miR-322-5p通過NOX4調控細胞凋亡抑制大鼠神經管畸形發生
2021-08-02 04:29:10黃琬淇劉玉思顧卉袁正偉
中國醫科大學學報 2021年7期
黃琬淇,劉玉思,顧卉,袁正偉
(中國醫科大學附屬盛京醫院,國家衛生健康委員會小兒先天畸形重點實驗室,沈陽 110004)
神經管畸形(neural tube defects,NTDs)作為最嚴重的出生缺陷之一,全球發病率約為1‰~10‰[1]。由于神經管的閉合失敗,可出現如無腦兒、脊柱裂、腦脊髓膜膨出等一系列表型[2]。神經上皮細胞的過度凋亡導致神經管閉合異常,產生NTDs[3]。全反式維甲酸(all-trans retinoic acid,ATRA)可通過誘導細胞凋亡活動影響神經管閉合,引起NTDs的發生[4-5],本研究組前期利用ATRA成功制作了NTDs胎鼠模型。
微RNA(microRNAs,miRNA)是一種長度為22~24 nt的內源性小型非編碼 RNA 分子,通過與靶基因的3’端非翻譯區(3’untranslated regions,3’-UTR)結合,誘導mRNA降解或抑制翻譯,從而調控靶基因的轉錄和表達[6]。miR-322是細胞增殖、分化、凋亡等生物學活動的重要調控因子,對于心血管系統、神經系統及其他器官均有一定的保護作用[7-8]。研究發現,miR-322-5p可抑制高糖誘導的小鼠神經干細胞凋亡和NTDs的發生。本課題組前期研究[9-10]發現,胚胎體外注射miR-322-5p-mimic,對NTDs有治療作用。但它與ATRA誘導的大鼠NTDs的關系仍有待研究。
煙酰胺腺嘌呤二核苷酸磷酸氧化酶4(nicotinamide adenine dinucleotide phosphate oxidase 4,NOX4)是NADPH氧化酶家族的一個亞型成員,它的過表達可導致神經元的自身毒性以及大腦的局部缺血[11]。在高糖誘導的神經病變動物模型中,周圍神經系統施旺細胞NOX4表達上調,促進細胞氧化應激參與周圍神經損傷[12]。但NOX4在ATRA誘導的大鼠NTDs中的表達及作用尚未見報道。通過TargetScan及starBase生物信息學分析發現,NOX4是miR-322-5p的靶點,在NOX4mRNA的非翻譯區存在miR-32-5p的潛在結合位點。本研究擬觀察miR-322-5p與NOX4在ATRA誘導的大鼠NTDs模型中的表達及其對凋亡活動的影響,探討NTDs的發生機制,以為其診療提供新靶點。
