枯萎期蘆葦收割時間對濕地脫氮效果及根系呼吸代謝的影響
2021-08-23 10:23:52王俊力付子軾喬紅霞陳桂發劉福興
環境科學研究 2021年8期
王俊力, 付子軾, 喬紅霞, 陳桂發, 劉福興*
1.上海市農業科學院, 上海 201403 2.上海低碳農業工程技術研究中心, 上海 201415
氮(N)是生態系統中構成生物體的重要元素,但也是引起水體富營養化的主要元素之一[1]. 自工業革命以來,全球范圍內人為源氮排放從1860年的15×1012g/a增至20世紀90年代初的156×1012g/a,預計到2050年將達到267×1012g/a[2]. 我國農業生產使用的氮肥中有25%的氮隨降雨徑流和滲漏進入水環境,是導致水體氮濃度升高和污染加重的重要原因[3]. 濕地作為陸地與水體之間的過渡帶,對攔截陸地方向的氮污染源、降低水體中的氮水平具有重要作用[4].
濕地通過復雜的物理、化學和生物等方面的作用實現氮去除,方式包括揮發、氨化、硝化/反硝化、植物吸收、基質吸附等[5],其中生物(植物和微生物)起到關鍵作用[6]. 研究[7-8]表明,超過50%的氮通過微生物硝化/反硝化作用去除,而植物會影響濕地微生物群落結構的時空變化. 蘆葦(Phragmitesaustralis)是濕地生態系統中的主要植物之一,常見于河口、湖濱、海岸等地,在全球廣泛分布[9]. 在我國的亞熱帶季風氣候區,分布有大面積蘆葦濕地,該地區積溫高,無霜期長,蘆葦通常在3月份出芽,到11月份進入枯萎期,冬季根系休眠時間較短[10]. 研究表明,生長期蘆葦濕地的脫氮效率為50%~90%[11-12],冬季則降至20%左右[13]. 雖然冬季蘆葦地上部分枯萎,但其根系仍存在活力[14],了解冬季蘆葦根系活動,對如何采取有效措施促進冬季濕地脫氮效果具有重要意義.
收割是濕地植物最主要的管理措施之一. 蘆葦一年收割一次能夠去除地上部枯死量,有效恢復其活力[15]. 在冬季開始時,濕地植物地上部分的氮逐漸向地下轉移,如果不收割,植物吸收的氮可能會隨著植株的枯萎腐爛重新回到濕地系統中,出現水體氮濃度升高現象[16]. 大型蘆葦濕地系統中,收割措施具有局限性[17],因為其需要更多的人力、物力和財力投入. 因此,選擇最佳收割時間,是影響冬季蘆葦濕地脫氮效果的重要因素.
呼吸代謝的持續進行是植物休眠過程的顯著特點[18],在亞熱帶地區,冬季蘆葦進入休眠,但其根系仍存在活力,冬季蘆葦根系呼吸代謝的變化可能會導致濕地脫氮過程發生變化. 枯萎期不同時間收割是否會影響根系的呼吸代謝特征,從而影響濕地脫氮效果,有關這方面的研究還鮮見報道. 該研究系統探討了枯萎期不同收割時間情況下,蘆葦根系活力和呼吸代謝途徑關鍵酶活性,及其與濕地脫氮效果的關系,以期探討枯萎期收割影響植物根系呼吸的作用機制,為人工濕地的管理和功能發揮、富營養化水體的治理提供理論依據.
1 材料與方法
1.1 研究區概況
試驗研究位于上海市農業科學院莊行綜合試驗站(30°53′N、121°23′E). 該區屬亞熱帶季風氣候,多年平均降水量 1 191.5 mm,蒸發量 1 236.8 mm,年均溫度16.1 ℃,全年日照時數 1 900.2 h,無霜期224.4 d.
1.2 試驗設計
試驗采取模擬蘆葦濕地的方式進行,構建12個相同結構的傳統水平潛流濕地系統(見圖1),長×寬×深為150 cm×50 cm×80 cm,進水端及出水端各設置長度為20 cm的布水區和集水區,并設有出水管,主體填料區域長度為110 cm. 進出水均采用穿孔板,出水孔與出水管底標高一致. 為了便于測定濕地內部的水質參數,在系統中間沿程豎向設置內徑5 cm穿孔管. 內部填充粒徑約1~2 cm的礫石基質,基質層深70 cm,運行時水在介質表面以下流動,水深為60 cm.
