龍珠果莖段離體培養和組培苗耐鹽性分析
2021-08-10 02:22:00劉艷艷司燦何春梅俞振明段俊
熱帶亞熱帶植物學報 2021年4期
關鍵詞:生長
劉艷艷, 司燦, 何春梅, 俞振明, 段俊*
龍珠果莖段離體培養和組培苗耐鹽性分析
劉艷艷1,2, 司燦2, 何春梅2, 俞振明2, 段俊2*
(1. 廣州花卉研究中心, 廣州 510360; 2. 中國科學院華南植物園, 廣東省應用植物學重點實驗室, 中國科學院華南農業植物分子分析與遺傳改良重點實驗室, 廣州 510650)
為建立龍珠果()的快繁再生體系,以實生苗莖段為外植體,研究了植物生長調節劑對叢生芽誘導、壯苗生根的影響,同時對組培苗的耐鹽性進行研究。結果表明,MS+6-BA 0.5 mg/L+NAA 0.05 mg/L培養基有利于誘導叢生芽并促進芽的生長;MS+6-BA 3.0 mg/L+NAA 0.3 mg/L培養基有利于誘導愈傷組織;1/2 MS+IBA 0.2mg/L培養基適合小芽壯苗生根。組培苗移栽至泥炭土∶蛭石∶珍珠巖(2∶1∶1)的基質中,成活率可達92.6%,且植株生長良好。0~200 mmol/L NaCl處理的組培苗生長不受影響;超過200 mmol/L NaCl處理,植株出現矮化、葉片萎蔫、變黃等現象。隨NaCl濃度升高,葉片的SOD活性逐漸升高,POD、CAT和APX活性則呈先升高后降低的趨勢。這為龍珠果的種苗繁育、海濱生態修復提供了技術支持。
龍珠果;莖段;叢生芽;愈傷組織;鹽脅迫
龍珠果(),別名香花果、天仙果、野仙桃等,為西番蓮科(Passifloraceae)西番蓮屬草質藤本植物。原產于西印度群島,在我國主要分布于廣東、海南、福建和臺灣等地,生長于荒山草坡、灌叢或近海邊沙灘[1]。龍珠果花朵秀麗,顏色多變;果實外形美觀,香甜可食[2]。
龍珠果莖桿和葉片中含有黃酮類化合物,主要成分為牡荊素、異牡荊素、葒草素,異葒草素等[3]。龍珠果具有清熱解毒、清肺止咳的作用,可用于治療肺熱咳嗽、小便混濁、癰瘡腫毒、外傷性眼角膜炎、淋巴結炎等癥[4–5]。果實可用作催吐劑,煎煮后可用于治療哮喘;葉片外敷可治療創傷,干燥葉片泡茶有助于提高睡眠質量;根可用于調經,對治療過度興奮和暴躁有作用[6]。
龍珠果的自然繁殖方式是種子繁殖,但種子萌發率低、繁殖周期長。關于龍珠果組織培養技術, 國內尚未見文獻報道。本文以龍珠果帶節莖段為外植體,通過調整植物生長調節劑的種類和濃度,探討適合叢生芽誘導、壯苗生根、煉苗移栽等階段的培養基,為龍珠果直接器官發生體系的建立提供試驗依據。龍珠果作為一種海濱植物,多生長于高鹽環境,組培苗的耐鹽程度是其能否在海濱環境成活的重要指標,通過對2月齡組培苗進行鹽脅迫處理,探討其對鹽脅迫的適應程度。
1 材料和方法
1.1 材料
龍珠果()種子發芽后種植在中國科學院華南植物園生物技術育種實驗室。3個月后苗高約12 cm,選取生長健壯、長勢基本一致的植株,截取帶節的莖段用于組織培養。取移栽至基質中生長2個月長勢基本一致的組培苗用于鹽脅迫試驗。
1.2 組織培養
以MS為基本培養基[7],pH 5.7。培養條件: (25± 2)℃、光照強度32.57mol/(m2·s)、12 h光照/12 h黑暗。將清洗過的龍珠果莖段置于超凈工作臺上, 用75%酒精浸泡約30 s,無菌水清洗2~3次,然后用0.1% HgCl2浸泡5 min,最后無菌水沖洗4~5次。將消毒過的莖段切成約2 cm帶節點小段,插入在已滅菌的培養基中。