1 材料與方法
1.1 材料
小鼠神經干細胞(C17.2,貝納創聯生物研究所),ATRA(美國Sigma公司),MEM培養基(美國Gibco公司),MEM NEAA(美國Gibco公司),胎牛血清(美國Gibco公司),青霉素鏈霉素溶液/雙抗(美國Hyclone公 司),miR-322-5p mimics/inhibitor、mimics/inhibitor-NC(上海吉瑪制藥技術有限公司),NOX4 plasmid(上海吉凱基因化學技術有限公司)。JetPRIME(法國PolyPlus-transfection公司),miRNeasy Mini Kit(美國Qiagen公司),miRNA第一鏈cDNA合成試劑盒(上海生工生物工程有限公司),SYBR Premix Ex Taq Kit(大連寶生物工程有限公司),BCA蛋白定量試劑盒(北京索萊寶科技有限公司),NOX4兔多克隆抗體(美國Rocklamd公司),Bcl-2鼠多克隆抗體(美國Sigma公司),Bax兔多克隆抗體(美國Cell Signaling Technology公司),β-actin鼠單克隆抗體、辣根過氧化物酶標記的山羊抗兔/抗小鼠IgG(美國Proteintech公司),UltraSensitive SP IHC kit、辣根過氧化氫DAB顯色試劑盒(福州邁新生物技術開發有限公司),4’,6-二脒基-2-苯基吲哚(4’,6-diamidino-2-phenylindole,DAPI)(上海碧云天生物技術有限公司)。
1.2 方法
1.2.1 大鼠NTDs模型制備及組織樣本收集:Wistar大鼠(北京華阜康生物技術有限公司)飼養于中國醫科大學附屬盛京醫院SPF級實驗動物房,按雌∶雄為2 ∶1于前一晚合籠,第二日早將大鼠分籠,陰道涂片見精子的雌鼠單獨放于一籠,記為孕0 d(E0)。于E10將已孕雌性大鼠隨機分為2組,NTDs組灌胃140 mg/kg橄欖油溶解的ATRA,對照組給予同體積的橄欖油灌胃。于E11剖宮取出胎鼠,獲取脊髓組織。
1.2.2 免疫組織化學染色:將胚鼠脊髓固定于4%多聚甲醛中,石蠟包埋,制4 μm厚切片,二甲苯脫蠟,梯度乙醇脫水,于去離子水中水化。抗原修復后,使用UltraSensitive SP IHC試劑盒處理切片,滴加一抗anti-NOX4(1 ∶800),4 ℃孵育過夜,第二日DAB顯色,蘇木素復染,流水返藍1 h除去余色,封片拍照。
