高寒草甸主要組分種開花物候對氮素添加的響應
2021-08-03 09:21:04劉旭東章志龍杜國禎
草業科學 2021年7期
劉旭東,章志龍,杜國禎
(1.蘭州大學生命科學學院 / 蘭州大學草地農業生態系統國家重點實驗室,甘肅蘭州 730000;2.甘肅民族師范學院化學與生命科學系,甘肅合作 747000)
物候是生物受環境因素驅動的周期性現象[1],植物物候對陸地生態系統變化具有重要的指示作用,是響應區域環境變化最敏感的性狀之一[2-3]。物候是研究植物響應氣候環境變化最簡單的方式[4],植物物候與環境變化(溫度、水分和土壤養分)存在高度的關聯性[5],是植物應對選擇壓力的適應性策略[6]。在全球變化日趨明顯的背景下,物候變化不僅關系到植物個體的生存和繁衍,還將對群落生物多樣性的維持產生重要影響[7]。
全球氮沉降的加劇被視為影響陸地生態系統結構和功能的重要驅動因子[8-9]。氮是高寒生態系統最重要的限制性養分元素[10],外源性氮輸入則能夠減輕氮營養對植物生長的限制[11]。高寒生態系統對環境變化的響應具有高度的敏感性[12-13],并對這些變化的響應具有超前性[14]。氮營養作為重要的環境因子明顯地影響著群落結構和功能[15-17],也對草地群落植物物候有著不同程度的影響[18-20]。同時,物候作為氣候變化的“指紋”,表現出生態系統對環境變化的響應和適應,在一定程度上決定著群落水平的進化過程[21-23]。盡管如此,目前關于環境變化影響群落花期物候的機理尚不清楚[24-25]。
對青藏高原草地群落物候的研究不僅有助于深入認識草地群落構建機制,也有助于了解氣候變化(例如氮沉降)對其生態系統服務功能的影響,為預測和評估青藏高原高寒草地生態系統提供數據支撐。本研究基于高寒草甸6年氮添加試驗平臺,通過對不同氮添加處理下高寒草甸群落花期物候的觀測,研究不同功能群物種花期物候對氮添加的響應和主要組分種花期物候參數之間的關系,以及氮添加條件下高寒草甸花期物候和群落格局的動態變化,以期明確氮添加對高寒草甸植物花期物候和群落動態的影響。
1 材料與方法
1.1 研究區域概況
研究地位于蘭州大學高寒草甸與濕地生態系統定位研究站(瑪曲縣阿孜站,33°40′N,101°52′E),海拔約3 550 m,年均溫1.2℃(在生長季6月–8月最高溫度達到11.7℃,在非生長季11月?翌年4月溫度低至?10℃),年均降水量為620 mm(圖1)。年均日照時數約為2 580 h,年均霜降天數 >270 d;植被屬于高寒草甸類,主要優勢種有禾草類植物垂穗披堿草(Elymus nutans)、草地早熟禾(Poa pratensis)、胡氏剪股穎(Agrostis hugoniana),雜草類植物條葉銀蓮花(Anemone trullifolia)、鈍裂銀蓮花(Anemone obtusiloba)、絲葉毛茛(Ranunculus tanguticus)、小花草玉梅(Anemone rivularis)和瑞苓草(Saussurea nigrescens)等。研究樣地自2008年起圍封,僅冬季放牧。
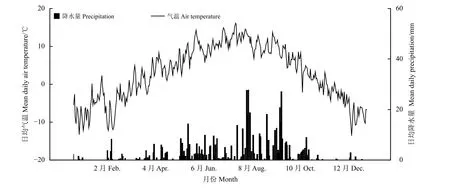
圖1 2014年研究地日均氣溫與降水量變化Figure 1 Mean daily temperature and daily precipitation of thestudy site in 2014
1.2 試驗設計
