深圳市紅樹林典型群落的物種組成及結構多樣性研究
2021-08-20 03:01:50關開朗張信堅譚廣文曾鳳易慧琳廖文波
生態科學 2021年4期
關開朗, 張信堅, 譚廣文, 曾鳳, 易慧琳, 廖文波
深圳市紅樹林典型群落的物種組成及結構多樣性研究
關開朗1, 張信堅1, 譚廣文1, 曾鳳1, 易慧琳1, 廖文波2,*
1. 廣州普邦園林股份有限公司, 廣州 510600 2. 中山大學生命科學學院, 廣州 510275
在深圳市紅樹林主要分布區設置8個調查樣地, 以調查深圳市紅樹林典型群落的物種組成和生物多樣性特征。結果表明: (1)秋茄為深圳市天然紅樹林的建群種或優勢物種; 白骨壤及海漆是天然紅樹林的重要組成物種; 海桑及無瓣海桑是人工林的主要組成物種; 桐花樹及老鼠簕是林下灌木層以及低發展程度的紅樹林群落的主要組成物種。(2)海桑、無瓣海桑、海漆及白骨壤等真紅樹植物豐富了以秋茄為建群種、優勢種或伴生種的深圳市紅樹林的物種種類、垂直結構和徑級結構, 從而提高了紅樹林的物種多樣性和結構多樣性, 使林分更加穩定性。在對紅樹林進行質量提升或營建紅樹林人工林時, 可以通過增加本土紅樹植物的種類和提高林分混交程度和結構多樣性, 來提高紅樹林的生態穩定性和景觀觀賞價值。
紅樹林; 物種組成; 物種多樣性; 結構多樣性; 深圳
0 前言
紅樹林是分布于熱帶和亞熱帶地區潮間帶的以紅樹植物為主體的常綠群落, 具有消浪護岸、凈化環境污染、維護生物多樣性等作用[1]。我國的紅樹林主要分布在東南沿海灘涂, 包括海南、廣東、廣西、福建、臺灣、香港和澳門[2]。在廣東, 南至雷州半島, 北達饒平和南澳島, 沿海灘涂均有紅樹林間斷分布[3]。
紅樹林是深圳市植被的重要組成部分[3]。歷史上, 紅樹林曾經廣泛分布于深圳海岸潮間帶, 如保安沙井至西鄉一帶沿海灘涂的紅樹林分布面積曾達到113.4 hm2(1988年), 深圳灣紅樹林分布面積達79.56 hm2(1979年), 壩光沿海灘涂有大面積的紅樹林約200 hm2[4–5]。自改革開放以來, 隨著深圳市城市建成區擴張、填海造陸、水庫建設、病蟲害等原因, 紅樹林生態系統受到極大破壞, 分布面積減少至原來的1/4[6]。目前, 深圳市的紅樹林總面積約169.7 hm2, 主要分布在寶安區的、南山區、福田區和大鵬新區的海岸、港灣、河口灣等周期性受淹水域中。其中, 福田區紅樹林包括天然林和人工林, 寶安區和南山區的紅樹林以人工林為主, 大鵬新區的紅樹林以天然林為主[7]。為更好地對深圳市紅僅存的樹林進行保護, 已有不少學者和單位通過人工造林、防治病蟲害、消除有機污染物等措施, 對退化紅樹林進行生態修復[8–11], 深圳市的紅樹林分布面積得到一定程度的增加, 林分質量得到一定程度的提升。本研究在對深圳市紅樹林全面考察的基礎上, 對紅樹林典型群落的物種組成、群落結構等做深入調查, 并對其物種多樣性和結構多樣性進行分析和比較, 了解物種組成及群落結構對群落生物多樣性的影響, 為深圳市對紅樹林進行質量提升和營造紅樹林人工林等提供理論支撐。
1 材料與方法
1.1 樣地設置與調查方法
在深圳市主要紅樹林分布區進行路線考察, 記錄優勢紅樹植物群落的組成、分布以及紅樹林內植物的種類。在福田紅樹林保護區、大鵬壩光、大鵬東涌河口3個天然紅樹林分布區, 根據優勢紅樹植物群落的類型和分布情況進行選點并設置8個調查樣方(表1)。樣方面積和調查方法參考《濕地生態系統定位觀測技術規范》[12], 每個調查樣方大小為20 m×20 m, 分為4個10 m×10 m。植物調查指標包括: 種名、胸徑(喬木)、樹高、冠幅、蓋度(灌木、草本、藤本)、數量。
1.2 數據處理
1.2.1 物種優勢度
物種的優勢度以重要值表示, 計算方法:

1.2.2 群落結構
采用韋霄等[13]的方法, 按5 m一個高度劃分片層, 對高度區間內的植株數量及所有物種的優勢度進行統計, 分析其垂直結構特征; 按5 cm一個等級劃分胸徑級別, 比較不同徑級的植株數量和優勢度, 分析群落徑級結構特征。