1.2.3 免疫熒光染色:切片在二甲苯中脫蠟,梯度乙醇脫水,去離子水水化,0.1% Triton X-100處理。動物血清封閉后,加入anti-NOX4抗體(1 ∶1 000),4℃孵育過夜。PBS沖洗3次,室溫熒光標記的二抗孵育(1 ∶200)2 h。DAPI染色,熒光顯微鏡觀察。
1.2.4 細胞的培養和轉染:用含10%胎牛血清、1%MEM NEAA、100 μg/mL青霉素、100 μg/mL鏈霉素的MEM培養液在37 ℃、5%CO2培養箱中培養小鼠神經干細胞C17.2。接種細胞至6孔板,轉染前培養24 h。以JetPRIME為轉染試劑,轉染miRNA-空白對照(miRNC)、miR-322-5p mimic、miR-322-5p inhibitor以 及NOX4高表達質粒至C17.2細胞。
1.2.5 實時定量PCR:用miRNeasy Mini試劑盒從胚胎脊髓組織和C17.2中提取總RNA,用miRNA First Strand cDNA Synthesis反轉錄RNA。以U6為內參,用SYBR Premix Ex Taq試劑盒行實時定量PCR。引物序列如下:miR-322-5p F鏈為AGCAGCAATTCATGTTT TGGAA,R鏈為GCTGTCAACGATACGCTACGTAAC;U6F鏈為CTCGCTTCGGCAGCACA,R鏈為AACGCT TCACGAATTTGCGT;β-actinF鏈為GGAGATTACTGC CCTGGCTCCTA,R鏈為GACTCATCGTACTCCTGCTT GCTG。
1.2.6 Western blotting:自胚胎脊髓和C17.2細胞中提取蛋白質。SDS-PAGE電泳分離,電轉印至PVDF膜,5%脫脂牛奶封閉2 h。加入一抗 NOX4(1 ∶1 000),Bax(1 ∶1 000),Bcl-2(1 ∶1 000),4 ℃孵育過夜。次日用TBST洗膜3次(10 min/次),室溫下與HRP兔和鼠二抗(1 ∶5 000)孵育2 h,ECL發光。
1.3 統計學分析
采用SPSS 26.0軟件進行統計分析,并用 GraphPad 8.0統計軟件作圖。計量資料以表示,采用獨立樣本t檢驗進行比較。P< 0.05為差異有統計學意義。