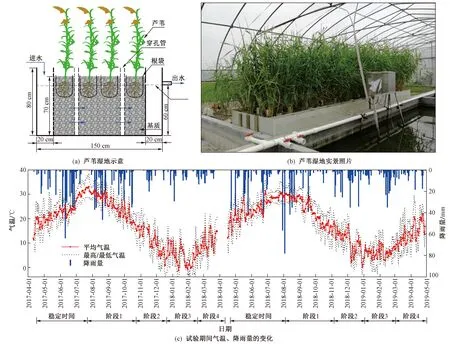
注: 試驗第1年(2017年7月—2018年3月)蘆葦于2017-04-10移栽,濕地穩定至2017-07-14正式開始試驗,階段1為正式試驗至2017-10-13枯萎前收割,階段2為枯萎前收割后至2017-12-12枯萎中期收割,階段3為枯萎中期收割后至2018-02-08枯萎末期收割,階段4為枯萎末期收割后至2018-03-27結束試驗;試驗第2年(2018年8月—2019年4月)蘆葦于2018-04-16移栽,濕地穩定至2018-08-01正式開始試驗,階段1為正式試驗至2018-10-31枯萎前收割,階段2為枯萎前收割后至2018-12-28枯萎中期收割,階段3為枯萎中期收割后至2019-02-28枯萎末期收割,階段4為枯萎末期收割后至2019-04-28結束試驗. 下同.圖1 蘆葦濕地示意、實景照片以及試驗期間氣溫、降雨量的變化Fig.1 Schematic diagram of a single wetland system, picture of the experimental site, and fluctuation of air temperature and daily precipitation during the experiment
植物采用當地常見蘆葦(Phragmitesaustralis),選取長勢相近的蘆葦幼苗移栽至根袋(300目尼龍網袋,20 cm×30 cm)中,袋內填充基質與袋外相同[19],系統內的初始種植密度為2株/袋(約29株/m2),蘆葦在系統中培養約2個月,待長勢平穩后開始進水,2年試驗時間軸如圖1所示(第1年為2017年7月—2018年3月,第2年為2018年8月—2019年4月). 進水利用河道水配置,用尿素調節進水ρ(TN),利用恒流泵進行連續進水,水力負荷(HLR)為100 mm/d,水力停留時間(HRT)為2.4 d. 系統運行約1個月趨于穩定后,正式開始試驗.
試驗設置4個處理,即W1(對照,蘆葦不收割)、W2〔枯萎前(10月)收割〕、W3〔枯萎中期(12月)收割〕、W4〔枯萎末期(翌年2月)收割〕,每個處理3次重復. 收割方式為去除濕地系統表面10 cm以上的植物部分,并定期對凋落物進行清理.
試驗在自然環境中進行,第1年試驗結束后,重新布置第2年試驗,2年的進水ρ(TN)不同,第1年較高〔相當于小麥季地表徑流中較高ρ(TN)〕,第2年較低〔相當于小麥季地表徑流中較低ρ(TN)〕[20],進水參數如表1所示. 以收割為節點將每年的試驗分為4個階段:階段1為試驗開始至W2收割前; 階段2為W2收割后至W3收割前; 階段3為W3收割后至W4收割前; 階段4為W4收割后至結束試驗.

表1 試驗期間進水水質情況
1.3 取樣與測定
水體取樣進出水同時進行,進水在布水區采集(3個重復),出水在每個濕地系統的出水口采集,每月一次(階段1),當收割開始后,每月2次(階段2、3、4),并現場測定系統中水體的pH、ORP、EC、ρ(DO)、溫度(T)等指標.ρ(TN)采用GB 11894—1989《水質 總氮的測定 堿性過硫酸鉀消解紫外分光光度法》測定,ρ(NH4+-N)、ρ(NO3--N)和ρ(NO2--N)使用流動分析儀(AA3,Seal,德國)測定.
