豬β-防御素的表達及其生物學功能研究進展
2021-08-09 02:00:36蘇國旗黃金秀劉作華
動物營養學報 2021年7期
關鍵詞:研究
蘇國旗 宋 陶 黃金秀 劉作華*
(1.重慶市畜牧科學院,重慶 402460;2.農業部養豬科學重點實驗室,重慶 402460;3.西南大學動物科學技術學院,重慶 402460)
近幾年,非洲豬瘟的爆發給我國養豬業帶來重創,同時也給畜牧生產者和研究者帶來啟示,對于健康養殖而言,需要外部減少感染的風險和內部提高動物機體的免疫力。2020年7月起我國開始禁止在飼料中使用抗生素,這就要求養殖生產中必須增強豬自身的免疫力,以減少由飼用抗生素禁用引起的發病率和死亡率;另外,還可通過開發新型綠色飼用抗生素替代品來緩解飼料禁用抗生素對養豬業帶來的負面影響。β-防御素是宿主先天免疫的重要組成部分,具有抗微生物活性,其主要作用機制是破壞細胞膜結構,且不易產生耐藥性,具有替代飼用抗生素的潛力。β-防御素還具有連接先天免疫和適應性免疫,減少感染、降低炎癥和提高免疫的功能,是提高動物機體抗病力的潛在位點。本文將就豬β-防御素(porcine β-defensins,PBD)的表達與分布、生物學功能、表達調控以及異源表達進行綜述,旨在為PBD的研究和應用提供參考。
1 PBD的表達與分布
防御素是一類含有3個二硫鍵的多肽,根據二硫鍵的連接方式不同可以分為α-防御素、β-防御素和θ-防御素[1]。α-防御素二硫鍵的連接方式是C1-C6、C2-C4和C3-C5,β-防御素二硫鍵的連接方式是C1-C5、C2-C4和C3-C6,θ-防御素二硫鍵的連接方式是C1-C4、C2-C5、C3-C6,連接形成環狀結構。θ-防御素在舊大陸猴和猩猩中有表達,但人和新大陸猴中不表達,人能夠表達α-防御素和β-防御素[2]。PBD1是被發現的豬的第1個β-防御素。它是用簡并引物從豬舌的cDNA庫中擴增獲得的,定位在豬的染色體15q14-q15.1[3]。通過生物信息學分析,目前已經發現了29種PBD[4]。這些PBD的氨基酸組成類似抗菌肽,富含帶正電荷的氨基酸,如賴氨酸和精氨酸。29種PBD基因主要定位在4個染色體上,分別是豬染色體SSC7、SSC14、SSC15和SSC17[4]。因為豬基因組不完整,所以PBD的數量并不確定,從目前研究結果來推測,可能與其他哺乳動物的類似,具有一系列的PBD[5]。系統發育分析發現PBD與人和牛的β-防御素有一定相似性,其中人和豬有27對直系同源β-防御素核酸序列的平均同源性達84.38%[4]。
PBD基因在機體的各個組織中廣泛表達,尤其是機體第1道防線組織(小腸和肺)及重要免疫器官(脾臟和胸腺)表達豐度比較高。其中PBD4、PBD122和PBD129基因在各個組織上的表達豐度都比較高(圖1)。豬的腎臟和睪丸廣泛表達多種PBD基因,另有研究表明PBD115基因在2周齡仔豬的睪丸上高表達,而在5月齡豬的睪丸上檢測不到,并且PBD115基因在肝臟上的表達與睪丸相反。PBD114和PBD133位點在同一染色體上并且相鄰,但它們的表達模式不同,而PBD4和PBD129位點在不同的染色體上,但它們的表達模式相似,說明PBD基因的表達是獨立的[4]。