2 結果
2.1 miR-322-5p和NOX4在大鼠NTDs形成中的表達
2.1.1 ATRA誘導的NTDs胎鼠脊髓組織神經管形態及NOX4定位表達情況:免疫組織化學染色和組織免疫熒光結果顯示,NTDs組神經管形態明顯異常,且胚胎脊髓中NOX4表達顯著高于對照組,見圖1。

圖1 E11對照組和NTDs組大鼠胚胎脊髓組織神經管的形態和NOX4表達情況 ×200Fig.1 Morphology and NOX4 expression of spinal cord neural tube between E11 control and NTDs rats ×200
2.1.2 miR-322-5p和NOX4在NTDs胎鼠脊髓組織中的表達情況:實時定量PCR和Western blotting結果顯示,與對照組相比,NTDs組脊髓組織中miR-322-5p mRNA和蛋白表達水平明顯下調(均P< 0.05),見圖2。提示miR-322-5p的表達對NTDs可能有抑制作用,而NOX4可能促進NTDs的形成。
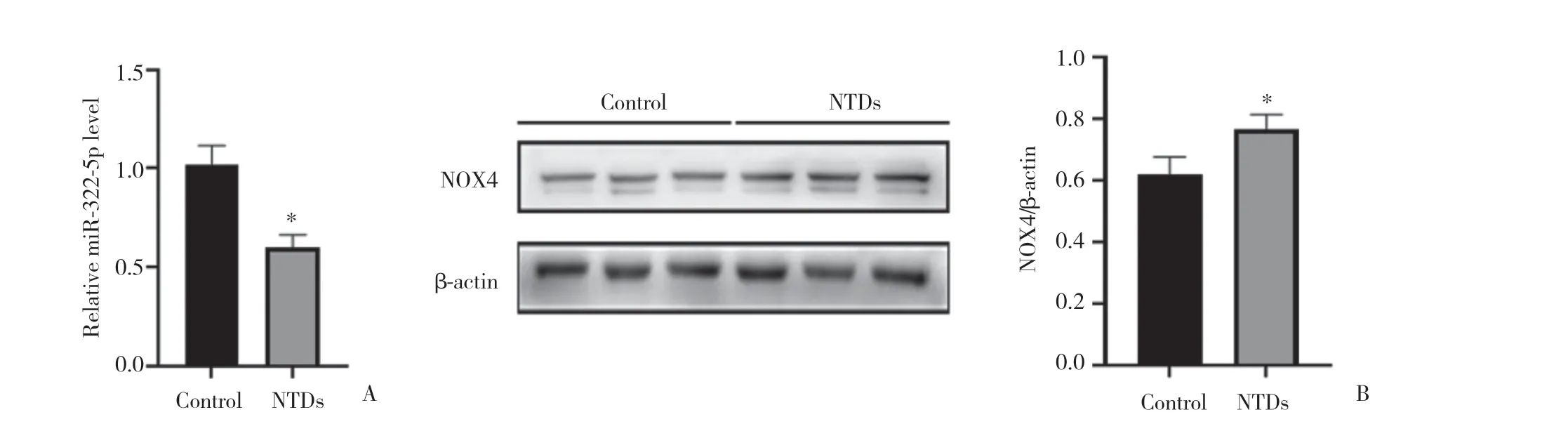
圖2 E11對照組和NTDs組大鼠胚胎脊髓組織miR-322-5p和NOX4表達情況Fig.2 Comprison of miR-322-5p mRNA and NOX4 protein expression in the embryonic rat spinal cord tissues between control and NTDs groups
2.2 ATRA致畸胎鼠脊髓中凋亡蛋白的表達情況
Western blotting結果顯示,ATRA誘導的NTDs組中,NOX4蛋白表達顯著上調,促凋亡蛋白Bax表達增強,抗凋亡蛋白Bcl-2表達減弱(P< 0.05),見圖3。表明NTDs胎鼠脊髓中細胞凋亡增加,提示NOX4可能促進凋亡的發生。
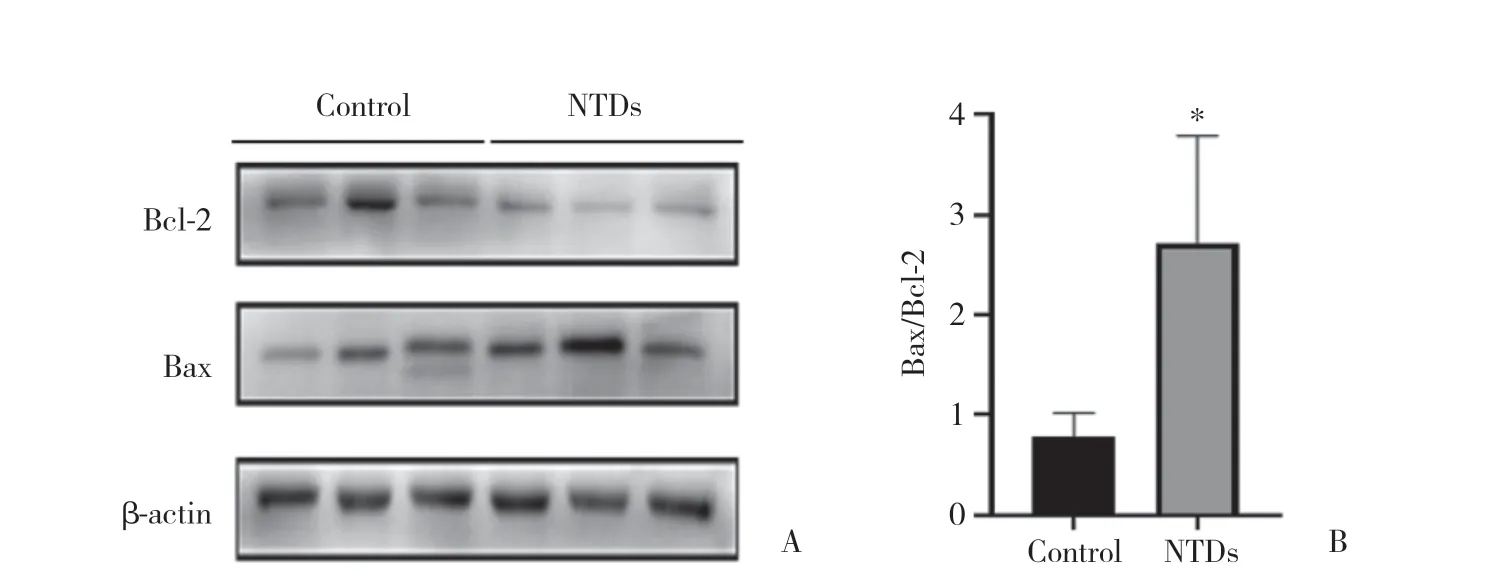
圖3 凋亡相關蛋白在對照組和NTDs組胎鼠脊髓組織中的表達情況Fig.3 The expression of apoptosis-related proteins in the embryonic rat spinal cord of control and NTDs groups