表1 深圳市紅樹林典型群落調查樣地情況
1.2.3 群落多樣性的測度方法
(1)林分物種多樣性
采用Shannon-Wiener多樣性指數、Simpson優勢度指數以及基于Shannon-Wiener指數的均勻度指數來評價群落的物種多樣性[14]。
Shannon-Wiener多樣性指數
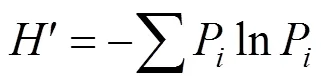
Simpson優勢度指數

均勻度指數

(2)林分結構多樣性
采用LEI等[15]方法, 通過測定群落物種直徑大小多樣性和物種多樣性的二維聯合熵以及物種直徑大小多樣性和樹高多樣性的二維聯合熵, 來表示林分結構多樣性。
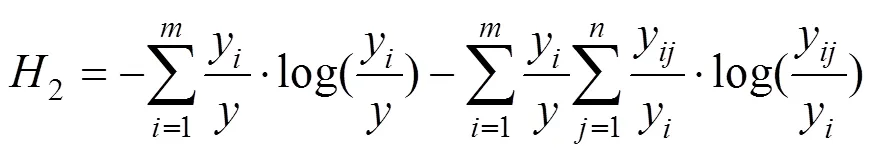
2 結果與分析
2.1 深圳市紅樹林典型群落的物種組成及優勢度
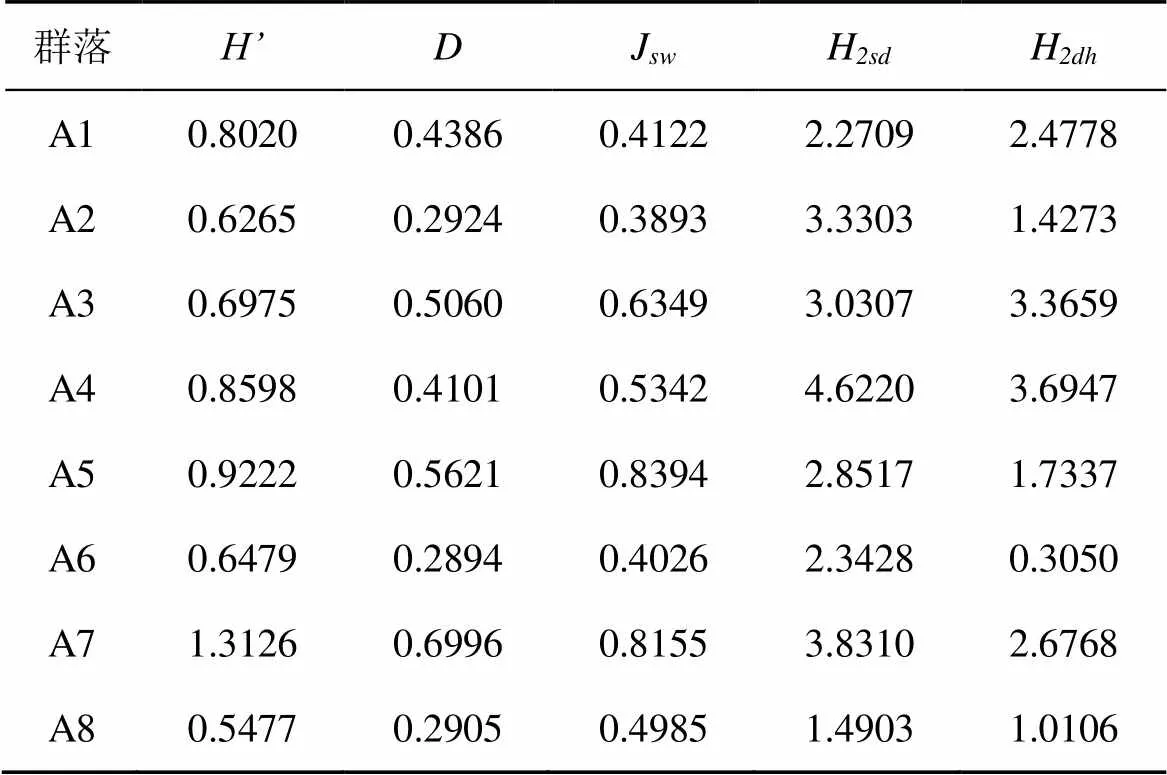
福田紅樹林典型紅樹群落喬木層組成物種主要有秋茄、白骨壤、無瓣海桑、海桑及木欖, 形成分別以秋茄、白骨壤、無瓣海桑、海桑中的一種或幾種為優勢種的紅樹群落。從重要值來看, 秋茄的重要值區間范圍為21.5—100, 表明秋茄主要作為建群種、優勢種或伴生種, 廣泛分布于福田紅樹林各群落內, 其種群年齡結構主要以平均高度從3.8 cm至7.7 cm不等的壯樹(2.5 cm 大鵬東涌的紅樹林為自然林, 雖然由于靠近東涌村而受到長期的人為干擾, 但總體上還是保留了比較完整的紅樹林群落。根據路線調查結果, 東涌紅樹林潮間內帶真紅樹植物種類主要有海漆、秋茄、桐花樹、老鼠簕、木欖及白骨壤。真紅樹植物群落共有兩類, 分別是以海漆為特征種的混合群落及以秋茄為特征種的混合群落, 還有沿著河涌上游小面積分布的桐花樹、老鼠簕群落。如表2所示, 東涌紅樹林真紅樹群落喬木層以秋茄和海漆為建群種或優勢種, 重要值區間范圍為11.6—66.1。其中, 秋茄主要以壯樹為主, 多分枝, 平均胸徑僅2.8—6.6 cm, 平均高度僅1.8—6.3 m, 表明秋茄種群總體年齡較小; 海漆主要以壯樹為主, 平均胸徑5.8—8.0 cm, 平均高度2.3—7.3 cm, 種群總體年齡適中; 木欖在東涌紅樹林零星分布, 局部形成優勢種, 其種群也主要以壯樹為主。灌木層主要有老鼠簕和桐花樹, 兩者優勢度相近, 但在河中心位置, 桐花樹優勢明顯, 可能與潮位低有關。 表2 深圳市紅樹林典型群落的物種組成及優勢度 大鵬壩光紅樹林是大鵬半島現存面積最大的紅樹林, 真紅樹植物主要有白骨壤、秋茄、桐花樹、老鼠簕及海漆。真紅樹群落類型主要以白骨壤群落為主, 其次為小面積的與白骨壤群落混交的秋茄人工群落。典型的白骨壤群落分布于鹽灶村入口灘涂。相對于深圳其他區域的紅樹林而言, 該白骨壤群落物種組成較為單調, 僅包括白骨壤、秋茄、桐花樹3種。其中, 白骨壤在喬木層占優勢, 重要值達78.4; 秋茄次之, 重要值為21.6。從胸徑及高度來看, 白骨壤及秋茄的平均胸徑為2.2—6.9 cm, 平均高度為2.3—2.8 m, 表明白骨壤和秋茄的種群年齡均偏小。灌木層稀疏, 僅見桐花樹, 數量十分稀少。 2.2.1 群落垂直結構特征 群落的垂直結構主要指群落的分層現象, 群落中各種種群的個體在空間不同層次上分布的象, 也叫成層性[16]。一般而言, 生境條件越優或者發育階段越老的群落, 層次結構越多, 群落也就越高[13]。 對深圳市紅樹林8個典型群落喬木層的垂直結構進行分析, 結果見圖1。在福田紅樹林, 秋茄–桐花樹+老鼠簕群落(A1)分為兩個片層, 10—15 m片層僅見少量秋茄, 5—10 m片層全部由秋茄組成, 5 m以下片層主要由桐花樹及少量的老鼠簕組成, 偶見無瓣海桑、白骨壤及短葉茳芏, 層間植物還有少量魚藤(圖1a)。