根系取樣在每次收割時和試驗結束時進行,方法是小心將根袋從系統中拿出,為了彌補系統取樣后的植物損失,補充另外培養的對應處理蘆葦維持系統平衡. 根系活力采用氯化三苯基四氮唑(TTC)法測定. 己糖激酶(HK)、丙酮酸激酶(PK)、磷酸果糖激酶(PFK)、異檸檬酸脫氫酶(ICDH)、細胞色素氧化酶(COX)等根系呼吸代謝關鍵酶活性采用紫外可見分光光度計(DR 5000,Hach,美國)進行測定,且以μmol/min(以每mg蛋白質計)表示.
1.4 數據處理
利用IBM SPSS 22.0軟件對數據進行統計分析;采用單因素方差分析(one way ANOVA)檢驗不同收割處理對各參數的影響,顯著性檢驗采用Duncan在0.05顯著性水平下進行;用SigmaPlot 12.5軟件作圖,圖表中數據為平均值±標準差;利用Canoco 5.0軟件進行冗余分析.
2 結果與分析
2.1 枯萎期蘆葦收割對濕地脫氮效果的影響
從蘆葦濕地氮去除效果(見圖2)可以看出,2年間TN和NH4+-N去除率的變化趨勢相似,與第1年相比,第2年TN和NH4+-N去除率更高,其中第1年W1處理的TN和NH4+-N平均去除率分別為51.2%±24.5%和59.0%±21.9%,第2年分別為64.5%±25.8%和71.8%±30.5%.
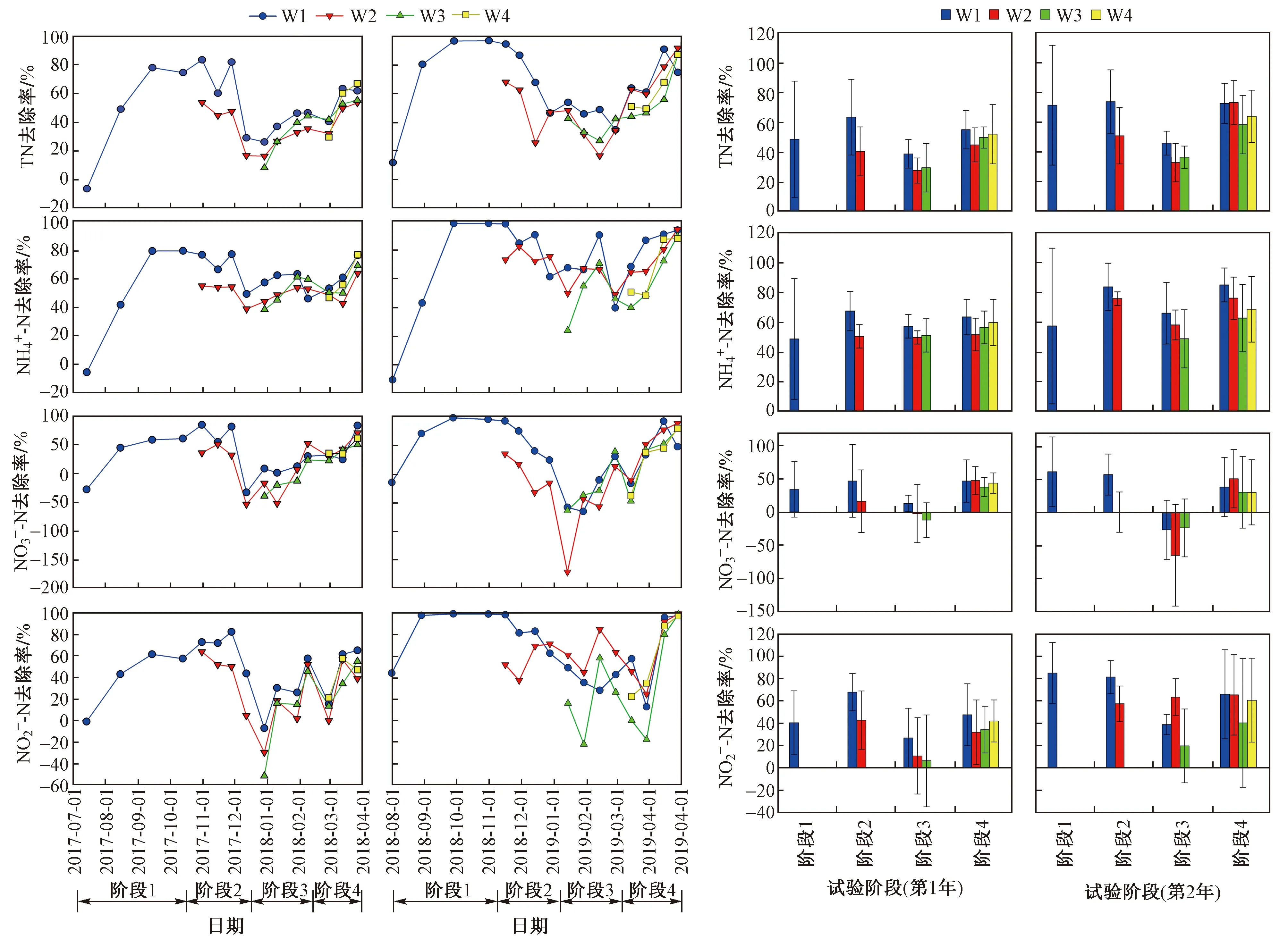