2 PBD的生物學功能
2.1 抗微生物活性
研究表明,β-防御素具有抗細菌和抗病毒的活性,本文主要就PBD1和PBD2的抗微生物活性進行綜述(表1)。不同來源的PBD1都表現出了抗細菌和抗病毒活性,例如:化學合成的PBD1具有抗大腸桿菌和枯草芽孢桿菌活性[6],內源表達的PBD1具有抗百日咳博德氏菌[7]和丙型肝炎病毒活性[8],大腸桿菌異源表達的PBD1具有抗大腸桿菌、金黃色葡萄球菌、傷寒沙門氏菌、單核增生李斯特菌和白色念珠菌活性[9-10]。PBD2的來源要多于PBD1,除了化學合成、內源表達和大腸桿菌異源表達外,PBD2的來源還有枯草芽孢桿菌異源表達[11]、畢赤酵母異源表達[12]和轉基因豬過表達[13-14]。化學合成的PBD2具有抗假單胞菌[6]、鼠傷寒沙門氏菌[15]、偽狂犬病病毒[16-17]和豬繁殖和呼吸綜合征病毒活性[18],由大腸桿菌異源表達的PBD2具有抗金黃色葡萄球菌和大腸桿菌活性,由枯草芽孢桿菌和畢赤酵母異源表達的PBD2的抗菌譜更廣,具有抗鼠傷寒沙門氏菌、大腸桿菌、副嗜血桿菌、金黃色葡萄球菌、表皮葡萄球菌、枯草桿菌和小膿皰桿菌活性[11-12]。另外PBD2轉基因豬也表現出抗A.pleuropneumoniae[13]和副豬格雷氏菌感染的能力[14]。除了PBD1和PBD2外,另有少數PBD的抗微生物活性有過研究,例如:內源表達的PBD3具有抑制豬繁殖和呼吸綜合征病毒感染的活性[19],大腸桿菌異源表達的PBD114有微弱的抗大腸桿菌DH5α活性[20],大腸桿菌異源表達的PBD129也有微弱的抗大腸桿菌DH5α和異乳鏈球活性[21]。
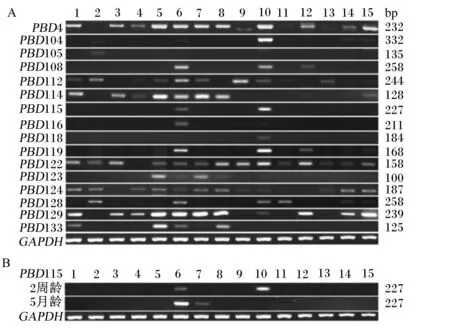
A:2周齡仔豬不同組織中PBD基因的表達豐度;B:2周齡和5月齡豬睪丸中PBD115基因表達豐度的變化。1:小腸;2:舌;3:眼睛;4:腦;5:脾臟;6:腎臟;7:肝臟;8:肺;9:胃;10:睪丸;11:肌肉;12:皮膚;13:直腸;14:氣管;15:胸腺。PBD:豬β-防御素基因;GAPDH:甘油醛-3-磷酸脫氫酶基因。
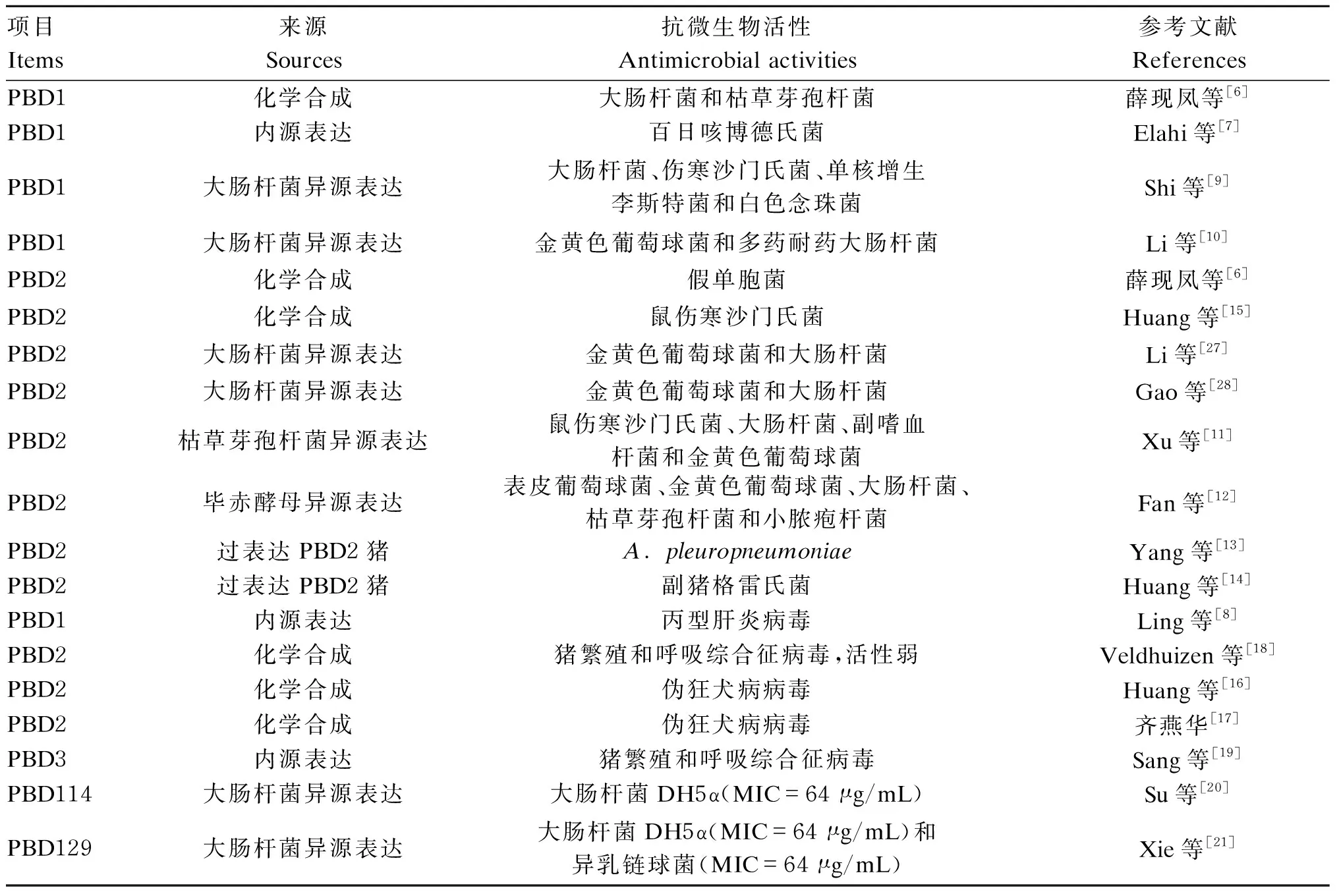
表1 PBD的抗微生物活性