于2009年4月在圍欄封育的季節性放牧草地中設置15個10 m ×6 m 的樣區,各樣區間距為2 m。在每個樣區內,隨機設置3個氮添加處理小區,分別為對照(CK,0 g·m?2)、低氮處理(LN,5 g·m?2)和高氮處理(HN, 10 g·m?2),每個小區的面積為2.5 m ×6 m,緩沖帶為1 m。在每個小區內設置1個0.5 m ×0.5 m的固定樣方用于物候觀測。所用含氮試劑為硝酸銨(NH4NO3),每年5月初,為了確保氮肥能夠被植物有效吸收利用,并且保證氮肥隨流水的淋失作用降到最小,在多云的陰天將氮肥施加到相應的梯度處理樣方中[26]。
1.3 物候觀測及花期計算
于2014年整個生長季(4月中旬? 10月初),每6~7 d 記錄一次樣方內出現的物種及物候期,每次記錄獨立于上次進行,共觀測24次。本研究只關注始花期(first flowering date,FFD)和花期持續時間(flowering duration,FD)以及每次觀測時群落中處于花期的物種豐富度,每個觀測樣方中,當雜草類植物出現可辨認的花瓣、花藥和花絲時,記錄為始花期;當禾草類植物的小穗只要不再被葉鞘包裹時,記錄為始花期;當花瓣、花藥、花絲或萼片開始枯萎時,記錄為終花期[23,27],并以儒略歷(Julian day,1月1日為1年中第1天)計數法計時。物種的始花期和終花期是處于該物候期的物種出現在開花樣方中時間的平均值[28]。花期持續時間為對應樣方中終花期時間與始花期時間的差值。
1.4 數據分析
在3種氮添加處理水平樣方中均出現開花且在樣本數量滿足的情況下,以對照(CK)為基準,分別用低氮處理(LN)的數據和高氮處理(HN)的數據減去對照的數據,從而得到物種的花期提前與延遲的天數。用單因素方差分析(One-way ANOVA)檢驗氮添加對主要組分種和功能群(禾草類和雜草類)的始花期(FFD)和花期持續時間(FD)的影響。本研究中禾草類物種主要指禾本科物種,雜草類物種指除禾本科和豆科外的其他物種。用多重比較中的最小顯著差數法(LSD method)比較始花期和花期持續時間在不同氮添加水平之間的差異(P<0.05)。始花期和花期持續時間之間的關系采用Pearson 相關分析進行驗證。所有的分析和繪圖分別在SPSS 25.0和OriginPro 2016中進行。
2 結果與分析
2.1 群落主要組分物種和功能群花期物候對氮添加的響應
相對于對照,施氮處理明顯影響了群落中主要組分種的花期物候(圖2和圖3)。

圖2 低氮添加處理對高寒草甸25個常見物種始花期和花期持續時間的影響Figure 2 Effect of low nitrogen addition treatment on the first flowering date and flowering duration of 25 common alpine species

圖3 高氮添加處理對高寒草甸25個常見物種始花期和花期持續時間的影響Figure 3 Effect of high nitrogen addition treatment on the first flowering date and flowering duration of 25 common alpine species
