姜(Zingiber officinale)青枯病的研究表明,地表球囊霉能夠有效促進根際促生菌S3-11 定殖生姜根圍,摩西管柄囊霉在一定的條件下促進S3-11 和S1-10 在根圍的定殖,生姜發棵期S1-10 能夠顯著提高根內球囊霉的侵染率,但會顯著降低地表球囊霉的侵染率,塊莖膨大期S3-11促進根內球囊霉和地表球囊霉的侵染,根內球囊霉的侵染率最高可達64%;因此,AM 真菌與協同因素的互作不僅僅是單一的相互增益,部分協同因素會降低AM 真菌對植物的侵染率,而高定殖率的AM 真菌–植物共生體對寄主植物的抗病能力提升更多。Chen 等[59]研究表明,在低氮條件下,接種AM 真菌能夠顯著提高三裂葉蟛蜞菊(Wedelia trilobata)對立枯絲核菌(Rhizoctonia solani)的抗性。AM 真菌與外生菌根真菌均能夠侵染紫蘇(Perilla frutescens)形成菌根,侵染率分別達到82.36%和73.25%,單接摩西球囊霉(G. mosseae)能夠使紫蘇的SOD、POD、CAT 和苯丙氨酸解氨酶(phenylalanine ammonia-lyase,PAL)活性分別提高49.95%、 68%、 222.67%和 81.95%, 降低丙二醛(malondialdehyde,MDA)濃度28.53%;單接外生菌根真菌使紫蘇的SOD、POD、CAT、PAL 活性分別提高39.81%、58.73%、111.91%和74.44%,MDA 濃度降低16.42%,二者混合接種的效果高于單接效果[62]。AM 真菌和促生酵母協同能夠提高向日葵(Heliantus annuus)的養分吸收能力,降低MDA 的含量[63]。AM 真菌對某些協同因素起到促進作用,也會對某些協同因素產生不利影響;比如,AM 真菌和禾草內生真菌共同侵染多年生黑麥草時,禾草內生真菌會降低AM 真菌的定殖率[60,64]。AM 真菌與根瘤菌是相互促進、相互依賴的,根瘤菌能提高AM 真菌的定殖率,AM 真菌能夠促進根瘤的形成[66],二者交互作用下的效果高于單一作用,能夠顯著提到抗氧化酶的活性,降低有害物質濃度[42,67]。
單一作物連作會使土地養分失衡,有害微生物大量繁殖。間作條件下AM 真菌在蠶豆上的定殖率會高于單作20.9%,間作條件下接種AM 真菌能夠降低尖孢鐮刀菌數量96.2%,使蠶豆發病率和病情指數降至0[68]。在農業生產中,作物的產量和土地的生產力水平不是單一因素影響的,針對不同的作物和環境條件選擇合適的AM 真菌種類以及配比,能夠幫助植物抵抗病原菌的侵染,穩定植物所處生態系統,整體上提高植物的抗病能力。
4 AM 真菌影響植物病害機理
AM 真菌與病原菌互作的機理是復雜的,是多種機理協同作用的結果。總的可以將AM 真菌影響植物病害的機理分為生物學及生理生化機理、分子機理和多因素協同作用機理3 類,其中多因素協同機理是近2 年多AM 真菌抗病機理研究領域的熱點。
4.1 生物學及生理生化機理
AM 真菌影響植物病害的生物學及生理生化機理主要有兩個方面,一方面是通過改善土壤環境,降低病原菌的數量,促進植物吸收養分,豐富根際微生物多樣性等方式提高植物自身活力間接發揮抗病作用;另一方面則是通過提高植物體內激素和抗氧化酶等的活性,降低體內有害物質等方式直接提高植物的抗病性。研究表明,與AM 真菌形成共生體的植物對病原菌的入侵能夠產生快速的防御反應[70-72]。在菌根化的過程中,AM 真菌會誘導寄主植物防御反應,導致局部或系統的激活寄主免疫,這稱為植物的啟動狀態(primed state),這種狀態下植物受到潛在敵人攻擊時會更快更有效地觸發防御反應[73]。“根際–根系–植株”三級防御體系[31]可為AM 真菌增強植物的抗病能力做形象的闡釋。在植物根系,AM 真菌能夠提高土壤有機質的含量,調節土壤環境[57]。Jonas等[74]認為AM 真菌很可能通過釋放來自寄主植物根中富含能量的碳化合物的分泌物來增加土壤有機質的含量。AM 真菌促進根系發育,即使植物感染病菌,也能通過促進側根的發育來擴大植物根系吸收養分的范圍。在植物的根際,AM 真菌能夠提高土壤根際磷酸酶活性[65],豐富根際微生物區系[75],有效降低根際土壤中可培養真菌的數量,增加可培養細菌和放線菌的數量[48]。為細菌提供特定的生態位,增加細菌多樣性[76],細菌能夠與病原菌競爭生態位,降低土壤中殘留的病原菌對植物根系的感染程度[57],AM 真菌能夠與病原菌競爭定殖位點和光合產物[64]。AM 真菌與病原菌處于相互拮抗的地位,AM 真菌能夠有效抑制病原菌的侵染,病害極其嚴重的植物體內,病原菌會抑制AM 真菌。唐燕等[77]通過田野調查發現無病正常的星油藤(Plukenetia volubilis)中AM 菌絲侵染率可達90%,發病嚴重的植株中基本不存在叢枝和泡囊。在植物自身方面,AM 真菌能夠纏繞病菌菌絲,阻擋其侵入根系[47]。AM 真菌對植物的抗病性會因為共生體對養分吸收能力的不同而有變化,定殖率高的共生體能夠大幅度提高植物吸收養分的能力,能夠促使植物生長,使植株健壯,從而反饋給AM真菌更多的營養物質,增加AM 真菌的定殖位點[56]。不同的AM 菌劑誘導的植物體內防御應激反應會有所差別[78],但都能通過增強植物的內部抗性和系統抗性來誘導植物產生各種次生代謝產物,調節植物激素水平以及提高防御性酶的活性,最終不同程度地提高植物整體的抗病能力。