植物生長調節劑組合的影響 將帶節莖段接種于添加不同質量濃度的6-BA (0.5、1.0、2.0、3.0 mg/L)與IBA (0.05、0.1、0.2、0.3 mg/L)或NAA (0.05、0.1、0.2、0.3 mg/L)組合的MS培養基中, 以不添加任何植物生長調節劑的MS培養基為對照。每處理接種5瓶,每瓶15個莖段。40 d后統計叢生芽誘導率、芽分化率、愈傷組織誘導率等。叢生芽誘導率(%)=誘導出芽或芽點的莖段數/接種的莖段總數×100%,芽分化率(%)=分化出芽的數量/芽點的總數×100%,愈傷組織誘導率(%)=誘導出愈傷組織的莖段數/接種的莖段總數×100%。
壯苗生根 將誘導出的叢生芽分離,接種于含不同質量濃度的NAA (0.05、0.1、0.2 mg/L)和IBA (0.05、0.1、0.2 mg/L)的1/2MS培養基中進行壯苗生根培養,以1/2MS培養基為對照。每處理接種8瓶,每瓶3個小芽。40 d后統計生根率、根長、根粗等。生根率(%)=誘導出根的芽數量/接種芽的總數×100%。
煉苗移栽 組培苗長至8~12 cm高時,選擇長勢旺盛、葉片較多且根系發育良好的組培苗,打開瓶蓋,轉移到自然條件下煉苗1周。小心從組培瓶中取出幼苗,用自來水將根部沖洗干凈,移栽至泥炭土∶蛭石∶珍珠巖為2∶1∶1的基質中,共移栽121株。基質澆透水,同時提高空氣濕度,保持適當通風,30 d后統計移栽成活率。移栽成活率(%)=成活苗數/移栽總苗數×100%。
1.3 鹽脅迫試驗
組培苗移栽2個月后,隔天用不同濃度的NaCl溶液澆灌,共設6個處理:0、100、200、400、600、800 mmol/L,以0 mmol/L NaCl為對照,每處理5株。5 d后每處理分別稱取0.5 g功能葉片(從上向下第二片),凍存于-80℃冰箱中,3次重復。
SOD、POD、CAT和APX活性測定 取出葉片置于預冷的研缽中,加入少量的石英砂和5 mL 0.05 mol/L的磷酸緩沖液(pH 7.8,1% PVP, 0.1 mmol/L EDTA),在冰上迅速研磨成均一渾濁的懸浮液,轉入15 mL離心管中。在4℃,7 104×離心10 min,取上清液,4℃放置。SOD活性采用核黃素-NBT法[8]測定;POD活性采用愈創木酚法[9–10]測定;CAT活性采用紫外分光光度計法[11]測定; APX酶活性采用抗壞血酸氧化法[12]測定。
1.4 數據統計
采用Excel 2010對數據進行整理和統計,顯著性分析采用SPSS 22.0中的鄧肯氏多重檢測法(Dun- can’s multiple range test, DMRT), 以<0.05表示差異顯著。
2 結果和分析
2.1 植物生長調節劑的影響
莖段接種到誘導培養基后定期觀察,約15 d在節點處開始長出芽點或小芽,隨后莖段兩端慢慢變黃,培養40 d后統計叢生芽誘導率、芽分化率、愈傷組織誘導率等指標(表1和圖1: A)。