秋茄+白骨壤–老鼠簕+桐花樹群落(A2)喬木層明顯可分為兩個片層, 5—10 m片層主要由秋茄、白骨壤的成年植株組成, 且植株數量與優勢度較高; 5 m以下片層主要由少量的秋茄、白骨壤的小樹以及林下的密集的桐花樹、老鼠簕組成。無瓣海桑–老鼠簕群落(A3)和海桑–老鼠簕群落(A4)的喬木層群落整體高度一般高于10 m, 且明顯分為四個片層, 各片層的植株數量與優勢度比較接近。15 m以上片層全部由海桑或無瓣海桑組成, 10—15 m片層主要由海桑或無瓣海桑組成, 還包括少量的秋茄; 5—10 m片層主要由秋茄以及部分海桑和無瓣海桑小樹組成; 5 m以下片層主要由秋茄小樹、海桑和無瓣海桑幼苗以及老鼠簕組成, 整體優勢度略高于其他片層(圖1c–d)。(秋茄)–桐花樹+老鼠簕群落(A5)明顯分為兩個片層, 5—10 m主要由秋茄和少量桐花樹組成, 5 m以下片層主要由大部分桐花樹以及老鼠簕組成(圖1e)。 與福田紅樹林相比, 大鵬東涌紅樹林、壩光紅樹林的群落喬木層垂直結構比較簡單。群落整體高度低于10 m, 個別海漆植株高度可超過15 m。其中, 東涌紅樹林的秋茄–桐花樹群落(A6)高度低于5 m, 群落喬木層分層不明顯, 秋茄、海漆、木欖組成了低矮的喬木層, 少量的桐花樹、老鼠簕組成了稀疏的林下灌草層(圖1f)。海漆–老鼠簕+桐花樹群落(A7)的結構稍復雜, 大致可分為四個片層。15 m以上片層僅見海漆, 植株數量及優勢度低; 10—15m片層由少量海漆及秋茄組成, 優勢度較低; 5—10 m片層主要由海漆及少量秋茄組成, 植株數量及優勢度遠大于其他片層; 5 m以下片層主要由少量的海漆及秋茄小樹組成, 優勢度相對較低(圖1g)。壩光紅樹林的白骨壤–桐花樹群落整體高度低于10 m, 群落垂直結構比較簡單。優勢片層為5 m以下片層, 主要由白骨壤及少量秋茄小樹組成; 5—10 m片層包括白骨壤及秋茄, 植株數量少, 優勢度低(圖1h)。 注: a. 秋茄–桐花樹+老鼠簕群落; b. 秋茄+白骨壤–老鼠簕+桐花樹群落; c. 無瓣海桑–老鼠簕群落; d. 海桑–老鼠簕群落; e. (秋茄)–桐花樹+老鼠簕群落; f. 秋茄–桐花樹群落; g. 海漆–老鼠簕+桐花樹群落; h. 白骨壤–桐花樹群落。 Figure 1 Vertical structures of arbor layers of typical mangrove communities in Shenzhen 2.2.2 群落徑級結構特征 按許格希等[17]方法, 對深圳市典型紅樹林群落的優勢物種作徑級結構分析。結果顯示, 福田紅樹林秋茄–桐花樹+老鼠簕群落(A1)、秋茄+白骨壤–老鼠簕+桐花樹群落(A2)及(秋茄)–桐花樹+老鼠簕群落(A5)主要以小徑級(d<5 cm, 下同)及中徑級(5 cm≤ d<15 cm, 下同)植株為主, 表明上述群落正分別處于幼齡林(圖2a)以及中林齡階段(圖2b, 2e)。無瓣海桑–老鼠簕群落(A3)和海桑–老鼠簕群落(A4)各徑級植株數量與優勢度比較一致, 表明群落正處于由中齡林向成熟林過渡階段, 同時, 林下幼苗或小樹的數量和優勢度也較大, 顯示出群落良好的更新狀況(圖2c–d)。(秋茄)–桐花樹+老鼠簕群落(A5)的植株徑級主要集中以中徑級為主, 少量植株胸徑小于5 cm或者大于15 cm(圖2e)。大鵬東涌紅樹林的秋茄–桐花樹群落(A6)主要以小徑級為主, 植株數量及優勢度占絕對優勢, 中徑級和大徑級(d≥15 cm, 下同)的植株數量較少, 優勢度較低, 顯示出群落正處于幼齡林階段, 群落發展空間較大(圖2f); 海漆–老鼠簕+桐花樹群落(A7)主要以及中徑級為主, 小徑級植株數量少于中徑級, 遠多于大徑級, 但優勢度僅略大于大徑級, 表明該群落正處于由中齡林向成熟林過渡的階段, 有一定的林分更新能力(圖2g)。