低氮處理條件下,禾草類物種草地早熟禾、密花早熟禾(Poa poophagorum)、異針茅(Stipa aliena)、胡氏剪股穎、垂穗披堿草、羊茅(Festuca ovina)、芒剪股穎(Agrostis trinii)的始花期普遍性推遲(P<0.05);禾草類物種花期持續時間除垂穗披堿草顯著延長(P<0.05)外,其余物種花期持續時間均有一定縮短,其中羊茅的花期持續時間顯著縮短(P<0.05)。雜草類物種始花期和花期持續時間對施肥的響應因物種而異。菊科物種瑞苓草的始花期顯著推遲(P< 0.05),花期持續時間縮短;毛茛科物種鈍裂銀蓮花、高山唐松草(Thalictrum alpinum)、條葉銀蓮花、展毛翠雀花(Delphinium kamaonense)、絲葉毛茛的始花期顯著提前(P<0.05),而鈍裂銀蓮花和小花草玉梅的花期持續時間顯著延長(P<0.05);龍膽科物種濕生扁蕾(Gentianopsis barbata)、華麗龍膽(Gentiana sino-ornata)和橢圓葉花錨(Halenia elliptica),玄參科物種短腺小米草(Euphrasia regelii)和毛果婆婆納(Veronica eriogyne),傘形科物種細葉東俄芹(Tongoloa tenuifolia)和薔薇科物種莓葉委陵菜(Potentilla fragarioides)的始花期顯著提前(P< 0.05),其中細葉東俄芹、短腺小米草和毛果婆婆納的花期持續時間顯著延長(P<0.05)。
高氮處理條件下,禾草類物種除洽草(Koeleria cristata)和針茅(Stipa capillata)的始花期變化不顯著(P> 0.05)外,其余物種始花期均顯著推遲(P<0.05)。禾草類物種花期持續時間除垂穗披堿草外普遍縮短,其中異針茅和羊茅的花期持續時間顯著縮短(P<0.05),垂穗披堿草的花期持續時間顯著延長(P<0.05)。雜草類菊科物種瑞苓草的始花期顯著推遲(P<0.05),花期持續時間縮短;毛茛科物種鈍裂銀蓮花、絲葉毛茛、小花草玉梅、條葉銀蓮花和高山唐松草的始花期顯著提前(P<0.05),花期持續時間顯著延長(P<0.05);龍膽科物種濕生扁蕾、橢圓葉花錨和華麗龍膽的始花期顯著提前(P<0.05),花期持續時間變化各異;玄參科物種毛果婆婆納和短腺小米草,傘形科物種細葉東俄芹和薔薇科物種莓葉委陵菜的始花期顯著提前(P<0.05),花期持續時間顯著延長(P<0.05)。
氮添加總體上導致禾草類物種的始花期顯著推遲(LN和HN:P<0.01);雜草類物種始花期提前,其中在低氮處理水平下變化不顯著(P>0.05),而在高氮條件下,雜草類物種始花期顯著提前(P<0.05)(圖4a)。禾草類物種花期持續時間在高氮處理下顯著縮短(P<0.05);雜草類物種的花期持續時間在不同氮處理水平下均顯著延長(LN:P<0.05;HN:P<0.01)(圖4b)。
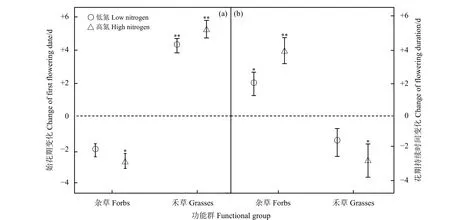
圖4 不同氮添加處理下功能群始花期和花期持續時間的變化Figure 4 Changes in the first flowering date and flowering duration of the different functional groups under the different nitrogen addition levels
相關性分析結果顯示(圖5),自然群落(r=?0.41,P=0.006 7)、低氮處理群落(r=?0.49,P=0.004 6)和高氮處理群落(r=?0.47,P=0.013 2)的物種始花期與花期持續時間之間表現出顯著的負相關性,表明物種開花時間越早,其花期持續時間越長,且在不同氮處理水平之間具有相同的趨勢。