培養基中添加0.5 mg/L 6-BA+0.05 mg/L IBA,誘導的叢生芽中有14.6%芽長可達4 cm;隨6-BA、IBA濃度的升高,叢生芽誘導率沒有顯著變化,芽分化率下降,小芽(1 cm長)比例增加。當6-BA濃度為3.0 mg/L、IBA濃度為0.3 mg/L時,芽分化率為2.3%,誘導出的都是小芽。這表明6-BA與IBA組合能誘導出叢生芽或芽點,且隨濃度升高芽分化率降低,小芽比例升高。
培養基中添加0.5 mg/L 6-BA+0.05 mg/L NAA,叢生芽誘導率為98.7%,4 cm大芽達9.9%;隨著6-BA、NAA濃度的升高,叢生芽誘導率和芽分化率顯著下降,小芽比例明顯增加,從莖段誘導出有白色愈傷層的淺綠色愈傷組織(圖1: B),且愈傷組織的數量隨6-BA、NAA濃度的升高而增多。當6- BA濃度為3.0 mg/L、NAA濃度為0.3 mg/L時, 叢生芽誘導率下降為40%,誘導出的全是1 cm小芽,愈傷組織誘導率為59.5%。這表明較低濃度的6-BA與NAA組合有利于莖段誘導叢生芽和芽的分化,較高濃度的6-BA與NAA組合有利于莖段誘導產生愈傷組織。
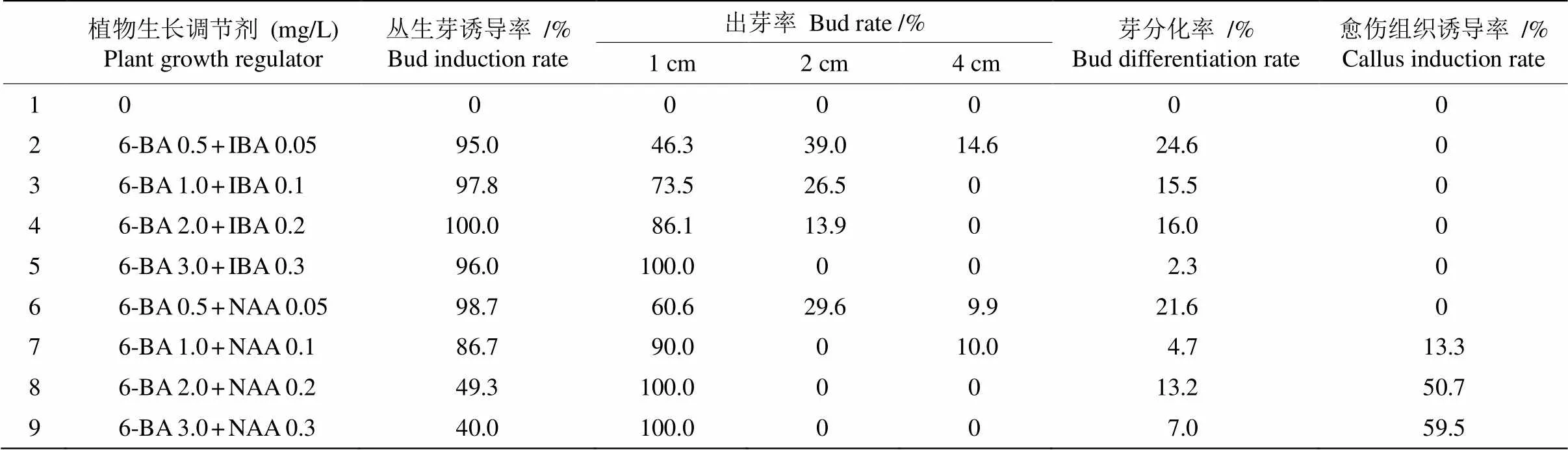
表1 植物生長調節劑對叢生芽誘導的影響

圖1 龍珠果莖段的離體培養和植株再生。A: 叢生芽誘導; B: 愈傷組織誘導; C: 小芽; D: 生根; E: 移栽30 d; F: 移栽90 d。
因此,MS+6-BA 0.5 mg/L+NAA 0.05 mg/L培養基有利于龍珠果莖段誘導叢生芽并促進芽的生長, 而MS+6-BA 3.0 mg/L+NAA 0.3 mg/L培養基有利于莖段誘導愈傷組織。
2.2 植物生長調節劑對壯苗生根的影響
將大小均勻一致的叢生芽(圖1: C)轉接到壯苗生根培養基后定期觀察,約15 d根開始形成,培養40 d后對根的生長情況進行統計、分析(表2)。
培養基中添加0.05 mg/L NAA, 生根率為100%, 29.2%的根長達5 cm,根細,植株生長正常;當NAA為0.2 mg/L時,生根率為83.3%,10.0%的根長達5 cm,根略粗,植株葉片較黃。這表明低濃度NAA有利于誘導分化芽長根,但根細長;高濃度NAA對根的誘導有抑制作用,但根較粗,葉片較黃。
培養基中添加0.05 mg/L IBA,生根率為54.2%,38.5%的根長達5 cm,根略粗,植株正常生長;當IBA為0.2 mg/L時,生根率為95.8%,73.9%的根長達5 cm,根略粗,葉片較綠(圖1: D)。這表明, 低濃度IBA誘導生根的效果較差,根短粗;而高濃度IBA有利于誘導生根,根長且粗,植株葉片較綠。
當NAA與IBA配合使用時,隨濃度升高,生根率反而下降,但根較粗,植株葉片較黃。因此, 培養基1/2MS+IBA 0.2 mg/L適合龍珠果小芽的壯苗生根培養。
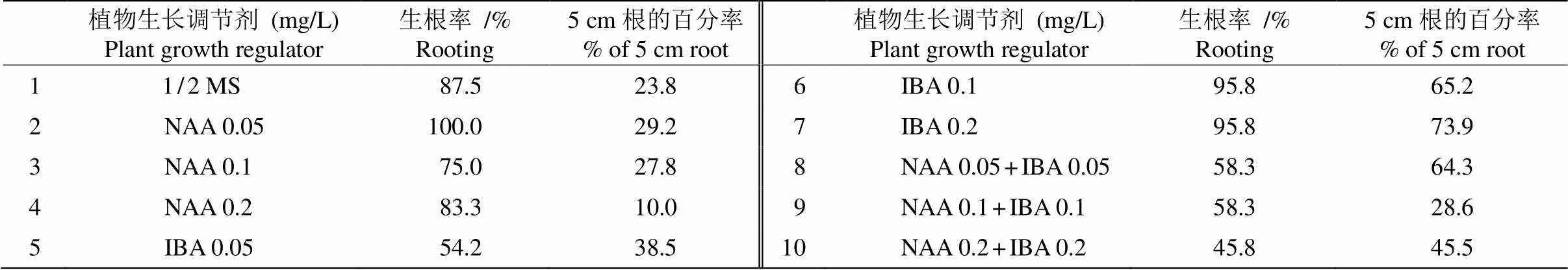
表2 植物生長調節劑對壯苗生根的影響
2.3 煉苗移栽
待組培瓶苗長到8~12 cm高時,打開瓶蓋置于自然條件下煉苗,7 d后將組培苗取出,洗凈根部培養基,移栽至泥炭土∶蛭石∶珍珠巖為2∶1∶1的基質中。移栽后定期補充水分,30 d后(圖1: E)龍珠果成活率達92.6%,90 d后植株長勢良好(圖1: F)。
2.4 鹽脅迫的影響