與東涌紅樹林群落類似, 大鵬壩光的白骨壤–桐花樹群落同樣以小徑級和中徑級植株為主, 大徑級植株數量較少, 同樣處于由中齡林向成熟林過渡的階段(圖2h)。 2.3.1 群落物種多樣性 Shannon-Wiener 物種多樣性指數、Simpson物種多樣性指數及基于Shannon-Wiener物種多樣性指數的均勻度指數是反映群落中物種種類與個體數量、優勢種的組成及其均勻性的綜合指標[14]。對深圳市8個典型紅樹林群落進行物種多樣性及均勻度進行分析, 結果如表3所示。與其他群落相比, 大鵬東涌的海漆–老鼠簕+桐花樹群落(A7)的物種多樣性指數、生態優勢度指數及均勻度指數均最高, 表明該群落具有最高物種多樣性, 群落中優勢物種最多, 且群落物種的分布最均勻。福田紅樹林的(秋茄)–桐花樹+老鼠簕群落(A5)的生物多樣性指數、生態優勢度指數及均勻度指數均高于福田紅樹林其他典型群落, 但低于大鵬東涌的海漆–老鼠簕+桐花樹群落(A7)。大鵬壩光的白骨壤–桐花樹群落的物種多樣性指數及優勢度指數均最低(A8), 均勻度指數略高于福田紅樹林的秋茄–桐花樹+老鼠簕群落(A1)、秋茄+白骨壤–老鼠簕+桐花樹群落(A2)及大鵬東涌的秋茄–桐花樹群落(A6), 表明該群落物種多樣性最低, 群落優勢物種單一, 物種分布比較集中。整體而言, 福田紅樹林整體上顯示出較高的物種多樣性, 優勢種比較突出, 群落物種分布比較均勻, 這可能與福田紅樹林處于長期的保護, 恢復程度較好有關。位于東涌河中心高地的海漆–老鼠簕+桐花樹群落(A7)與位于河岸的秋茄–桐花樹群落(A6)在物種多樣性、優勢度及均勻度方面呈現兩極分化的情況, 可能是由于群落位置的不同導致受干擾的程度不同有關。壩光紅樹林由于長期受到人為干擾, 導致物種組成簡單, 群落發展程度低, 所以物種多樣性、群落優勢度與均勻度均較低。 注: a.秋茄–桐花樹+老鼠簕群落; b.秋茄+白骨壤–老鼠簕+桐花樹群落; c.無瓣海桑–老鼠簕群落; d.海桑–老鼠簕群落; e.(秋茄)–桐花樹+老鼠簕群落; f.秋茄–桐花樹群落; g.海漆–老鼠簕+桐花樹群落; h.白骨壤–桐花樹群落。 Figure 2 Diameter structures of arbor layers of typical mangrove communities in Shenzhen 2.3.2 群落結構多樣性 Shannon-Wiener多樣性指數、Simpson多樣性指數及的均勻度指數均從物種多樣性角度對群落生物多樣性進行定量描述, 不能反映出群落結構對群落生物多樣性的影響。而研究表明, 提高林分結構多樣性和復雜性, 可以維持和增加林分的生物多樣性[18]。LEI等[15]認為, 基于聯合熵的多維多樣性指數可以代表林分結構的多個組成部分, 是描述林分總體結構多樣性的好方法。本研究以5 cm為一個胸徑等級, 以5 m為一個高度等級, 對深圳市紅樹林典型群落的結構多樣性進行分析。結果顯示, 各群落的結構多樣性大小順序與物種多樣性大小順序差異較大, 主要體現在具有較豐富的物種組成、徑級結構和層次結構的群落, 結構多樣性一般較大, 如福田紅樹林的海桑–老鼠簕群落(A4)、無瓣海桑–老鼠簕群落(A3)及東涌的海漆–老鼠簕+桐花樹群落(A7)。福田紅樹林的秋茄+白骨壤–老鼠簕+桐花樹群落(A2)具有較豐富的物種和徑級結構, 但垂直結構簡單(表2, 圖1b), 故樹種和林木大小的總體多樣性較高而林木大小和樹高的總體多樣性較低(表3)。(秋茄)–桐花樹+老鼠簕群落(A5)的物種多樣性、優勢度及均勻度較高, 但林木大小和樹高的總體多樣性較低(表3), 原因是該群落具有較大的植株密度和較多的優勢種, 但群落物種較少且垂直結構簡單(表2, 圖1e)。與其他群落相比, 秋茄–桐花樹+老鼠簕群落(A1)因物種組成、優勢種數量與分布、垂直結構和徑級結構比較均勻, 因而結構多樣性指數處于比較處于居中位置。東涌的秋茄–桐花樹群落(A6)與壩光的白骨壤–桐花樹群落(A8)的結構多樣性較低, 與這兩個群落的物種組成、徑級和層次結構均較少有很大關系。 