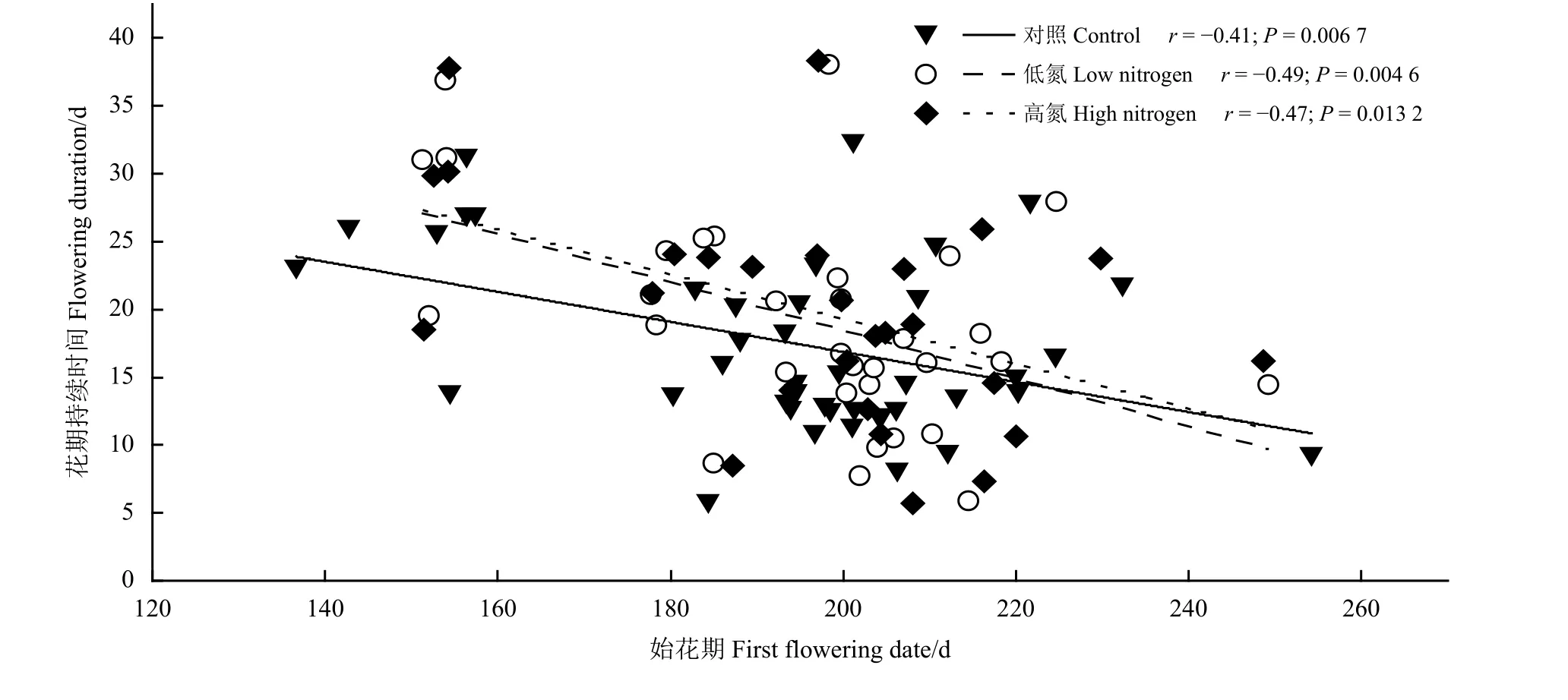
圖5 氮添加條件下物種始花期和花期持續時間的相關性Figure5 Relationship between the first flowering date and flowering duration of thedifferent nitrogen addition levels
2.2 氮添加后群落花期物種豐富度的變化
對試驗群落中開花物種物候觀測發現,群落中花期物種遍及整個生長季,群落種間花期在生長季內呈連續交錯趨勢并持續5個月時間。大部分物種花期集中在7月? 8月,其中在自然群落中觀測到49個物種開花,在低氮處理群落中觀測到35個物種開花,在高氮處理群落中觀測到25個物種開花。在3個處理中處于花期的物種多樣性在整個生長季內的變化都是迅速的,從4月下旬群落返青伊始便進入花期,以較快的速度在6月末7月初達到頂峰后迅速下降,直到10月初生長季結束。群落中處于花期的物種多樣性隨時間呈峰型分布(圖6)。在不同氮添加水平下,花期物種豐富度變化顯著,且隨著施氮量的增加群落中花期物種豐富度降低,每次觀測時群落中開花物種組成的相似性隨著施氮量的增加而降低,低氮群落和高氮群落中花期物種豐富度與自然群落相比分別降低了29%和49%。

圖6 生長季群落中花期物種豐富度對氮添加的響應Figure 6 Species richness of flowering response to nitrogen addition during growth season
3 討論
本研究表明,氮添加能夠顯著影響高寒草甸群落中25個主要組分種的花期物候,具體表現為禾草類物種始花期普遍推遲,開花持續時間縮短,而大部分雜草類物種始花期提前,花期持續時間延長。這一結果說明植物花期物候的變化除自身生活史特性決定外,環境因素變化也發揮著重要的作用。本研究中,物種花期物候的變化趨勢類似于北美一年生草地群落[29]和內蒙古草原群落中所研究的花期物候變化結果[30],但是與高原苔原群落中所研究的結果相反,該地區研究發現氮添加導致禾草類植物始花期提前,而雜草類植物始花期推遲[18]。這說明氮添加對植物物候的影響在一定程度上依賴于植物所處的生境類型。
在不同生境條件下,同一功能類型的植物可能使用不同的生長策略。大量研究發現,相較于雜草類,氮添加更加有利于禾草類植物的生長[31]。植物從營養生長到生殖生長的轉變是以生物量分配的變化和隨后營養攝取減少為特征[32]。在營養豐富的生境中,植物通過攝取更多的營養和花費更長的時間用于營養生長[33],對于晚期開花物種來說更是如此。通過增加營養生長,禾草類物種對限制性光源和氮的競爭更具有優勢,在施氮的樣方中占據更加明顯的優勢地位,從而表明在高寒地區禾草類物種是良好的氮競爭者[34]。大多數雜草類物種開花較早,這種早開花策略有助于它們避免與禾草類植物進行激烈的光資源競爭[35]。因此,在施氮條件下,于生長季早期大多數雜草類物種花期提前可能有利于它們在后期生長中避免激烈的光競爭。