鹽脅迫對龍珠果組培苗的生長總體表現為抑制作用,隨NaCl濃度的升高,植株出現矮化現象, 葉片逐漸萎蔫卷曲,顏色變黃(圖2: A)。隨NaCl濃度的升高,株高逐漸降低(圖2: B)。當NaCl濃度< 200 mmol/L時,株高與對照的差異不顯著(>0.05);NaCl濃度>400 mmol/L時,株高顯著低于對照(<0.05)。表明龍珠果組培苗能適應200 mmol/L以下的NaCl脅迫。

圖2 鹽脅迫下龍珠果組培苗的表型(A)和株高(B)變化。n=3; 柱上不同字母表示差異顯著(P<0.05)(DMRT)。
從圖3可見,龍珠果組培苗葉片的SOD活性隨NaCl濃度升高呈逐漸升高的變化趨勢,當NaCl濃度大于400 mmol/L時,SOD活性顯著升高(<0.05)。NaCl濃度為0~400 mmol/L時,POD活性較低,當NaCl濃度大于400 mmol/L時,POD活性顯著升高(<0.05)。CAT活性隨NaCl濃度升高呈先升后降的變化趨勢,0~200 mmol/LNaCl處理時,CAT活性逐漸升高,以200 mmol/L NaCl處理的最高,隨后(200~800 mmol/L NaCl) CAT活性逐漸降低。APX活性隨NaCl濃度升高呈先升后降的變化趨勢,低濃度NaCl (0~400 mmol/L)處理的APX活性總體升高,以400 mmol/L NaCl處理的最高,隨后(400-800 mmol/L NaCl) APX活性逐漸降低。

圖3 鹽脅迫下龍珠果葉片的SOD、POD、CAT和APX活性
3 結論和討論
6-BA與NAA或IBA組合誘導莖段產生叢生芽在很多植物中都有報道[13–15],本試驗結果表明,6-BA與NAA組合誘導龍珠果莖段產生叢生芽的效果較好,且較低濃度的6-BA與NAA組合有利于莖段誘導叢生芽和芽的生長;較高濃度的6-BA與IBA組合不能使龍珠果莖段產生愈傷組織,而較高濃度的6-BA與NAA組合誘導莖段產生愈傷組織的效果較好,推測NAA在龍珠果莖段誘導產生愈傷組織過程中占據主導地位,且NAA濃度的升高有利于龍珠果莖段形成愈傷組織。辜夕容等[16]的研究表明,NAA濃度增高有利于香樟()莖段誘導出愈傷組織,這與本試驗結果一致。由于愈傷組織數量有限,本試驗沒有進行愈傷組織增殖、分化的研究,還有待今后深入研究。
植株受到鹽脅迫時,細胞內會產生大量的活性氧,抗氧化酶系統只能清除已經產生的活性氧,無法減少活性氧的生成,屬于被動應答機制,一旦活性氧產生大于抗氧化酶系統的清除能力,植物會出現一系列不良癥狀,最明顯的表現是生長受到抑制,甚至死亡[17–18]。本研究結果表明,0~200 mmol/L NaCl對龍珠果組培苗生長沒有影響;200~800 mmol/L NaCl使植株變矮,葉片變黃萎蔫,這可能是較高濃度鹽使細胞液泡積累較多的無機離子,水勢降低, 同時根系吸收不到充分的水分和礦質元素,進而影響植株生長。
SOD能促進超氧陰離子轉化為H2O2,并傳遞H2O2給下游酶;POD能與SOD聯合作用,將SOD催化反應產生的H2O2分解清除;CAT是SOD的下游保護酶,能夠進一步催化H2O2轉化為H2O;APX也是SOD的下游保護酶,是葉綠體內解毒過氧化氫的關鍵酶,能夠以抗壞血酸為底物,催化H2O2生成H2O[19–20]。張乃群等[21]對野生大豆()幼苗的研究表明,隨NaCl濃度的升高,POD和CAT活性呈先升高后下降的變化趨勢,與本研究結果相似。