表3 深圳市紅樹林典型群落多樣性指數 深圳市紅樹林主要以本土少數幾種真紅樹植物以及引種真紅樹植物形成的單優或者混合群落為主, 在植被類型與種類組成上與淇澳島紅樹林保護區[19]、惠州紅樹林保護區[20]等珠三角地區的紅樹林類似。本研究調查的8個群落反映出了秋茄、白骨壤、桐花樹、老鼠簕、海漆、海桑、無瓣海桑及木欖這幾種真紅樹植物在深圳市紅樹林的物種組成和群落結構上不同的地位。由表2可以看到, 秋茄是上述調查的全部紅樹植物群落的建群種、優勢種或伴生種, 一定程度上說明了秋茄是深圳市紅樹林的建群種或優勢種。白骨壤在福田紅樹林中潮位區域形成局部優勢群落, 以及是大鵬壩光紅樹林的建群種, 在深圳市天然紅樹林物種組成及群落結構的重要性上僅次于秋茄。海漆在東涌河口成規模分布并且是優勢種, 在福田紅樹林僅小面積分布, 在深圳其他區域零散分布, 因此是深圳市紅樹林的重要組成物種之一。海桑及無瓣海桑是深圳市紅樹林人工林的主要組成物種, 主要集中分布在福田紅樹林及寶安西灣紅樹林公園內, 在群落內的優勢度非常明顯, 甚至形成單優群落, 在一些入海河口也有零星分布。桐花樹及老鼠簕是紅樹林林下灌草層主要組成物種, 兩者在低、中、高潮位均有分布, 無明顯分布規律。在一些喬木層發達, 郁閉度較高的林分內, 桐花樹、老鼠簕的多度均較低, 可能與這兩個物種的生長需要一定強度的光照有關[21–23]。 以物種多樣性指數、優勢度指數及均勻度指數為代表的一維熵在一定程度上反映出了群落物種組成、植株密度及混交程度的差異。群落中優勢物種的種類越多, 且在群落中的分布越均勻, 生物多樣性越高, 代表性群落為大鵬東涌的海漆–老鼠簕+桐花樹群落。群落中的優勢物種越少, 且在群落中的分布越集中, 生物多樣性越低, 代表性群落為大鵬壩光的白骨壤–桐花樹群落。也就是說, 群落中更多的優勢物種組成以及更高的混交程度, 意味著更高的物種多樣性(表2、表3)。但僅從物種多樣性的角度來評價群落生物多樣性顯然是不夠充分的。同樣, 僅從群落的徑級結構多樣性或層次結構多樣性來評價群落生物多樣性同樣也是有缺陷的[15]。物種與胸徑、胸徑與高度的二維聯合熵可以綜合反映出群落的物種組成、垂直結構和徑級結構的復雜程度, 是一個能夠很好衡量群落結構多樣性的指標。從本研究分析結果來看, 以海桑、無瓣海桑為優勢種的人工林, 由于高大喬木海桑和無瓣海桑的存在, 使得林分具有較豐富的徑級結構和垂直結構, 林分的結構多樣性較高, 群落也就比較穩定。與此類似, 東涌紅樹林的海漆–老鼠簕+桐花樹群落也因海漆的存在, 群落的結構多樣性也均較高。 從以上結果來看, 通過增加紅樹林物種種類, 來提高以秋茄為建群種或優勢種的紅樹林的物種多樣性和結構多樣性, 從而提高林分的穩定性和健康度, 作用是十分明顯的。從群落演替過程上來看, 紅樹林在發展到一定程度時, 會發生自疏效應, 主要表現為植株分枝干枯頻繁發生為特征, 其原因主要是種內個體對資源的競爭[24–25]。因此, 除了要控制好種群密度外, 還要增加處于不同生態位的物種, 以充分利用空間資源, 減少生態位重疊。另外, 研究表明, 紅樹林的害蟲種類與樹種有關, 不少害蟲有其主要的寄主[10]。因此, 通過增加紅樹林物種種類群落混交度, 增加群落的物種多樣性, 以提高群落的穩定性, 從而增強對病蟲害的抵抗能力, 預防針對單一樹種的大規模病蟲害。海桑和無瓣海桑對提高紅樹林的結構多樣性的作用是明顯的, 但也會擠壓其他種群的生長空間, 對林分的物種多樣性可能會存在消極影響。目前, 海桑及無瓣海桑給紅樹林帶來的生態影響尚存在爭議。ZAN等[26]從種群生物生態學特性、生態位理論、群落更新理論等方面分析后, 認為海桑和無瓣海桑在深圳灣造成生態入侵的可能性不大。廖寶文等[27]對海南東寨港紅樹林的無瓣海桑群落進行研究后, 認為無瓣海桑目前不會造成生態入侵, 但它對今后的生態影響仍需加強監測。