圖2 不同收割時間下蘆葦濕地TN、NH4+-N、NO3--N和NO2--N去除率的變化Fig.2 Removal of TN, NH4+-N, NO3--N and NO2--N from the reed wetlands under different harvesting time
從不同處理上看,與收割處理相比,不收割處理的TN和NH4+-N去除率較高. 每次收割后,TN和NH4+-N去除率均為下降趨勢,W2處理使階段2的TN和NH4+-N去除率在第1年分別下降了23.0%和17.0%,第2年分別下降了22.5%和7.5%,隨著時間的延長,去除率降低趨勢逐漸減小,但始終低于W1處理. W3處理TN和NH4+-N去除率的變化趨勢與W2處理相似,階段3的TN和NH4+-N去除率在第1年分別下降了24.0%和10.8%,第2年分別下降了18.2%和25.2%. 與W2和W3處理相比,W4處理TN和NH4+-N去除率的下降趨勢較小,階段4的TN和NH4+-N去除率在第1年分別下降了3.0%和3.7%,第2年分別下降了8.0%和16.0%.
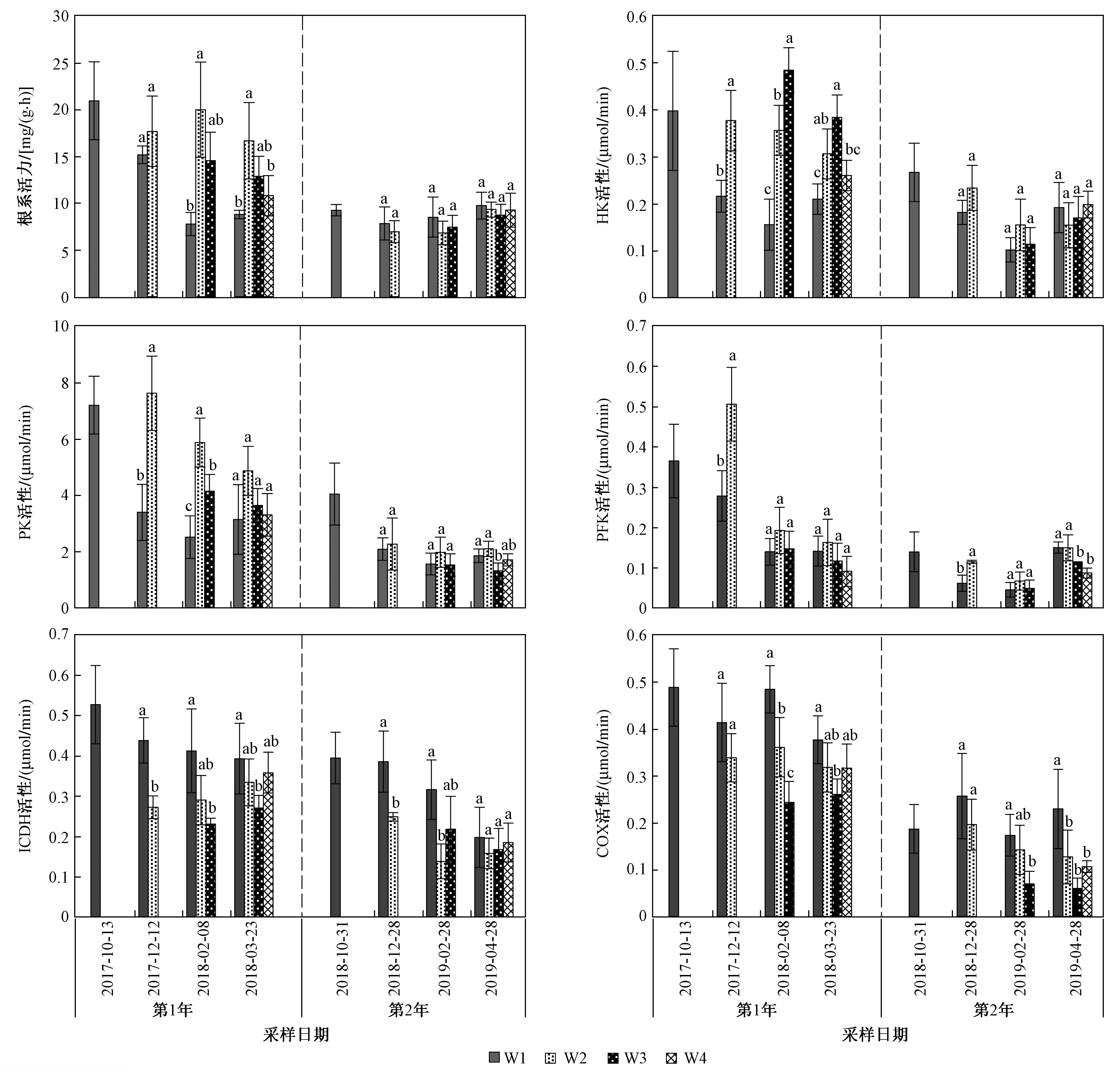
注: 不同小寫字母表示相同采樣日期不同處理下各參數差異顯著(P<0.05).圖3 不同采樣日期不同處理蘆葦根系活力和呼吸代謝關鍵酶活性Fig.3 Reed root activity and key enzyme activity of respiratory metabolism under different treatment on different sampling dates