抗菌肽的殺菌機制包括胞內和破膜兩大方式,抗菌肽對胞內的靶點包括DNA、RNA和蛋白質等,破膜方式也有聚集體模型、環孔模型、桶板模型和地毯模型[22-23]。防御素是有3個二硫鍵的抗菌肽,具有抗菌肽的理化特性。透射電鏡結果顯示,PBD1和PBD2可通過跨膜孔的形成殺滅細菌[6]。經PBD2處理的鼠傷寒鏈球菌的透射電鏡顯示,相對低劑量的PBD2會導致細胞質膜收縮,而接近最小抑菌濃度(MIC)的PBD2會導致細胞質泄漏,細菌細胞完全裂解[18],這就表明PBD具有破壞病原菌細胞膜殺菌的機制。進一步的研究表明,PBD2主要定位到金黃色葡萄球菌的細胞膜,使細胞膜破裂、膨脹和穿孔,同時有少量直接進入細胞質。PBD2也能與涉及到跨膜運輸、可遺傳信息運輸和其他代謝過程的DNA反應,最終殺死金黃色葡萄球菌[24]。PBD2可通過多種作用模式殺死大腸桿菌,主要機制是通過靶向細胞膜損傷后的細胞內分子來影響DNA轉錄和翻譯,同時也會影響轉運和代謝蛋白質[25]。有研究用精氨酸和賴氨酸取代了PBD2的8個氨基酸殘基,這些突變蛋白對大腸桿菌和金黃色葡萄球菌表現出不同的抗菌活性,表明這些帶正電荷的殘基并不是影響其抗微生物活性的唯一因素,其他因素如這些殘基在防御素表面的分布也可能影響其抗微生物活性[26]。
2.2 免疫調節功能
隨著對防御素抗微生物活性研究的不斷深入,人們發現防御素的抗微生物活性在生理鹽水濃度下或者是在宿主其他生理條件下嚴重受損,過去20年的研究表明,防御素除了具有抗微生物活性外,還具有免疫調節功能,這有助于其抗微生物活性的發揮[29-31]。防御素具有多種多樣的免疫調節功能,通過不同的機制直接作用于特定的細胞,可以誘導或抑制炎癥反應,還可以與包括蛋白質、核酸和碳水化合物在內的細胞分子形成復合物來調節免疫反應[31]。然而對PBD免疫調節功能的研究比較匱乏且不夠深入,主要研究了PBD的抗氧化能力、腸道屏障功能和抗炎活性。薛現鳳[32]研究表明,PBD1和PBD2能夠清除1,1-二苯基-2-三硝基苯肼自由基和羥基自由基,表現出一定的抗氧化能力。腸道黏膜屏障可以直接阻擋腸道病原微生物對宿主的感染,是保護機體的重要屏障,緊密連接蛋白和黏蛋白有助于維持腸道黏膜屏障的完整性,保持腸道正常生理功能。研究表明,PBD1和PBD2能夠增加腸道緊密連接蛋白——閉合小帶蛋白-1(ZO-1)、閉合小帶蛋白-2(ZO-2)和封閉蛋白-1(Claudin-1)以及黏蛋白——黏蛋白1(Mucin1)、黏蛋白2(Mucin2)和黏蛋白20(Mucin20)基因或蛋白的表達[32-33],PBD129同樣也能增強腸道緊密連接蛋白的表達[21]。在Caco-2細胞中,PBD2增加了跨上皮電阻和黏蛋白基因的表達,降低了細胞的通透性和凋亡,減少了結腸上皮的炎癥浸潤,同時保持了緊密連接的結構完整性[33]。過量的炎癥因子會降低腸上皮細胞緊密連接蛋白的表達,破壞腸黏膜屏障的完整性。將人工合成的PBD2添加到豬飼糧中,能夠降低空腸黏膜Toll樣受體4(TLR4)、腫瘤壞死因子-α(TNF-α)、白細胞介素-1β(IL-1β)、白細胞介素-8(IL-8)基因的表達[34]。