本文首次以海濱植物龍珠果為研究對象,以不同NaCl濃度模擬海濱附近生長環境,并通過4種抗氧化酶活性的變化探討龍珠果組培苗對鹽脅迫的適應程度。結果表明,SOD活性隨NaCl濃度升高逐漸升高,POD、CAT和APX活性整體呈先升高后下降的變化趨勢。0~200 mmol/L NaCl使葉片的SOD、POD、CAT和APX活性均呈現不同程度的升高,說明這4種酶協同發揮保護作用;當NaCl濃度大于400 mmol/L時,CAT和APX活性下降明顯,說明鹽濃度已超過龍珠果植株的耐鹽閾值,生長受到嚴重抑制。綜合來看,作為一種海濱植物,龍珠果對鹽脅迫具有一定的耐受能力,200 mmol/L以下的NaCl對其生長影響不大,耐鹽閾值為400 mmol/L,SOD、POD、CAT和APX等抗氧化酶共同構筑了龍珠果抵御鹽脅迫的保護系統。
本研究成功建立了龍珠果莖段叢生芽誘導和植株再生體系,探究了組培苗對鹽脅迫的適應程度,為龍珠果種苗繁育和移栽提供了理論依據和技術支持,對海濱生態環境綠化和修復具有重要意義。
[1] WANG D, LU H J, SONG H D, et al. Determination of vitexin from different medicinal parts ofLinn. [J]. Food Drug, 2014, 16(6): 425–427. doi: 10.3969/j.issn.1672-979X.2014.06.014.
王東, 盧慧娟, 宋洪東, 等. 龍珠果不同入藥部位中牡荊素的含量測定[J]. 食品與藥品, 2014, 16(6): 425–427. doi: 10.3969/j.issn.1672- 979X.2014.06.014.
[2] HUANG D. Ornamental plants and their application in Hainan seashore [J]. Flowers, 2016(3): 43–47.
黃丹. 海南海濱觀賞植物及應用 [J]. 花卉, 2016(3): 43–47.
[3] Editorial Board of Chinese Materia Medica of State Administration of Traditional Chinese Medicine. Chinese Materia Medica, Vol. 7 [M]. Shanghai: Shanghai Science and Technology Press, 1999: 478–479.
國家中醫藥管理局《中華本草》編委會. 中華本草, 第7卷 [M]. 上海: 上海科學技術出版社, 1999: 478–479.
[4] JI S G, YANG K W, HE C Y, et al. Microscopic identification of[J]. Chin Med Mat, 2012, 35(3): 391–393. doi: 10.13863/ j.issn1001-4454.2012.03.020.
姬生國, 楊克偉, 何純瑤, 等. 龍珠果的顯微鑒定 [J]. 中藥材, 2012, 35(3): 391–393. doi: 10.13863/j.issn1001-4454.2012.03.020.