2年試驗中階段3的氮(TN、NH4+-N、NO3--N、NO2--N)去除率均最低,且NO3--N和NO2--N去除率波動均較大,甚至有負值出現. 與TN和NH4+-N去除率趨勢一致,每次收割后,NO3--N和NO2--N去除率基本為下降趨勢,且W4處理的去除率下降趨勢最小.
2.2 枯萎期蘆葦收割對根系呼吸代謝的影響
年份和處理對根系活力和根系呼吸代謝關鍵酶HK、PK、PFK、ICDH、COX活性均有顯著影響(P<0.05);年份與處理的交互作用對根系活力以及HK、PK活性有顯著影響(P<0.05),但對PFK、ICDH、COX活性無顯著影響(P>0.05,見圖3).
試驗期間2年同期相比,第1年的蘆葦根系活力和呼吸代謝關鍵酶活性均明顯高于第2年(P<0.05). 第1年,與不收割相比,收割處理均增加了蘆葦根系活力以及HK、PK活性,且W2和W3處理增加的更為明顯(P<0.05);第2年,處理間根系活力以及HK、PK活性基本無顯著差異. 2年試驗中,W2處理均顯著增加了剛收割后的根系PFK活性(P<0.05),而隨著試驗時間的延長,差異逐漸不明顯,試驗第2年W4處理顯著降低了PFK活性(P<0.05). 與W1相比,收割處理均降低了根系ICDH和COX活性,且W3處理對第1年的ICDH活性和2年的根系COX活性降低的更為明顯(P<0.05);第2年,W2處理對根系ICDH活性降低的更顯著(P<0.05).