而PBD129可通過降低小鼠血清中炎性細胞因子IL-1β、白細胞介素-6(IL-6)和TNF-α的濃度來緩解脂多糖誘導的炎癥反應,高劑量PBD129顯著降低脂多糖誘導小鼠的血清二胺氧化酶(DAO)活性,通過可調節凋亡相關基因[B淋巴細胞瘤-2基因(Bcl-2)和Bcl-2相關X蛋白基因(Bax)]和蛋白[半胱天冬酶-3(Caspase-3)和半胱天冬酶-9(Caspase-9)],降低腸上皮細胞凋亡[21]。體內試驗研究表明PBD2能夠降低硫酸葡聚糖鈉(DSS)誘導的小鼠結腸炎性細胞因子(IL-8、IL-6和TNF-α)的基因表達和蛋白質翻譯,可能是PBD2通過抑制蛋白激酶B(AKT)和核因子-κB(NF-κB)抑制蛋白(IκBα)的磷酸化,抑制NF-κB的磷酸化,從而抑制炎癥調節分子誘導型一氧化氮合酶(iNOS)和環氧合酶-2(COX-2)的基因表達,以及降低一氧化氮(NO)和前列腺素E2(PGE2)的濃度來降低炎癥[33]。
3 PBD的表達調控
3.1 病原微生物
動物感染病原微生物會引起免疫反應,首先是與病原微生物直接接觸的先天免疫,可通過物理、化學和分子屏障阻止病原微生物入侵動物機體。研究發現病原菌(鼠傷寒沙門氏菌、大腸桿菌、鏈球菌和牛型結核桿菌)能夠誘導PBD1、PBD2、PBD114和PBD129基因的表達[20,35-39],不同病原菌特異性地誘導相應的PBD基因的表達,例如:鼠傷寒沙門氏菌能夠誘導PBD1基因表達,但是不能誘導PBD2基因表達[35],并且只有活的鼠傷寒沙門氏菌才能誘導PBD1基因表達。但是當把鼠傷寒沙門氏菌DT104灌注到生理狀況更復雜的小腸中時,就不能誘導PBD1和PBD2基因表達[40]。病原體同樣也能抑制PBD基因的表達,豬肺炎支原體通過阻斷展開蛋白反應來抑制PBD2的產生,從而促進其在上皮細胞的黏附和感染[41]。
3.2 益生菌
益生菌是有益于宿主的微生物,能夠抑制腸道有害菌群的定植,改善腸道菌群結構,從而減少疾病,提高畜禽生產性能[42-43]。近些年的研究表明,益生菌能夠以非炎癥方式上調PBD基因的表達,從而起到平衡腸道微生物的作用。例如:羅伊氏乳桿菌能夠誘導IPEC-J2細胞的PBD2、PBD3、PBD114和PBD129基因表達[43];植物乳桿菌能夠誘導哺乳仔豬空腸PBD2和PBD3以及回腸PBD1、PBD2和PBD3基因表達[44];植物乳桿菌ZLP001和羅伊氏乳桿菌LR1能夠誘導PBD2基因表達[45-46]。羅伊氏乳桿菌誘導IPEC-J2細胞表達PBD基因具有時間效應,處理6 h時表達量最高,12 h后開始下降,并且PBD1基因表達基本不受羅伊氏乳桿菌的影響[43]。體內試驗不同于體外試驗,羅伊氏乳桿菌不誘導回腸PBD基因表達,但羅伊氏乳桿菌顯著提高新生仔豬空腸PBD2以及結腸PBD2、PBD3、PBD114和PBD129基因表達,這種變化并不是由于羅伊氏乳桿菌的直接刺激造成的,而是通過增加結腸中丁酸的濃度,上調過氧化物酶體增殖物活化受體γ(PPARγ)和G蛋白偶聯受體41(GPR41)基因的表達來調節腸道β-防御素的表達并改善腸道健康[47]。