[5] Delectis Florae Reipublicae Popularis Sinicae, Agendae Academiae Sinicae Edita. Florae Reipublicae Popularis Sinicae, Tomus 52(1) [M]. Beijing: Science Press, 1999: 111.
中國科學院中國植物志編輯委員會. 中國植物志, 第52卷第1分冊 [M]. 北京: 科學出版社, 1999: 111.
[6] KRISHNAVENI A, THAAKUR S R. Pharmacognostical and preli- minary phytochemical studies of[J]. Anc Sci Life, 2008, 27(3): 19–23.
[7] MURASHIGE T, SKOOG F. A revised medium for rapid growth and bio assays with tobacco tissue cultures [J]. Physiol Plant, 1962, 15(3): 473–497. doi: 10.1111/j.1399-3054.1962.tb08052.x.
[8] BEAUCHAMP C, FRIDOVICH I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels [J]. Anal Biochem, 1971, 44(1): 276–287. doi: 10.1016/0003-2697(71)90370-8.
[9] MU?OZ-MU?OZ J L, GARCíA-MOLINA F, GARCíA-RUIZ P A, et al. Enzymatic and chemical oxidation of trihydroxylated phenols [J]. Food Chem, 2009, 113(2): 435–444. doi: 10.1016/j.foodchem.2008.07. 076.
[10] QUINTANILLA-GUERRERO F, DUARTE-VáZQUEZ M A, GARCíA- ALMENDAREZ B E, et al. Polyethylene glycol improves phenol removal by immobilized turnip peroxidase [J]. Bioresour Technol, 2008, 99(18): 8605–8611. doi: 10.1016/j.biortech.2008.04.031.
[11] CHANCE B, MAEHLY A C. Assay of catalases and peroxidases [J]. Method Enzymol, 1955, 2: 764–775. doi: 10.1016/S0076-6879(55)023 00-8.
[12] NAKANO Y, ASADA K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts [J]. Plant Cell Physiol, 1981, 22(5): 867–880. doi: 10.1093/oxfordjournals.pcp.a076232.
[13] WANG Q, CHEN D Y, YANG G, et al. Tissue culture and rapid propa- gationof[J]. Plant Physiol J, 2018, 54(2): 232–236. doi: 10.13592/j.cnki.ppj.2018.0006.
王強, 陳冬怡, 楊國, 等. 鱷嘴花的組織培養和快速繁殖 [J]. 植物生理學報, 2018, 54(2): 232–236. doi: 10.13592/j.cnki.ppj.2018.0006.
[14] HOU S X, ZHAO Y X, LI J W, et al. Effects of different growth regulators on the rapid propagationof[J]. Mol Plant Breed, 2018, 16(3): 938–942. doi: 10.13271/j.mpb.016. 000938.
侯少霞, 趙永秀, 李佳雯, 等. 生長調節劑對香石竹離體快繁的影響 [J]. 分子植物育種, 2018, 16(3): 938–942. doi: 10.13271/j.mpb. 016.000938.
[15] WANG G P, XIAO R, LI C Y, et al. Study on the establishment of stem regenerating system from[J]. Chin Agric Sci Bull, 2011, 27(31): 184–188.
王國平, 肖蓉, 李春燕, 等. 棗莖段再生體系建立的研究 [J]. 中國農學通報, 2011, 27(31): 184–188.
[16] GU X R, HUANG J G, YANG Q. Initial culture of(L.) Presl[J]. Chin Agric Sci Bull, 2005, 21(2): 97– 100. doi: 10.3969/j.issn.1000-6850.2005.02.030.
辜夕容, 黃建國, 楊慶. 香樟離體培養體系的構建初探 [J]. 中國農學通報, 2005, 21(2): 97–100. doi: 10.3969/j.issn.1000-6850.2005.02. 030.
[17] QI Q, MA S R, XU W D. Advances in the effects of salt stress on plant growth and physiological mechanisms of salt tolerance [J]. Mol Plant Breed, 2020, 18(8): 2741–2746. doi: 10.13271/j.mpb.018.002741.
齊琪, 馬書榮, 徐維東. 鹽脅迫對植物生長的影響及耐鹽生理機制研究進展 [J]. 分子植物育種, 2020, 18(8): 2741–2746. doi: 10.13271/ j.mpb.018.002741.