4.2 分子機理
近年來,隨著基因組學的發展,大量的植物抗病基因(resistance gene,R)和病原菌無毒基因(avirulence gene,Avr)被克隆,植物的免疫反應機制研究有了重大進展。植物在長期的進化過程中形成了兩類反應機制:由病原菌模式分子(pathogenassociated molecular patterns,PAMPs)引發的免疫反應(PAMP-triggered immunity,PTI)和由效應因子引發的免疫反應(effector-triggered immunity,ETI)[79]。AM 真菌可以通過調節防御途徑轉導相關基因的表達來減輕植物病害的嚴重程度[48]。Marquez 等[80]對菜豆殼球孢菌(Macrophomina phaseolina)引起大豆(Glycine max)炭腐病的研究發現,AM 真菌上調病程相關蛋白、抗病蛋白、轉錄因子和次生代謝的相關基因,上調水楊酸途徑相關的PR1防御基因,與絲氨酸羧肽酶和凝集素前體相關的基因被高度誘導。AM 真菌侵染大豆根系表現出編碼PRPs基因家族、絲氨酸蛋白酶、受體激酶和苯丙烷途徑衍生物的基因轉錄增加[81]。AM 真菌定殖桃樹(Prunus persica)引發PpCHI、PpLOX1、PpLOX5、PpAOC3、PpAOC4、PpOPR2基因在根部表達,提高茉莉酸濃度,進而提高抗病能力[82]。Prihatna 等[83]研究發現菌根共生的幾個常見基因中,CYCLOPS/IPD3基因是共生所必需的,雖不涉及對病害的抗性,但是更高程度的共生能夠更大程度上提高植物自身的抗性。
4.3 與其他因素協同機理
農業生產中,影響作物產量和生產力的因素很多,是多因素共同作用下的結果。AM 真菌與其他因素的協同機理主要體現在兩個方面,一方面是AM 真菌與協同因素相互依賴,共同提高植物抗病能力,例如AM 真菌與根瘤菌的協同,能夠全面提高紫花苜蓿對根腐病、黑莖病、枯萎病的抗性[65-67]。另一方面是與協同因素的相互拮抗,抵消不利協同因素(干旱、營養不良環境等)對病害的促進效應。死谷芽孢桿菌HJ-5 與AM 真菌協同對大麗輪枝菌(Verticillium dahliae)引起的棉花(Gossypiumspp.)黃萎病有顯著地抵抗效應,AM 真菌提高根際HJ-5 定殖數量,HJ-5 分泌脂肽類物質伊枯草菌素(iturin A)和表面活性素(surfactin),導致病原菌菌絲畸形[84]。相對的,根際微生物也可以促進AM 共生體的發育[85]。蚯蚓和AM真菌共同作用下,土壤有機質含量和土壤結構改變,有利于植物吸收養分[56]。對AM 真菌與其他因素的協同研究更容易將研究成果轉化為生產力。同一作物連作對土壤的傷害嚴重,并且土壤連作病害會嚴重抑制AM 真菌的定殖[86],而AM真菌和間作能夠有效提升土壤的生產能力,保護土壤恢復[68]。AM 真菌的多因素協同機理很復雜,參與進協同系統的因素越多,產生的效果越難以控制。綜上所述,不論是相互依賴型的協同系統還是相互拮抗型的協同系統,AM 真菌總是能與協同因素一起促進植物的抗病能力,對于拮抗型協同系統,AM 真菌產生的有益促進效果總是能夠多于不利因素的消極效果,在植物整體水平上加強植物的抗病能力。
5 展望
植物病害是影響生產力的重要因素,AM 真菌在提高植物防御和抵抗病原物入侵的能力、防控土傳病害與緩解連作障礙方面表現出巨大的應用前景與生產價值。盡管目前關于AM 真菌與病原物之間的互作研究已經取得了長足的進展,但是還有一些問題亟待解決。首先,目前研究涉及的AM 真菌與植物、病原與已知的AM 真菌–植物–病原菌多樣性相比遠遠不夠,需要更廣泛、深層次的研究。同時,考慮到AM 真菌在土壤中具有高度多樣性,目前研究多是單個AM 真菌的研究結果,因此未來需關注AM 群落的抗病功能的研究。其次,在AM 真菌對植物病害的影響方面,目前AM 真菌與植物的互作多停留在溫室盆栽研究以及AM 真菌對植物病害的影響效果和機制,而缺乏生物學、生態學以及AM 真菌能否對植物遺傳進化等方面的研究,應加強AM 真菌與植物互作的分子層面研究和AM 真菌在生態系統中對植物影響層面研究。多因素的協同效應更貼近自然環境,雖然已有AM 真菌與單一因素協同對植物抗病能力影響的研究,但是更多的研究還是偏向于人工模擬下的AM 真菌–植物的互作,因此需要加強人工模擬下AM 真菌與多因素協同的研究和大田條件下可控的多因素協同研究。在機理方面,AM 真菌能夠通過促進植物吸收養分和光合作用等提高植物自身的健康活力,間接提高植物的抗病能力,也能通過調節生理生化代謝等方面直接提高抗病能力。近年的研究偏向于多因素影響下AM 真菌與植物的互作效應,然而更多的研究只停留在表象,對于機理未有深入探討,這是未來研究的一個新方向。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