2.3 miR-322-5p負調控NOX4的表達并減少細胞凋亡
2.3.1 轉 染miR-322-5p mimics和inhibitor后C17.2神經干細胞中NOX4的表達:Western blotting結果顯示,C17.2細胞中轉染miR-322-5p mimic 48 h后,miR-322-5p表達水平顯著上調(圖4 A,P< 0.05),NOX4蛋白表達下調(圖4B、4C,P< 0.05);轉染miR-322-5p inhibitor后,miR-322-5p表達水平下調(圖4D,P<0.05),NOX4表達下調(圖4E、4F,P< 0.05)
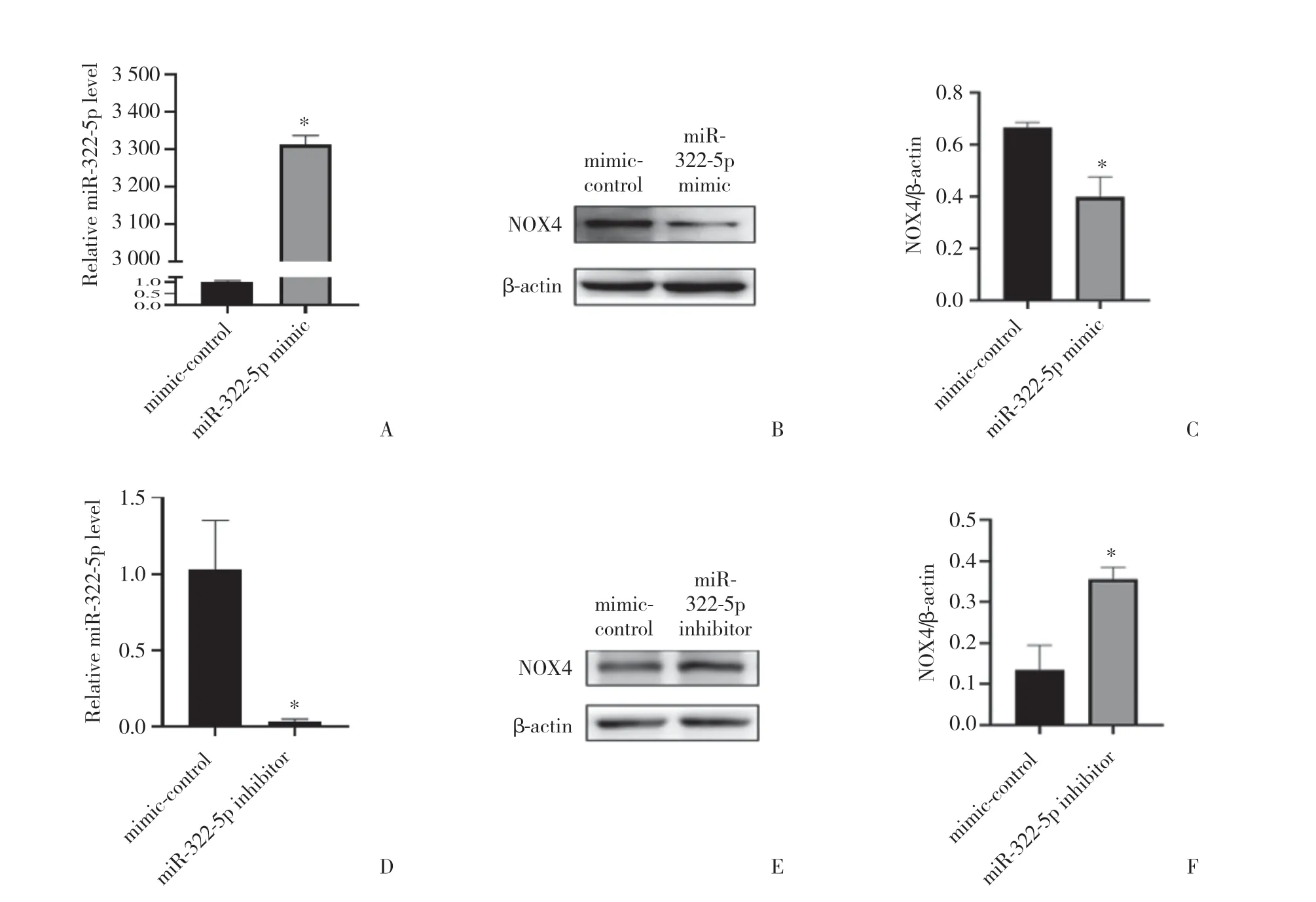
圖4 轉染miR-322 mimic/inhibitor對C17.2細胞中NOX4表達的影響Fig.4 Regulation of NOX4 after transfection of miR-322 mimic/inhibitor
2.3.2 miR-322/NOX4對神經干細胞的凋亡的影響:Western blotting結果顯示,轉染NOX4高表達質粒的神經干細胞中,NOX4過表達,促凋亡基因Bax蛋白表達上調,抗凋亡基因Bcl-2蛋白表達下調。miR-322-5p mimic與NOX4過表達載體共轉染細胞后,miR-322-5p mimic抑制NOX4表達的同時,也抑制了Bax表達,促進了Bcl-2表達(P< 0.05),見圖5。提示miR-322-5p能夠通過抑制NOX4表達抑制神經干細胞的凋亡。

圖5 C17.2細胞轉染miR-322-5p mimic和NOX4后NOX4和凋亡蛋白的表達水平Fig.5 Expression of NOX4 and apoptotic proteins after transfection miR-322-5p mimic and NOX4 of C17.2
3 討論
NTDs作為一種多因素疾病,對于其發病機制的了解仍較局限。本研究利用ATRA誘導大鼠NTDs模型,發現NOX4作為miR-322-5p的靶標,可以通過參與細胞凋亡調控大鼠神經管閉合的過程,在大鼠NTDs發生中扮演重要角色。