本研究中,氮添加使得禾草類物種推遲開花,這意味著植物從營養生長到生殖生長轉變的推遲,在一定程度上表明植物加強營養生長或克隆生長[29]。相比之下,雜草類物種在氮添加條件下提前開花并延長花期持續時間,而開花持續期的延長意味著植物在花期時間生態位上有更多的時間吸引傳粉者或實現成功傳粉,從而有利于有性繁殖。相關性分析結果顯示,物種花期持續時間與始花期之間呈現顯著的負相關關系,這表明氮添加后,在生長季早期開花的物種較生長季中后期開花的物種來說,花期持續時間更長。高寒草甸群落中條葉銀蓮花、鈍裂銀蓮花以及高山唐松草等物種在生長季早期開花,即使它們能夠進行繁殖后營養生長,但是相對于生長季中后期開花物種的營養生長來說,其高度仍處于劣勢,所以早期開花物種為了避免在生長季后期資源競爭(如光、傳粉者等)中處于不利地位,采用了有利于其生殖生長的開花對策。本研究結果表明,在生長季前期物種始花期普遍提前,且花期持續時間延長,使得其能夠在繁殖階段充分利用環境中有限的營養資源。同時,由于在高寒地區生長季較短,晚開花物種受環境變化因素影響的風險增大(如霜降等)。因此早開花物種受限于營養生長,而遲開花物種通常受限于生殖生長。高寒草甸植物群落中大多數物種花期物候對氮添加的響應較為敏感,表現出不同的變化。由于群落中物種受生物、非生物以及其內在因素的影響,因此開花時間頻繁地處在多種選擇壓力之下[36-37]。
長期物候觀測研究表明,在融雪提前和冬季溫度升高條件下,大多數植物始花期顯著提前,這說明表型可塑性在植物開花過程中扮演著重要作用[38]。但是,也有研究發現植物物候過程主要由遺傳因素決定[33]。植物對環境因素(如光周期、降水、溫度、春化作用、融雪時間和土壤養分)的響應,引起開花時間的變化,是植物應對環境時空異質性變化的重要策略之一[39]。雖然氮是影響高寒草甸植物物候的重要環境因子,但是要詳細地闡明其對不同植物物種和功能群物候的影響,還需進一步觀察研究。同時,高寒草甸群落物種在時間生態位上的權衡,對于物種共存來說至關重要[40]。
群落中植物花期物候分布受多種環境因素共同作用[37],對環境變化十分敏感,而花期物候的變化可能會對生態系統結構與功能產生深遠的影響[41]。本研究通過對群落中物種整個生長季的物候觀測發現,物種間開花期呈現連續交錯趨勢,氮添加后群落中花期物種豐富度顯著降低,低氮群落和高氮群落的花期物種豐富度相較于自然群落分別降低了29%和49%。氮是影響草地群落物種生長的主要限制元素[42],氮添加導致環境異質性降低,從而影響植物的功能性狀和繁殖策略[43-44],當環境中氮含量超過一定值時則會造成物種多樣性降低,進而改變群落的組成結構和自然格局[45-46]。在高寒區域植物生長期短暫,群落物候的微小變化足以改變種間物候補償效應[47]。氮添加后群落蓋度和高度的增加可能會阻礙傳粉者的活動[48]。同時,傳粉者與其訪問的物種花期出現時間錯位時也會降低傳粉功能[49]。群落物種花期物候變化和群落水平花期物候分布格局的偏移會降低群落物候補償[50],形成的時間生態位分化會進一步影響群落的物種組成,進而導致群落結構和功能發生變化。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
小讀者(2021年2期)2021-03-29 05:03:48
華人時刊(2019年13期)2019-11-17 14:59:54
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
文苑(2018年22期)2018-11-19 02:54:14
汽車觀察(2018年10期)2018-11-06 07:05:26
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