有研究表明, 無瓣海桑能夠對與其混交的桐花樹、白骨壤種群的發展產生不利影響[28], 但能提高秋茄的種群密度[28]。與海桑和無瓣海桑相比, 本土物種海漆、白骨壤與木欖既能增加林分的物種多樣性和結構多樣性, 又不會造成生態入侵, 是改造以秋茄為建群種或優勢種的深圳紅市紅樹林或營造紅樹林人工林的優良樹種。東涌的海漆林是深圳地區乃至華南地區難得一見的天然海漆林。每年夏天, 海漆葉子在會逐漸變黃最終變紅, 東涌紅樹林便形成華南地區難得一見的彩葉紅樹林景觀, 為華南地區紅樹林林分改造、質量提升或營建兼具生態價值和景觀價值的紅樹林人工林提供了優良的模板。 [1] 段舜山, 徐景亮. 紅樹林濕地在海岸生態系統維護中的功能[J]. 生態科學, 2004, 23(4): 351–355. [2] 王文卿, 王瑁. 中國紅樹林[M]. 北京:科學出版社, 2007. [3] 繆紳裕, 廖文波, 蘇志堯. 廣東紅樹林研究概況及廣東紅樹林的主要特征[C]∥黃玉山, 譚鳳儀. 廣東紅樹林研究論文選集. 廣州: 華南理工大學出版社, 1997: 73–84. [4] 陳保瑜, 宋悅, 昝啟杰, 等. 深圳灣近30年主要景觀類型之演變[J]. 中山大學學報(自然科學版), 2012, 51(5): 86–92. [5] 張倬綸, 侯霄霖, 梁文釗, 等. 深圳現存紅樹林群落的生境及保護對策[J]. 濕地科學與管理, 2012(4): 49–52. [6] 王志明, 劉晶. 一座城市容不下一片紅樹林?[N]. 中國環境報, 2012–11–30(008). [7] 姜劉志, 楊道運, 梅立永, 等. 深圳市紅樹植物群落碳儲量的遙感估算研究[J]. 濕地科學, 2018, 16(5): 618–625. [8] 陳桂珠, 彭紹堅, 王震宇, 等. 深圳市西部地區灘涂濕地生態的改造與恢復示范研究[C]//周光召. 加入WTO和中國科技與可持續發展–挑戰與機遇、責任和對策(上冊). 北京: 中國科學技術出版社, 2002: 497. [9] 沈凌云, 寧天竹, 吳小明, 等. 深圳灣鳳塘河口紅樹林修復工程[J]. 價值工程, 2010(14): 55–57. [10] 昝啟杰, 譚鳳儀, 李喻春. 濱海濕地生態系統修復技術研究–以深圳灣為例[M]. 北京: 海洋出版社, 2013. [11] 沈小雪, 姜立得, 陶佳偉, 等. 退化紅樹林的生態修復實驗研究[J]. 北京大學學報(自然科學版), 2019 (4), doi: 10.13209/j.0479–8023.2019.046. [12] LY/T 2898—2017, 濕地生態系統定位觀測技術規范[S]. 北京: 中國標準出版社, 2008. [13] 韋霄, 柴勝本, 陳宗游, 等. 珍稀瀕危植物金花茶保育生物學研究[M]. 南寧: 廣西科學技術出版社, 2015. [14] 王伯蓀, 余世孝, 彭少麟, 等. 植物群落學試驗手冊[M]. 廣州: 廣東高等教育出版社, 1996. [15] LEI X D, LU Y C, Information entropy measures for stand structural diversity: joint entropy[J]. Forestry Studies in China, 2004, 7(2): 12–15. [16] 姜在民, 賀學禮. 植物學[M]. 咸陽: 西北農林科技大學出版社, 2016. [17] 許格希, 史作民, 唐敬超, 等. 物種多度和徑級尺度對于評價群落系統發育結構的影響:以尖峰嶺熱帶山地雨林為例. 生物多樣性[J], 2016, 24(6): 617–628. [18] 鄭景明, 羅菊春. 長白山闊葉紅松林結構多樣性的初步研究[J]. 生物多樣性, 2003(11): 295–302. [19] 周凡, 鄺棟明, 簡永強, 等. 