益生菌對PBD基因表達的誘導不僅具有時間效應,還具有劑量效應。Zhang等[48]研究發現,唾液酸桿菌B1在十二指腸黏膜早期定植,唾液中的PBD2基因表達水平隨著腸上皮細胞暴露于唾液中而增加,并且與它的劑量呈正相關。正常生理狀態下益生菌能夠以非炎癥方式上調PBD基因的表達;而當機體受到病原微生物感染時,益生菌又能降低由炎癥方式誘導的PBD基因的表達,從而避免免疫過度激活對機體造成的損傷[49]。
3.3 脂肪酸
丁酸是一種揮發性短鏈脂肪酸,能夠給腸上皮細胞提供能量,同時還具有調節免疫功能的作用。研究發現,丁酸能夠顯著誘導IPEC-J2細胞、3D4/31細胞和原代單核細胞的PBD2和PBD3基因表達,并且在斷奶仔豬小腸上皮細胞上得到同樣的結果,但是不影響PBD1基因表達[50-51]。丁酸鈉能夠誘導豬腎上皮細胞PK-15表達更多的PBD基因,能夠強烈誘導PBD3、PBD128、PBD123和PBD115基因表達[52]。丁酸鈉有一系列的類似物,包括甘油三丁酸、丁酸芐和4-苯基丁酸等,研究發現,它們對PBD基因表達的誘導作用與丁酸鈉的相當[50,53]。中鏈脂肪酸辛酸和壬酸能夠誘導IPEC-J2細胞的PBD1和PBD2基因表達,但是誘導強度要低于丁酸[50,54],膠囊包被棕櫚油與未包被棕櫚油相比可顯著提高仔豬腸道PBD2和PBD129的分泌[55]。
3.4 氨基酸
近些年,氨基酸的功能性備受關注,尤其是氨基酸的免疫調節功能。研究表明,L-異亮氨酸能夠誘導IPEC-J2細胞的PBD1、PBD2和PBD3基因表達,并且具有時間和劑量效應[56-58]。任曼等[57]研究了3種支鏈氨基酸和丙氨酸對IPEC-J2細胞的PBD基因表達的影響,發現對PBD基因表達誘導最強的是異亮氨酸,且3種支鏈氨基酸在誘導PBD基因表達時具有協同效應。精氨酸在機體內不僅能夠參與鳥氨酸循環,還有重要的免疫調節功能。飼糧中添加L-精氨酸能夠增加仔豬PBD1、PBD2和PBD3基因表達,也能增加間歇熱處理的育肥豬PBD2基因表達[59];體外研究也得到類似的結果,L-精氨酸能提高豬胎兒皮膚纖維細胞(PFSF)的PBD1、PBD2和PBD3基因表達[60]。
3.5 其他
除了微生物、脂肪酸和氨基酸能夠誘導PBD基因表達外,微量元素[56]、維生素[61]、功能性添加劑[62]和毒素[63]也能誘導PBD基因的表達,不同品種[64-65]和不同日齡[4]豬的PBD基因表達也不同。例如:Mao等[56]的研究表明,IPEC-J2細胞經過25或100 μmol/mL的硫酸鋅處理12 h后,提高了PBD1、PBD2和PBD3的基因和蛋白表達;陳金永[61]研究發現,維生素A、維生素D和維生素E都能夠調控PBD1、PBD2和PBD3基因的表達;染料木素和大豆苷元是大豆異黃酮的主要活性物質,能夠誘導豬子宮內膜上皮細胞中PBD2基因表達,以及PBD2和PBD3分泌[62];嘔吐毒素顯著降低IPEC-J2細胞的PBD1和PBD3基因表達[63];抗病力高的地方品種豬,PBD基因的表達水平高于杜-長-大三元雜交豬[20,61,64-65]。