[18] LIAO Q L. Influence of soil salinity on growth and physiological indexes ofseedlings [J]. J Trop Subtrop Bot, 2012, 20(2): 163–168. doi: 10.3969/j.issn.1005-3395.2012.02.010.
廖啟炓. 土壤鹽度對加拿利海棗幼苗生長與生理指標的影響 [J]. 熱帶亞熱帶植物學報, 2012, 20(2): 163–168. doi: 10.3969/j.issn. 1005-3395.2012.02.010.
[19] ISHIKAWA T, SAKAI K, YOSHIMURA K, et al. cDNAs encoding spinach stromal and thylakoid-bound ascorbate peroxidase, differing in the presence or absence of their 3?-coding regions [J]. Febs Lett, 1996, 384(3): 289–293. doi: 10.1016/0014-5793(96)00332-8.
[20] SI C, ZHANG J Y, XU H C. Advances in studies on growth meta- bolism and response mechanisms of medicinal plants under drought stress [J]. China J Chin Mat Med, 2014, 39(13): 2432–2437. doi: 10. 4268/cjcmm20141316.
司燦, 張君毅, 徐護朝. 藥用植物在干旱脅迫下生長代謝變化規律及應答機制的研究進展 [J]. 中國中藥雜志, 2014, 39(13): 2432– 2437. doi: 10.4268/cjcmm20141316.
[21] ZHANG N Q, LIU Y N, XIE Y H. Effects of salt stress on growth of wild soybean () seedlings [J]. Soybean Sci, 2012, 31(6): 920–923. doi: 10.3969/j.issn.1000-9841.2012.06.013.
張乃群, 劉彥娜, 解艷華. 鹽脅迫對野生大豆幼苗生長的影響 [J]. 大豆科學, 2012, 31(6): 920–923. doi: 10.3969/j.issn.1000-9841.2012. 06.013.
Stem Cultureofand Salt Tolerance of Seedlings
LIU Yan-yan1,2, SI Can2, HE Chun-mei2, YU Zhen-ming2, DUAN Jun2*
(1. Guangzhou Flowering Research Center,Guangzhou 510360, China; 2. Guangzhou Provincial key laboratory of Applied Botany, Key Laboratory of South China Agricultural Plant Molecular Analysis and Gene Improvement, South China Botanical Garden, Chinese Academy of Sciences,Guangzhou 510650, China)
In order to establish a rapid regeneration system of, the effect of plant growth regulators on adventitious bud induction, rooting were studied by using stems as explants, as well as the salt tolerance of seedlings. The results showed that the induction and growth of adventitious buds were suitable on MS+6-BA 0.5 mg/L+NAA 0.05 mg/L medium; the callus induction was appropriate on MS+6-BA 3.0 mg/L+ NAA 0.3 mg/L medium; and the rooting of plantlets was available on 1/2MS+IBA 0.2mg/L medium. The survival rate of plantlets could up to 92.6% after transplanted on matrix (peat∶vermiculite∶perlite=2∶1∶1). The growth ofseedlings was not affected under 0-200 mmol/L NaCl, while plants appeared dwarfing, leaf wilting, and yellow under more than 200 mmol/L NaCl stress. The SOD activity in leaves increased with increment of NaCl concentration, while POD, CAT and APX activities increased at first, and then decreased.These would provide technical support for the breeding ofand the restoration of seashore ecology.
; Stem; Cluster shoot; Callus; Salt stress
10.11926/jtsb.4323
2020–10–21
2021–01–25
廣東省林業科技創新項目(2015KJCX040)資助
This work was supported by the Project for Forestry Science and Technology Innovation in Guangdong (Grant No. 2015KJCX040).
劉艷艷(1992~ ), 女, 碩士。E-mail: 1970291025@qq.com
E-mail:duanj@scib.ac.cn
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14

- 熱帶亞熱帶植物學報的其它文章
- Advances in Chemical Constituents and Pharmacological Activities of Pleuropterus multiflorus
- 成都地區19個木芙蓉品種的孢粉學研究及其分類學意義
- 秋石斛花青素提取液成分分析及其體外抗氧化活性和刺激性研究
- 谷子產量和品質相關性狀的雜種優勢及遺傳特性分析
- Cloning and Expression Analysis on PmPGK1 and PmGPIC Genes in Pinus massoniana
- Michelia taishanensis Y. H. Tong, X. E. Ye, X. H. Ye & Yu Q. Chen (Magnoliaceae), A New Species from Guangdong