2.3 濕地脫氮效果與蘆葦根系呼吸代謝的關系
用RDA分析濕地脫氮效果與蘆葦根系呼吸代謝的相互關系,以根系活力和呼吸代謝關鍵酶活性為環境變量,解釋濕地氮去除率的變化. 結果(見表2)顯示,試驗第1年,軸1和軸2的特征值分別為0.847和0.069,去除率與呼吸代謝關系累計解釋量為96.8%;試驗第2年,軸1和軸2的特征值分別為0.696和0.061,去除率與呼吸代謝關系累計解釋量為93.4%,說明軸1和軸2能較好地反應二者之間的關系,且主要由軸1決定.
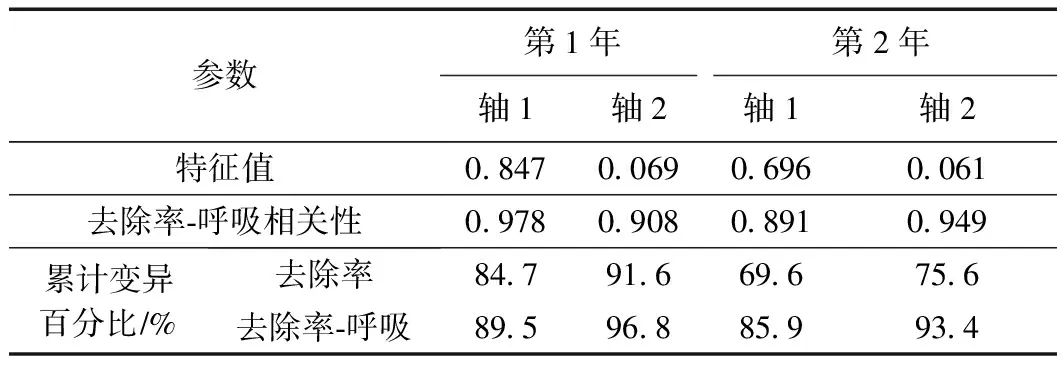
表2 濕地氮去除率與蘆葦根系呼吸代謝關系的RDA結果
由圖4可見,試驗第1年,濕地氮去除率的變化整體上由ICDH和HK活性主導,其與ICDH活性呈正相關,與HK活性呈負相關. 試驗第2年,濕地氮去除率的變化由根系呼吸代謝指標(HK、PK、PFK、ICDH、RA、COX活性)主導,均呈正相關,且HK活性對濕地氮去除率的影響最大.
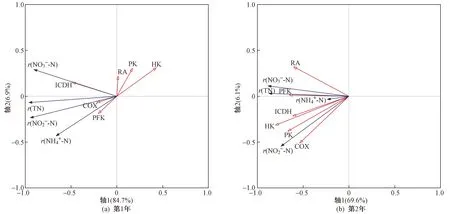
注: r(TN)表示TN去除率; r(NH4+-N)表示NH4+-N去除率; r(NO3--N)表示NO3--N去除率; r(NO2--N)表示NO2--N去除率; RA表示根系活力; HK表示己糖激酶活性; PK表示丙酮酸激酶活性; PFK表示磷酸果糖激酶活性; ICDH表示異檸檬酸脫氫酶活性; COX表示細胞色素氧化酶活性.圖4 濕地氮去除率與蘆葦根系呼吸代謝關系的RDA分析Fig.4 RDA analysis of wetland nitrogen removal and reed root respiratory metabolism
3 討論
3.1 不同收割時間對濕地脫氮效果的影響
溫度影響濕地的脫氮效率[21],該研究中,階段3的濕地脫氮效率最低,可能與低溫條件抑制生物過程有關[22],這與Hu等[23]的研究結果一致,其證明了冬季(12月—翌年2月)濕地出水ρ(NO3--N)增加,反硝化作用受到抑制. 該研究中,試驗第1年的進水ρ(TN)高于第2年,而第2年的TN和NH4+-N去除率高于第1年,這與Liu等[24]的研究結果不同,其表明進水氮濃度增加,TN和NH4+-N去除率顯著提高. 濕地的脫氮效率不是絕對的,受到各種因素的影響,如環境因子(pH、溫度、ORP等)、系統配置、水力狀況、進水模式、植物種類及植物收割方式等[25-27].