3.6 PBD基因表達的調控機制
綜上所述,能夠誘導PBD基因表達的因素很多,而不同因素對不同PBD基因的表達調控機制也有所不同,這就使PBD基因的表達調控機制變得非常復雜(圖2)。研究發現丁酸能夠上調PPARγ和GPR41基因的表達來調節腸道β-防御素的表達[47]。體外試驗發現丁酸鈉能夠激活NF-κB誘導腎臟上皮細胞表達PBD基因,抑制絲裂原活化蛋白激酶(MAPK)信號通路能阻止丁酸鈉對PBD基因表達的誘導[23]。苯甲酸鈉通過激活NF-κB信號通路誘導腸上皮細胞表達PBD1和PBD3基因,并且這一過程依賴于Toll樣受體2(TLR2)和TLR4[24]。辛酸和壬酸能夠促進了組蛋白H3賴氨酸9(H3K9)在PBD1和PBD2啟動子上的乙酰化,從而增加了PBD1和PBD2基因的表達[54]。任曼等[57]研究發現,抑制細胞外信號調節激酶(ERK)會降低β-防御素的表達,并且降低了由異亮氨酸誘發的ERK1/2和90 ku核糖體S6蛋白激酶(90RSK)的磷酸化,同時抑制沉默信息調節因子2相關酶1(Sirt1)信號通路降低異亮氨酸對β-防御素表達的促進作用,說明異亮氨酸能夠通過Sirt1/ERK/90RSK信號通路調控IPEC-J2細胞的PBD基因表達。另外,Lan等[66]研究發現,L-精氨酸能夠通過哺乳動物雷帕霉素靶蛋白(mTOR)信號通路增加IPEC-J2細胞的PBD基因表達。維生素A對不同的防御素的調控機制有所不同,維生素A通過NF-κB信號通路調控PBD2基因表達,通過絲裂原活化細胞外信號調節激酶(MEK)-ERK信號通路調控PBD1和PBD3基因表達[61]。

VA:維生素A vitamin A;TLR2:Toll樣受體2 Toll-like receptor 2;TLR4:Toll樣受體4 Toll-like receptor 4;PPARγ:過氧化物酶體增殖物活化受體γ peroxisome proliferator-activated receptor γ;GPR41:G蛋白偶聯受體41 G protein-coupled receptor 41;ERK:細胞外信號調節激酶 extracellular signal-regulated kinase;MEK:絲裂原活化細胞外信號調節激酶 mitogen-activated extracellular signal-regulated kinase;mTOR:哺乳動物雷帕霉素靶蛋白 mammalian target of rapamycin;90RSK:90 