珠海市淇澳島紅樹林群落組成初步研究[J]. 生態科學, 2003, 22(3) : 237–241. [20] 姚少慧, 孫妮, 苗莉, 等. 惠州紅樹林保護區紅樹植物群落結構特征[J]. 廣東農業科學, 2013(17): 153–157. [21] 刁俊明, 曾憲錄. 光強對桐花樹根系生長和根系活力的影響. 光強對桐花樹根系生長和根系活力的影響[J], 嘉應學院學報, 2013(8): 68–75. [22] 刁俊明, 鐘福生, 劉惠娜. 遮光處理對桐花樹光合特性的影響[J]. 嘉應學院學報, 2011, 29(5): 71–77. [23] 劉濱爾, 廖寶文. 老鼠簕幼苗在潮汐環境下對不同光強的生理生態響應[J]. 林業科學研究, 2013(2): 102–199. [24] LIAO W B, LAN C Y, ZAN Q J, et al. Growth dynamics and self-thinning of the dominant populations in the mangrove community[J]. Acta Botanica Sinica, 2004, 46(5): 522–532. [25] 孫龍, 國慶喜, 孫慧珍. 生態學基礎[M]. 北京: 中國建工業出版社, 2013. [26] ZAN Q J, WANG B S, WANG Y J, et al. Ecological assessment on the introducedandat the mangrove forest of Shenzhen Bay, China[J]. Acta Botanica Sinica, 2003, 45(5): 544–551. [27] 廖寶文, 李玫, 鄭松發, 等. 外來種無瓣海桑種內、種間競爭關系研究[J]. 林業科學研究, 2003, 16(4): 418–422. [28] 李玫, 廖寶文, 鄭松發, 等. 無瓣海桑的直接引入對次生桐花樹群落的擾動[J]. 林業科學與環境, 2004, 20(3): 19–21. [29] 姚歷強. 無瓣海桑引入對龍海九龍江口鄉土紅樹林影響的調查[J]. 林業勘察設計, 2010(2): 76–78. Species composition and structural diversity of typical mangrove communities in Shenzhen GUAN Kailang1, ZHANG Xinjian1, TAN Guangwen1, ZENG Feng1, YI Huilin1, LIAO Wenbo2,* 1. Pubang Landscape Architecture Co., LTD, Guangzhou 510600, China 2. School of Life Science, Sun Yat-sen University, Guangzhou 510275, China To investigate the species composition and structural diversity of typical mangrove communities in Shenzhen, eight plots were set up in its main mangrove distribution areas. The results are as follows. (1)was the constructive or dominant natural mangrove species;andwere important constituent species in the natural mangroves;andwere the main constituent species for man-made mangroves;andwere the main