收割是濕地植物的主要管理方式[14],應優化收割時間,以提高濕地在冬季的脫氮效率. 該研究中,2個試驗年之間表現出相同的脫氮趨勢,均為不收割處理的TN和NH4+-N去除率最高,這與Shuai等[28]的研究結果不同,其表明與不收割處理相比,收割處理提高了濕地脫氮效率. 該試驗過程中定期對凋落物進行清理,避免蘆葦中的氮通過腐爛返回濕地中,更有利于解釋收割時間對濕地脫氮效果的影響. 研究[29]表明,植物地上部收割會影響反硝化微生物生長的根際微環境,從而影響濕地脫氮過程. 該研究結果表明,收割時間越晚,對濕地脫氮效果的影響越小,與不收割處理相比,枯萎末期收割的TN和NH4+-N去除率下降趨勢較小,可能是因為不收割蘆葦可以為濕地保溫,促進了冬季的反硝化作用,而枯萎末期收割后,氣候逐漸變暖,為植物的生長提供了有利條件[30],從而有利于濕地系統恢復脫氮效果. 另外,植物根系的泌氧作用會影響微生物的硝化-反硝化作用,從而間接影響濕地的脫氮效率[31]. 枯萎末期收割保持了蘆葦在冬季的完整性,這可能會保護植物根系,從而持續為根際環境供給氧氣[32],減少了對濕地脫氮效率的影響.
3.2 不同收割時間對根系呼吸代謝的影響
氮是植物生長發育必需營養元素,環境中的氮含量與植物的生理生化過程有密切關系,對維持植物呼吸的作用較為顯著[33]. 根系呼吸作用是植株地下部代謝的重要過程,它對養分的吸收、根系更新以及植株生長發育具有重要意義,極易受到外界條件變化的影響[34]. 收割作為濕地人工干預最常見的方式,可能會引起收割后植物根系呼吸代謝的變化,這也是植物適應干擾的方式. 根系活力的大小反映根系代謝能力強弱,研究[35]表明,根系對外界產生應激反應的表現是根系活力先快速提高,通過加強代謝來緩解干擾傷害,促進養分和水分的吸收,隨著脅迫時間延長,植物本身儲存物質消耗加劇,根系活力又降至對照水平. 該研究發現,試驗第1年,收割處理提高了蘆葦根系活力,且越早收割越能增強蘆葦冬季的根系活力,枯萎末期收割則對根系活力影響較小;這可能與根系的應激反應有關,早期的收割處理對蘆葦的干擾可能較大,植物為獲得足夠的氮而做出補償性反應[36],使得根系活力增加;晚期收割時,由于植物本身處于休眠期結束開始生長階段,收割處理對其干擾較小,根系活力與不收割相比差異不顯著. 試驗第2年,收割處理對根系活力無顯著影響,可能是在低進水ρ(TN)條件下,植物根系活力顯著低于高進水ρ(TN)條件(見圖3),收割干擾引起的應激反應與不收割相比差異不顯著,說明根系活力不僅受收割時間的影響,還與環境中氮濃度有關.
植物呼吸代謝中的糖酵解(EMP)途徑是有氧呼吸和無氧呼吸的共同途徑,HK、PK、PKF是調控EMP途徑的關鍵酶. 一般情況下,植物呼吸代謝以EMP途徑進入三羧酸(TCA)循環為主,TCA循環是植物維持生命活性所需的主要能量來源,ICDH是調控TCA循環的關鍵酶. 細胞色素(CP)途徑是末端氧化電子傳遞鏈的主要途徑,COX是鏈上的終端酶[34]. 該研究中,收割處理使根系呼吸代謝關鍵酶活性變化趨勢在2年試驗之間有顯著差異,說明根系呼吸代謝的變化與進水ρ(TN)高低有關,環境中的氮對根系呼吸的調控作用甚為重要,根系呼吸在進水ρ(TN)高的情況下較高,這與陳貴等[37]研究結果一致. 收割處理下HK、PK和PFK活性的變化規律基本一致,試驗第1年,枯萎前期收割均顯著增加了HK、PK和PFK活性,可能是收割時根系活力較強,干擾后植物為獲得足夠的氮而做出補償性反應[36],EMP途徑加快,而枯萎末期根系活力較弱,收割處理與不收割處理相比差異不顯著. 但是冬季植物呼吸代謝并非越高越好,有研究[38]表明,冬季植物加強維持呼吸,會導致植物碳儲量耗盡,減少翌年春季新根生長能力和植物的生產力,抵消春季溫暖給植物帶來的積極影響. 試驗第2年,枯萎中期和末期收割均會降低PK、PFK活性,EMP途徑也有一定程度的受阻. 該研究中,2年試驗的收割處理均降低了ICDH和COX活性,且與枯萎末期收割相比,早期收割降低的更為明顯,說明收割會影響根系呼吸中的TCA循環和CP途徑,且末期收割的影響程度相對較小.