ku核糖體S6蛋白激酶 90 ku ribosomal S6 kinase;H3K9:組蛋白H3賴氨酸9 histone H3 lysine 79;NF-κB:核因子-κB nuclear factor-κB;IκBα:核因子-κB抑制蛋白 inhibitor α of nuclear factor-κB;PBD:豬β防御素 porcine β-defensins;Degraded:被降解的。
4 PBD制備與應用
蛋白的生產方式主要有人工合成、直接提取和異源表達,目前用于研究的PBD主要是人工合成和異源表達。但是由于PBD氨基酸序列比較長,并且還有3個二硫鍵,合成難度比較大,因此異源表達是制備PBD經濟有效的方式。用于制備PBD的異源表達方式主要是大腸桿菌,其次是畢赤酵母和枯草芽孢桿菌。表2總結了PBD的異源重組表達系統及其重組蛋白抗菌活性,發現畢赤酵母表達的PBD的抗菌譜要更廣。通過重組枯草芽孢桿菌工程表達PBD2,在豬飼糧中添加該工程枯草芽孢桿菌可顯著促進仔豬生長,降低腹瀉發生率[11]。Tang等[34]使用合成的PBD2作為抗菌生長促進劑添加到飼糧中,發現每天攝入1 mg的PBD2可以提高仔豬的采食量、日增重以及空腸絨毛高度/隱窩深度,促進胰島素樣生長因子的表達,減少直腸中大腸桿菌、脆弱擬桿菌和鏈球菌的數量,并抑制促炎因子IL-1β、IL-8、TNF-α基因的表達。Peng等[67]使用畢赤酵母重組的PBD2替代抗生素應用于斷奶仔豬,發現飼糧中添加5 g/kg的PBD2可以增加仔豬的采食量、日增重、十二指腸和空腸絨毛高度,并減少盲腸中致病菌的數量,降低腹瀉發生率。因此,PBD2應用于仔豬生產中具有抗菌作用,可改善仔豬的生長性能,降低炎癥細胞因子的表達,改善腸道形態指標,具有開發成飼用抗生素替代品的潛力。

表2 PBD的異源表達
5 小 結
PBD是豬重要的先天免疫分子,在豬的各個組織中廣泛表達和分布。PBD表現出抗菌和抗病毒的活性,主要是通過破壞病原菌細胞膜,以及進入細胞影響DNA轉錄和蛋白質翻譯來實現。PBD具有免疫調節功能,通過NF-κB信號通路降低宿主炎癥,提高腸道緊密連接蛋白表達,改善腸道健康。微生物能夠通過TLR2/TLR4-NF-κB/MAPK信號通路誘導PBD基因的表達抑制病原菌的感染,而營養素(脂肪酸、氨基酸和微量元素)能夠通過Sirt1/ERK/90RSK+mTOR信號通路調節PBD基因的表達提高宿主的抗病力。在仔豬的應用中,PBD能提高生長性能,降低腹瀉,具有替代飼用抗生素的潛力。然而,目前研究主要集中于PBD1和PBD2,對其他27種PBD的研究十分匱乏。對PBD仍需大量的基礎性研究,其中營養素靶向調控PBD的內源表達和PBD免疫調節功能是未來研究的重點。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