species constituting the shrub layer and the less developed mangrove areas. (2) True mangrove plants, such as,,and, etc., enriched the species composition, vertical structure, and diameter structure of Shenzhen mangroves; whileacted as the constructive, dominant or associated species in the region, thus improved the species diversity and structural diversity within the mangroves in becoming a stable stand. The development in natural mangrove quality or cultivated mangroves could improve the ecological stability and landscape value of mangroves by increasing the number of native mangrove plant species, stand structural diversity and mixture level. mangroves; species composition; species diversity; structural diversity; Shenzhen 關開朗, 張信堅, 譚廣文, 等. 深圳市紅樹林典型群落的物種組成及結構多樣性研究[J]. 生態科學, 2021, 40(4): 83–91. GUAN Kailang, ZHANG Xinjian, TAN Guangwen, et al. Species composition and structural diversity of typical mangrove communities in Shenzhen[J]. Ecological Science, 2021, 40(4): 83–91. 10.14108/j.cnki.1008-8873.2021.04.010 Q948.15 A 1008-8873(2021)04-083-09 2020-01-19; 2020-03-02 深圳市生態環境局科研項目(SZDL2019333099) 關開朗(1988—), 男, 碩士, 主要從事生物資源調查與評估工作, E-mail: guankailang@foxmail.com 廖文波, 男, 博士, 教授, 主要從事植物區系研究, E-mail: lsslwb@mail.sysu.edu.cn
2.2 群落結構特征

2.3 群落多樣性

3 討論
猜你喜歡
歌海(2024年6期)2024-03-18 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
兒童故事畫報·自然探秘(2022年3期)2022-04-27 00:17:10
趣味(語文)(2020年3期)2020-07-27 01:42:46
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
瘋狂英語·初中天地(2018年6期)2018-11-24 02:39:24
汽車觀察(2018年10期)2018-11-06 07:05:26
歌海(2018年4期)2018-05-14 12:46:15
作文與考試·初中版(2017年12期)2017-04-19 20:26:27
少兒科學周刊·少年版(2015年1期)2015-07-07 17:15:12