3.3 濕地脫氮效果與根系呼吸代謝的關系
濕地污染物凈化效果與植物的種類及生理特性關系密切[39]. 研究表明,根系呼吸最小值出現在12月—翌年3月[40],而該研究結果顯示該時間段濕地脫氮效率同樣最低. 通過RDA分析表明,根系活力和呼吸代謝關鍵酶活性與濕地氮去除率之間密切相關(第1年解釋量為96.8%,第2年解釋量為93.4%),說明冬季蘆葦根系呼吸代謝機制是其適應收割干擾的重要策略,與濕地脫氮效果存在密切關系. 試驗第1年,濕地氮去除率的變化整體上由ICDH和HK主導,其與ICDH活性呈正相關,與HK活性呈負相關. ICDH是連接C-N代謝酶的關鍵,是TCA循環中最重要和最具有調控意義的限速酶[41]. ICDH活性增加會提高根系呼吸和根系活力,從而增加根系泌氧促進微生物的硝化-反硝化過程[31],提高濕地的脫氮效率. HK催化EMP途徑的第一步反應,將葡萄糖磷酸化,這有利于它進一步參與合成和分解代謝[34]. HK活性對EMP途徑的影響最大,HK活性降低會使EMP途徑貢獻率降低,而濕地氮去除率增加,這與第2年試驗結果(HK活性與濕地氮去除率呈正相關)正好相反,可能是根系呼吸代謝途徑對環境氮濃度高低的響應不同,從而影響濕地脫氮效果. 該研究中采取的試驗方式是將濕地中發生的生物和非生物反應整合起來評估濕地脫氮效果,以探索最佳收割時間,根系相關的或其誘導機制還需要更加深入的研究,以了解各反應機制在冬季濕地脫氮過程中的貢獻.
4 結論
a) 枯萎期不同時間收割影響蘆葦濕地的脫氮效果和根系的呼吸代謝特征. 與不收割處理相比,收割處理會降低濕地的脫氮效果,但枯萎末期收割對濕地脫氮效果的影響較小.
b) 收割時間對蘆葦根系活力和呼吸代謝關鍵酶的影響與進水ρ(TN)高低有關,在進水ρ(TN)較高的年份,枯萎前收割顯著增強了蘆葦的根系活力以及HK、PK、PFK活性;在進水ρ(TN)較低的年份,收割處理對根系活力以及HK、PK、PFK活性的影響不大,而收割處理均降低了2個年份的根系ICDH和COX活性,但枯萎末期收割對其影響程度相對較小.
c) 收割時間對冬季蘆葦濕地脫氮效果的影響與根系呼吸代謝特征存在一定的相關性,進水ρ(TN)較高的年份,濕地氮去除率的變化整體上由ICDH和HK活性主導;進水ρ(TN)較低的年份,濕地氮去除率的變化由根系呼吸代謝指標(HK、PK、PFK、ICDH、RA、COX活性)主導,均呈正相關,且HK活性對濕地氮去除率的影響最大.
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
中老年保健(2021年12期)2021-11-30 02:58:01
當代陜西(2021年2期)2021-03-29 07:41:24
攝影之友(影像視覺)(2019年2期)2019-03-05 08:27:14
中華詩詞(2018年11期)2018-03-26 06:41:34
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
Coco薇(2016年8期)2016-10-09 02:11:50
中國塑料(2016年3期)2016-06-15 20:30:00
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