細胞凋亡對于維持胚胎的正常發育具有重要作用,在神經管閉合進程中,中線細胞的過度凋亡導致神經管的閉合失敗以及一系列NTDs表型的形成[13]。研究[14]發現,miRNA對NTDs具有調控作用。在高糖環境下,miR-129-2通過抑制細胞自噬可誘導NTDs發生[15]。本課題組前期研究[16]發現,孕產婦糖尿病或體外高血糖會增加miR-200b的水平,CITED2是miR-200b的靶標,被高糖下調。由此可見,miR-200b抑制劑可逆轉培養的胚胎中高葡萄糖誘導的CITED2下調,內質網應激,進而改善胚胎發育中的NTDs。miR-322也參與了細胞凋亡活動的調控[17]。缺氧條件可引起心肌細胞出現凋亡,并伴隨miR-322明顯上調[18]。本研究組前期還發現miR-322-5p可以通過抑制TRAF3減少高糖誘導的神經干細胞凋亡[9]。但miR-322-5p是否參與ATRA誘導的大鼠NTDs尚無報道。
NOX4作為NOX酶家族成員是細胞超氧陰離子的來源,參與中樞神經系統的發育、神經干細胞生物學以及成熟神經元的功能[19],并可通過產生活性氧(reactive oxygen species,ROS)來誘導細胞發生氧化應激,進而導致細胞凋亡。研究[20-21]顯示,糖皮質激素可通過NOX4/ROS/p38 MAPK途徑誘導軟骨細胞凋亡,心肌細胞NOX4高表達,凋亡也隨之增加。由于神經元的抗氧化性較弱,ROS的產生是神經系統疾病和細胞凋亡的主要因素。NOX4對神經元的凋亡也具有相似作用,它參與大鼠缺血再灌注過程中ROS誘導的神經元凋亡和腦損傷[22]。此外,慢性氟暴露會誘導中樞神經系統發生氧化應激,誘導NOX4的表達,而NOX4高表達會導致血腦屏障損害,小膠質細胞活化和神經元喪失,從而導致腦功能受損[23]。但關于大鼠NTDs胚胎中脊髓NOX4的表達尚無研究。本研究結果顯示,在NTDs胎鼠模型脊髓中,NOX4表達水平上調,且伴隨促凋亡相關蛋白表達增加,抗凋亡相關蛋白減少。結合生物信息學分析的結果,NOX4是miR-322-5p的靶基因,當神經干細胞C17.2在轉染NOX4高表達質粒的同時轉染miR-322模擬物,可降低NOX4的表達水平,且伴隨抗凋亡相關蛋白增加,促凋亡蛋白減少。由此可見,miR-322-5p能夠抑制NOX4的表達,抑制神經管發育過程神經細胞的過度凋亡。
綜上所述,miR-322-5p能夠下調NOX4的表達,減少大鼠胚胎神經管閉合過程中細胞凋亡,減少NTDs的發生,因此,miR-322-5p可能成為NTDs治療的新靶點。
